
古代象の生態 (中新世の古代ゾウ)
TOP 車山お知らせ 車山ブログ 歴史散歩 車山高原野鳥 たびネット信州
| 目次 |  |
| 1)現生のゾウの起源 大西洋中央海嶺とメタンハイドレート 円石藻 |
|
| 2)アジアゾウと2種のアフリカゾウの系統 | |
| 3)サバンナゾウとマルミミゾウの交雑 (ウガンダ共和国) / (アフリカゾウの交雑) |
|
現生のゾウの起源については、新生代古第三紀暁新世の約6,000万年前に遡る。中生代白亜紀末の大量絶滅は、約6,600万年前に恐竜を含む多くの生物が絶滅した。それから600万年も経たないうちに、地球上では新しい哺乳類の時代が始まっていた。その裏付けとされているのが、新たに発見された6,000万年前の生物の頭骨の破片で、確認されている限りではゾウの祖先と言われている。
フランスのパリの国立自然史博物館の古生物学者エマニュエル・ゲールブラン氏が、モロッコ最大の都市カサブランカの東100kほどの盆地でこの頭骨の破片を発見した。ウサギほどの大きさしかないが原始ゾウのものだった。さらに、化石には、頭蓋骨(前頭骨と鼻骨)、下顎の破片と歯、上顎と下顎を含む15の化石が含まれていた。肩の高さは約20cm、体重は約5-6kgでした。最も古く、最も小さく、最も原始的な、ゾウの親戚として考えられている。そのゾウの祖先とされる生物の学名は エリテリウム・アズーゾルムEritherium azzouzorumと呼ばれ、恐竜が絶滅した次代の暁新世に生息していた。
エリテリウム・アズーゾルムEritherium azzouzorumの名前は、ギリシャ語の「eri」(原始的、初期)と「therion」(獣)から来ており、azzouzorumは化石が発見されたモロッコのオウレド・アズーズ村にちなむ。 この生物はウサギほどの大きさで、現生のゾウのような長い鼻や大きな耳を持っていたかどうかは不明だが、牙の原型と考
 えられる歯を持っていたとされている。
えられる歯を持っていたとされている。約6,600万年前に起こった地球規模の大量絶滅事件で爬虫類の時代は終わりを迎えると、それから600万年も経たないうちに、新しい哺乳類の時代が始まっていたという。その裏付けとされているのが、新たに発見された6,000万年前の生物エリテリウム・アズーゾルムEritherium azzouzorumである。
azzouzorumは、化石が発見されたモロッコのシディ・チェンナン近くのオウレド・アズーズ村に捧げられている。
フランス国立自然史博物館の古生物学者エマニュエル・ゲールブラン氏が、モロッコ最大の都市カサブランカの東100kmほどの盆地でこの頭骨の破片を発見した。それがウサギほどの大きさしかない原始ゾウのものだった。
暁新世(約6,600万年前~5,600万年前)は、新生代の最初の世であり、この時代に、恐竜が絶滅した後に哺乳類が急速に進化し始めた時期として知られている。最初に現れた哺乳類としては、齧歯類(げっしるい)や初期の霊長類が確認されていたが、今回の発見でゾウの祖先も含まれた。
初期の霊長類は、暁新世初頭約6,590万年前に生息していた「プルガトリウス・マッキーベリPurgatorius mckeeveri」という小型の哺乳類が知られている。この種が生息していたのは、小惑星の衝突により、恐竜など地球上の生物の約4分の3が死滅した大量絶滅から、わずか10万年後のことだった。この時期の霊長類は、木の上で果物を食べる生活をしており、ゴリラやチンパンジー、最終的には人類へとつながる進化の系譜を形成する。
齧歯類は小型の哺乳類で、後に多くの種に分化し、哺乳類の中で最も種類が多い。リス・ネズミ・ヤマアラシの3亜目に大別される。その特徴は、上下の顎に一生伸び続ける一対の門歯(前歯)を持ち、これを使って木の皮や種子などをかじる。その鑿の形をした門歯は常に物をかじることで削れるが、一生伸び続けるため適切な長さを保つ。
霊長類は、現代のサルやヒトの祖先にあたるグループで、暁新世の終わり頃には、より進化した形態を持つようになった。
霊長類の進化は約6,590万年前、暁新世初期に始まったと見られている。最古の霊長類の化石、プルガトリウス・マッキーベリPurgatorius mckeeveriは、アメリカ合衆国のモンタナ州北東部ジョーダン近郊の「ヘルクリーク累層」で発見された。このヘルクリーク地域では、恐竜が絶滅した白亜紀末から暁新世初期にかけてのプレシアダピス目 Plesiadapiformesの化石が多く見つかっている。プレシアダピス類は、暁新世前期の約6500万年前から始新世前期の5500万年前にかけて、主にヨーロッパや北アメリカに生息していた絶滅したグループであるが、霊長類に非常に近い関係にあるとされている。化石は歯とアゴの一部、足首の関節しか見つかっておらず、サル目(霊長目)に分類しない研究者もいるが、見つかった足首の関節は可動域が広く、サル並みに木登りに長けていたようだ。プレシアダピス目が生きていた時代は、新生代古第三紀暁新世で、地球全体の気温が高く、世界中で熱帯雨林が広がっていた。プレシアダピス目は主にヨーロッパや北アメリカに生息しており、霊長類の祖先または近縁種とされている。その特徴としては、長い指や爪、そして木の上での生活に適応した体型が挙げられいる。この小型の哺乳類は、恐竜が絶滅した直後の時代に生きており、現在の霊長類の先駆者とされている。
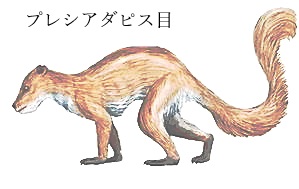 |
| プレシアダピス目は、約6,500万年前から5,500万年前の暁新世から始新世にかけて生息していた絶滅した哺乳類の目の一つである.。 霊長目の近縁またはその祖先と考えられている。霊長類は通常、握るのに適した手足と長い指、反対の親指と親指、ほとんどの指に爪があり、そして木の中を動き回るのに有益な骨格特性を持っている。 プレシアダピスは主にヨーロッパや北アメリカに生息しており、小型の哺乳類で、体長は10cmから1m程度、体重は最大で2〜3kgと推定されている。 食性は主に昆虫や小型の爬虫類、哺乳類などを捕食していたようだ。 霊長類の起源をめぐる議論の焦点となっている化石のグループは、プレシアダピフォームと呼ばれている。これは、11の異なるファミリーに配置された140以上の名前付き種を含む、極めて多様なグループである。 そのプレシアダピスの最初の記録は、非鳥類の恐竜が約6,500万年前、暁新世の初め近くに絶滅したちょうどそのときに現れた。 一部のプレシアダピス目は始新世晩期まで存続し、最後の種は始新世約3,700万年前頃.に絶滅した。 |
「この化石の下アゴから突き出ている2本の短い前歯から、この生物の素性を詳しく知ることができる。同時代の化石で、ほかにこのような歯を持つものは発見されていない。現生ゾウが有する牙の原型なのかもしれない」。
「頭骨の破片から判断して、体長は50㎝ほどしかなかったと推定される。体格的には、大型のウサギといったところだ。ただ、頭骨とアゴの破片しか発見されていないため、正確な姿形は不明だ。長い鼻や大きな耳といった現生のゾウに似た特徴を有していたかどうかは確認できない」。
この動物の化石には、2つの後小臼歯と3つの大臼歯を持つ顎、前頭骨と鼻骨、顎の破片、その他の孤立した歯が含まれていた。プレシアダピス目の化石は、パリにあるフランス国立自然史博物館に保存されている。その残骸だけでは動物の詳細な復元にまでいたらなかったが、エリテリウムEritheriumが、ゾウ目Proboscidea(長鼻目)の絶滅した属で、最古のゾウ目を代表するとされた。エリテリウムは約6,160万年前から5,600万年前の暁新世に北アフリカのモロッコで生息していた。この小型の哺乳類は、肩高約20cm、体重約5〜6kgの大きさであった。エリテリウムの化石は、モロッコのウルド・アブドゥン盆地で発見され、ゾウ目の進化を理解する上で重要な手がかりとなった。このエリテリウムの下顎骨の歯列は、下顎枝ごとに3つの切歯、1本の犬歯、4本の小臼歯、3本の大臼歯という2つの残された断片から再建された。歯列は閉じられており、犬歯と奥歯の間に隙間はなかった。この特徴は、他の既知のゾウ目には見られない。ゾウ目の現生種は3種で、全てゾウ科に含まれるアフリカゾウとアジアゾウである。ゾウはその巨大な体と長い鼻(象鼻)が特徴的で、知能が高く、社会的な動物である。
前述されているように、エリテリウムEritheriumは、ゾウ目(長鼻目)の最古の祖先とされている。エリテリウムは約6,000万年前の暁新世に生息していた小型の哺乳類で、モロッコで化石が発見された。この生物は、ゾウ目の進化を理解する上で重要な手がかりとなっている。 エリテリウムは、肩高約20cm、体重約5〜6kgと推定されており、現代のゾウとは大きく異なる小型の動物であった。その名前は、ギリシャ語の「eri」(早い)と「therium」(獣)に由来しているように、ゾウ目の進化の過程で、エリテリウムのような初期の祖先から、現在のアフリカゾウやアジアゾウのような大型のゾウへと進化していったと考えられている。
6,000万年前、中生代白亜紀が終わり新生代古第三紀暁新世の始まるあたりに恐竜が絶滅し、哺乳類が急速に進化し始めた時期である。暁新世(約6,600万年前から約5,600万年前)は、地球全体の気温が高く、湿度も高い時代であった。北極や南極も温暖で、氷河の形成もない、この時代の当初は、白亜紀末の気候変動の影響を受けており、気候はやや不安定であったが、全体的には温暖な環境が続いていた。
さらに、暁新世の終わり頃には、急激な全地球的温暖化現象「暁新世-始新世温暖化極大Palaeocene–Eocene thermal maximum(PETM)」が発生し、地球の平均気温が5〜9℃上昇した。アフリカ大陸は既に北方のユーラシア大陸から分離しており、植物が豊富に生い茂っていた。フランス国立自然史博物館の古生物学者エマニュエル・ゲールブラン氏の言葉を借りると、当時のアフリカの温暖化は「進化のゆりかご」だった。
今回の発見によって、恐竜が絶滅した直後の地球上にゾウのような哺乳類が出現していたことが判明した。
大西洋中央海嶺
暁新世が終わる約5,580万年前に起きた急激な温暖化は「暁新世-始新世温暖化極大」と呼ばれている。それは数千年あるいはそれ以下の短期間で進行したと推定されている。この時期、地球全体の平均気温が5〜9℃上昇し、海洋の酸性化を進行させた。その原因として、北大西洋で海底火山活動が活発化し、海底堆積物中のメタンハイドレートが融解したという説が有力視されている。
大西洋中央海嶺は、中生代三畳紀Triassic period(約2億5217万年前~約2億130万年前)に形成が始まり、この時期に超大陸パンゲアが分裂し、アフリカ大陸・南米大陸・北米大陸・ユーラシア大陸が分離していき、その結果、大西洋が拡大し、その地殻の割目から大西洋中央海嶺が形成された。この海嶺は、地球のプレートが東西に広がる境界に位置しているため、マントルから噴出したマグマが新しい海底地殻を生成する場所となる。そのため、現在でも大西洋は毎年約4cmずつ広がっている。この三畳紀中期には初期の恐竜や翼竜、ワニなどが出現し、陸上生態系において支配的な地位を占めるようになる。
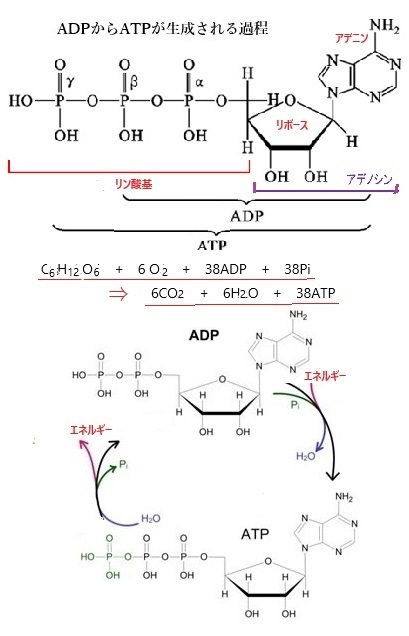 |
| ADPからATPへの合成反応は、細胞内でエネルギーを蓄える重要なプロセスとなる。この反応は主に次のように行われる。 ミトコンドリア内で行われるこのプロセスでは、電子伝達系を通じてプロトンH+の濃度勾配が形成され、そのエネルギーを利用してADPにリン酸基が付加され、ATPが生成される。 植物の葉緑体で行われる光合成の一部では、光エネルギーを利用してADPにリン酸基が付加され、ATPが生成される。 ATP合成酵素(ATPシンターゼ)は、これらのプロセスで重要な役割を果たし、ADPと無機リン酸PiからATPを合成する。ATPは、アデノシンという物質に3つのリン酸基Pが結合している。ATP分解酵素の働きによってATPが加水分解すると、ひとつのリン酸基(P)がはずれてADPになり、その際にエネルギーを放出する。 このエネルギーの供給は、 ATPは「エネルギーの通貨」とも呼ばれ、細胞内にエネルギーを供給する役割を担う。ATPが分解されてADPとリン酸に変わる際にエネルギーが放出され、そのエネルギーが生物の生命活動や化学反応の推進などに利用される。 |
| ADPと水H2OからATPを生成するプロセスは、細胞のエネルギー代謝において非常に重要で、この反応により生成されたエネルギーが、それを必要とする多くの生物的合成に関わるプロセスに参加する。
具体的には、ADPと無機リン酸Piが結合してATPを形成する。この反応にはエネルギーを吸収するためのエネルギー供給源が必要で、通常、このエネルギーは細胞呼吸や光合成などの過程で供給される。 ATPは、細胞内でエネルギーを運搬する「エネルギー通貨」として機能し、必要なときにエネルギーを放出してADPとPiに戻る。このサイクルが繰り返されることで、細胞は効率なエネルギー循環を達成する。 プロトンH+勾配によりATP合成酵素を通過する際に、ADPと無機リン酸Piが結合してATPを生成する。 この反応には、エネルギーを効率的に蓄えるために不可欠で、ATPは細胞内のエネルギー通貨として機能し、様々な生物生命活動に必要なエネルギーを供給するためである。 ATPの加水分解は、エネルギーを放出する重要な反応で、この反応により、ATPが水H2Oと反応してADPと無機リン酸Piに分解される。この過程でエネルギーが放出され、生物のさまざまな活動に利用される。 ATP + H₂O → ADP + Pi + エネルギー |
(好気的呼吸における、ブドウ糖glucoseの酸化の具体例は
ブドウ糖C6H12O6の1分子が酸素6分子と反応して、二酸化炭素CO2の6分子と水H2Oの6分子を生成する。この過程で、アデノシン二リン酸ADPからアデノシン三リン酸ATPが生成され、エネルギーが供給される。
この反応は
C6H12O6 + 6 O2 + 38ADP + 38Pi → 6CO2 + 6H2O + 38ATP
このプロセスは、解糖系・クエン酸回路、および酸化的リン酸化の3つの段階で進行する。)
海中の微生物は、海水中の酸素O2を利用して有機物を分解し、エネルギーを得ている。この海水中の有機物が分解される過程で、二酸化炭素CO2が生成される。これが海洋の炭素循環の重要なプロセスとなる。
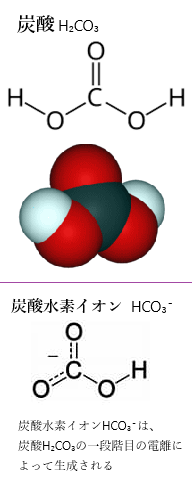
有機物が海水中で分解されると、酸素O2が消費され、二酸化炭素CO2が生成される。このCO2は水H2Oと反応して炭酸H2CO3を形成する。これにより、海水中の炭酸の全濃度が増加する。
このプロセスで生成される二酸化炭素CO₂は海水に溶け込み、炭酸H₂CO₃として存在するようになる。この結果、海水中の炭酸の全濃度が増加する。この現象は、水素イオンH+濃度を高め海洋酸性化の一因となり、海洋生態系に影響を与える。特に、酸素が少なくなると、海洋生物にとって生存が難しくなる。
大気中に排出される二酸化炭素は、地球温暖化を引き起こす主要な温室効果ガスとなる。地球温暖化は、海水温の上昇や海面水位の上昇を引き起こし、海洋環境や気候変動に大きな影響を及ぼす。さらに近年、大気中に放出された二酸化炭素を海洋が吸収していることによる「海洋酸性化」が指摘されている。
大気から海洋へのCO₂の移動量と、海洋から大気へのCO₂の移動量が等しくなり平衡状態になる。この状態が維持されることで、見かけ上はCO₂の濃度に変化がないように見えるが、実際には両者の間で活発な交換が行われている。
大気中の二酸化炭素濃度が年々増加すれば、大気と海洋が接している海面では、大気と海洋の間で二酸化炭素の交換が行われている。つまり大気中の二酸化炭素の増加に伴って、海水に溶け込む量が増えるため海洋表面の二酸化炭素も年々増加する。この交換が均衡状態にあるとき、大気から海洋へのCO₂の移動量と、海洋から大気への移動量が等しくなり、この状態を「平衡状態」と呼ぶ。しかし、現在のように人間活動によって大量のCO₂が大気中に放出されると、この均衡が崩れ、大気中のCO₂濃度が増加する。海洋に吸収された二酸化炭素は、海洋の循環や生物活動により深層に運ばれ蓄積されていく。
海水中のpHは一般的に弱アルカリ性を示し、表面海水中のpHは約8.1、そこから深くなるにつれてpHは下がり、北西太平洋亜熱帯域では水深1,000m付近で約7.4と最も低くなる。これは、深くなるにつれて有機物の分解により海水中の酸素O2が消費され、炭酸H₂CO₃の全濃度が増加することによる。この過程で二酸化炭素CO₂は海水に溶け込み、炭酸H₂CO₃を生成することになる。この結果、海水中の炭酸の全濃度が増加する。この現象は、海洋酸性化の一因ともなり、海洋生態系に影響を与える。特に、酸素が少なくなると、海洋生物にとって生存が難しくなる。
二酸化炭素が多く溶け込むとpHが下がり、海水のアルカリ性が弱まる。海洋酸性化の指標として用いられるpHは、水素イオンH+濃度の逆数の対数で定義される値であり、水素イオン濃度が増えるとpHは下がりる。このように海洋のpHが長期にわたって低下する現象を「海洋酸性化」と呼んでいる。海で進化してきた多くの水生生物はpH 8付近の環境に適応した生理をもち、極端なpH環境では限られた生物しか生きられない。大気濃度を安定化できても、海洋が「吸収してしまった」CO2を処理して海洋のCO2濃度を抑制することはほぼ不可能と言われている。海洋からCO2を取り除くことが困難となれば、一旦海洋生物と生態系へ影響が起こってししまえば回復方法がないことを意味する。
地球温暖化の進行により、海洋の二酸化炭素吸収能力や海洋の循環が変化することが想定される。
メタンハイドレートは、低温・高圧の環境下でメタン分子CH4が水分子H2Oに囲まれて形成される氷状の物質である。メタンハイドレートは「燃える氷」とも呼ばれ、1㎥のメタンハイドレートからは、常温常圧となれば約160㎥のメタンガスが取り出せる。つまり融解すると、メタンガスが大放出される。メタンハイドレートは低温高圧で安定しているが、温度が上昇するだけで融解しやすくなる。地表付近の大気の主な成分は、窒素N2が78.08%、酸素O2が20.95%、アルゴンAr(単原子分子として存在する)が0.93%、二酸化炭素CO₂が0.03%であれば、この融解過程で放出されるメタンガスCH4は、大気より軽い。それでも、二酸化炭素と比較しても、メタンガスの温室効果は約25~28倍も強力です温室効果ガスとしての影響が大きく、地球温暖化を促進するのは、メタンは赤外線を非常に効率的に吸収し、地球の表面から放出される熱を捕捉する。このため、少量でも強力な温室効果を発揮する。
(メタンは温室効果ガスの一つであり、赤外線を非常に効率的に吸収する。これにより、メタン分子はエネルギーを蓄え、そのエネルギーを熱として放出するため、周囲の温度が上昇する。 メタンの温室効果は二酸化炭素CO2に比べて約25倍も強力であるため、少量のメタンでも地球温暖化に大きな影響を与えることが知られている。)
北大西洋における火山活動の活発化は、北大西洋における海洋底拡大によるものであった。火山活動にともなって放出された二酸化炭素が原因となり、海水が酸性化する。海の表面に近い海水中の二酸化炭素濃度は、大気中の二酸化炭素を吸収するため、大気濃度を追いかけるようにして増加していることが世界中の海で観測されている。海洋は二酸化炭素を吸収することで、大気中の二酸化炭素濃度増加を抑え、その分だけ地球温暖化を減速させるが、海水中の二酸化炭素が増えると、炭酸が生成され、その結果として水素イオンH+が増加し、海水のpH値が低下し海洋の酸性化をもたらす。
 |
 |
CO2 + H2O ⇔ H2CO3
H2CO3 ⇔ H+ + HCO3- ⇔ 2H+ + CO32-
海水中に溶け込んだ二酸化炭素CO2は、炭酸水素イオンHCO3-や炭酸イオンCO32-と化学平衡の状態となり、その可逆反応において、順方向の反応と逆方向との反応速度が釣り合って反応物と生成物の組成比が巨視的に一定になる。つまり大気中の二酸化炭素CO2が増えると、これらの反応に伴って水素イオH+が解離し、海洋を酸性化させる。
炭酸H2CO3は、海洋中では水素イオンH+が解離した炭酸水素イオンHCO3-や炭酸イオンCO32-との間の反応により化学平衡の状態を保っている。そのため大気中の二酸化炭素が増えると、海水に溶け込む二酸化炭素も増え、水素イオンH+が発生する。これが海洋酸性化で、その海の生物に及ぼす最も重大な現象が、炭酸カルシウムCaCO3の形成を阻害することにある。
植物プランクトンの円石藻(えんせきそう)、動物プランクトンの有孔虫、サンゴ、ウニ、貝類などは、海中のカルシウムイオンCa²⁺と炭酸イオンCO32-を結合させて、水に溶けない炭酸カルシウムの殻や骨格を形成している。
Ca2+ + CO32- -> CaCO3
ところが、CO₂が海に溶けて、水素イオンH⁺濃度が上がると、増えた水素イオンH⁺のほとんどは海水中の炭酸イオンCO₃²⁻と反応して炭酸水素イオンHCO₃⁻を増やす。そのため海水中の炭酸イオンCO₃²⁻が減る。
H+ + CO₃²⁻ -> HCO3-
すると、海中のカルシウムイオンCa²⁺と炭酸イオンCO₃²とを結合させて炭酸カルシウムの殻や骨格を作っている生物は、殻や骨格を作りにくくなる。動植物プランクトンやサンゴ、ウニなどが減るため、魚の餌や生息域が減るので、魚にもその影響が長期化する。
(円石藻の細胞表面には炭酸カルシウムの鱗片があり、これが円石と呼ばれる。その円盤型の構造を持つ植物プランクトンで、単細胞の真核藻類として、海洋に広く分布する光合成独立栄養生物である。海洋の水深200m以上の太陽光が届く範囲の有光層で主に浮遊している。
葉緑体【小胞状の細胞小器官】にはクロロフィル【葉緑素】があり、これが光エネルギーを吸収して化学エネルギーに変換する。円石藻の葉緑体には核はないので、細胞内で自律的に分裂して増殖することはない。クロロフィルは特に青と赤の光を吸収し、緑色の光を反射するため、円石藻は緑色に見える。
海水1ℓあたりの生息密度は、熱帯~温帯の暖かい海では数千~数万個体、富栄養な温帯~寒帯の海では数万~数十万個体に達する。ただ沿岸にのみに分布している沿岸種も約40種あり、外洋ではほとんど見つからない。沿岸種の中には、海底や大型藻などに付着して生活しているものもある。その細胞の直径は2-50㎛と極めて微小で、肉眼では観察できない。
その微小な円石藻こそが、海洋の主要な一次生産者となり、他の生物にとってのエネルギーや栄養素の食物連鎖の重要な基盤となって生息している。また、円石藻の炭酸カルシウムCaCO3の鱗片は、海洋の炭素循環の要となり、地球の気候調節にも貢献している。円石藻の細胞表面を覆う円石により他のハプト藻と区別され、これを基に種の同定が行われている。)
海洋植物プランクトンの一種、ハプト藻類円石藻は細胞殻として方解石型炭酸カルシウム結晶か ら成る構造体を有する。円石藻は中生代三畳紀約 2 億年前に出現し、中生代白亜紀に大量に繁茂したこと で知られる。その大増殖の結果、円石藻が大量に海底 へと沈降し、そして蓄積した。そこで細胞殻は、主に炭酸カルシウムCaCO₃を50%以上含む堆積岩・石灰岩limestoneを形成し、また細胞の有機質部分は地球化学的作用により 原油・天然ガスの基となる有機物を大量に生産し、大気中のCO2を減少させた。
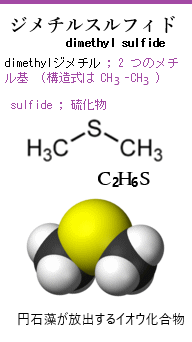
円石藻が放出するイオウ化合物の一つにジメチルスルフィド(DMS;化学式はC2H6S)が含まれる。 このDMSは、大気中に放出されると、酸化されて硫酸イオンや二酸化硫黄となり雲の凝結核(CCN)を形成する。これが雲核となり、雲の形成を促進する。これにより、太陽光の反射率が上昇し、気候に直接的な影響を与えることが知られている。
(「磯の香り」の本体である硫化ジメチルdimethyl sulfide【DMS】は、海洋から大気に放出され、大気中で硫酸エアロゾルとなり雲核として作用する。その主な発生源は、海藻や植物プランクトンが、硫黄を含む有機化合物を代謝する際に、ジメチルスルフィド (DMS) を生成する。 海中には多くの硫黄を含む有機化合物が存在する。これらは主に海洋生物の代謝産物や、海底の堆積物から発生する。
例えば、硫黄を含む有機化合物としては、硫化水素・硫酸塩・硫黄酸化物などがあり、これらの化合物は、海洋環境における化学反応や生態系の重要な要素となっている。
特に、ジメチルスルホニオプロピオネート(DMSP)は、海洋植物プランクトンや大型藻類の浸透圧調節物質として生成され、海水中の微生物の硫黄源として利用される。海水中に放出されたDMSPは、主にDMS代謝細菌によって3つの異なる代謝経路(酸化還元反応を通じて代謝産物の一つとして生成)で分解を受ける。
それぞれの代謝経路に関与する主なDMS代謝細菌は、それぞれ「DMSP リアーゼを持つ細菌」はDMSPを直接分解し、DMSを生成する。例えば、一部の細菌はDMSPをリアーゼという酵素を使って分解する。
「DMSP メチルトランスフェラーゼを持つ細菌」によるDMSPのメチル化とは、DMSPがメチル基(-CH3)を付加される化学反応のこと言う。この反応により、DMSが生成される。具体的には、DMSP メチルトランスフェラーゼの作用によりメチル基が転移され、DMSが生成されるというプロセスである。
「DMSP オキシダーゼを持つ細菌」は、DMSPを酸化し、DMSを生成する。例えば、一部の細菌はDMSPをオキシダーゼという酵素を使って酸化する。 これらの代謝経路を通じて、DMSは海洋中に放出され、大気中に拡散することで、地球の大気に影響を与えることがある。
大気中で酸化されたDMSは硫酸エアロゾルとなり、雲核として雲の形成が促進され、地球の気候環境に影響を与える可能性がある。 )
メタンガスは、海底のメタンハイドレートという氷状の物質から放出されることもある。メタンハイドレートが分解されると、メタンガスが海水中に放出され、これが酸性化を促進する。メタンガスは温室効果ガスとしても強力で、気候変動に大きな影響を与える。
さらに、メタンガスが海水に溶けると、炭酸ガスと同様に酸性化を引き起こす化学反応を進行させる。これにより、海洋生態系に悪影響を及ぼし、サンゴ礁や貝類などの石灰化生物の成長を妨げる。海洋酸性化とメタンガスの関係は複雑であるが、どちらも地球環境に重大な影響を及ぼす。
メタンガスCH₄が海水に溶けると、酸性化を引き起こす主な反応は
メタンガスが酸化されて二酸化炭素CO₂と水H₂Oを生成する。
CH4 + 2O2 → CO2 + 2H2O
生成された二酸化炭素が海水に溶けて炭酸水素H₂CO₃を形成する。
CO2 + H2O → H2CO3
この炭酸水素が水素イオンH⁺と炭酸水素イオンH2CO3に解離する。
H2CO3 → H+ + HCO3−
この一連の反応により、海水中の水素イオン濃度が増加し、pHが低下して酸性化が進行する。
目次へ
長鼻類(ちょうびもく)で最も進化したグループであるゾウは、新生代第四紀にはオーストラリアと南極大陸以外の全ての大陸に分布していたが、更新世の初頭ジェラシアン期の約258万年前から約1万年前までの新生代更新世であれば、主に氷河時代として知られ、地球全体で寒冷な氷期と比較的温暖な間氷期が繰り返された。この時代には、マンモスやネアンデルタール人などが生息していた。また、現生人類(ホモ・サピエンス)もこの時期に出現していた。その厳しい自然環境の変化や人類の狩猟などによりやがて衰退し、現在はサハラ砂漠以南のアフリカに生息するサバンナゾウLoxodonta africanaとマルミミゾウLoxodonta cyclotis(Loxodonta;アフリカゾウ属)、そしてインドおよび東南アジアに生息するアジアゾウElephas maximus(英語名:Indian elephant)のわずかに3種類が残るのみであり、滅亡へ向かいつつある哺乳類と言える状況にある。
 |
 |
 |
 |
 |
これらのゾウたちは、それぞれ異なる地域や環境で進化し、多様な形態や特徴を持っていた。
そのうち、現存しているのは、アジアゾウ・サバンナゾウ・マルミミゾウの 3 種となる。アジアゾウとアフリカに生息する 2 種サバンナゾウとマルミミゾウの共通祖先は、新生代新第三紀中新世の約760万年前に分岐し、サバ ンナゾウとマルミミゾウの共通祖先は、その約 100 万年後の同じ中新世分岐し始めた。
サバンナゾウとマルミミゾウ(森林ゾウ)の2種は、DNA解析により、両者の間には約58%の遺伝的違いがあることが分かっている。
染色体数は、サバンナゾウとマルミミゾウは、56本の染色体を持ち、対してアジアゾウは58本の染色体を持つ。
2 種間の交雑は限定的なものであると思われ、最近、調査が行われた広大な森林地帯からサバンナ移行 帯にまたがる 100 を超える地点のうち、交雑が明確に認められたのは 14 地点のみであった。こ れら 14 地点のうち 9 地点では、交雑個体はどちらか一方の種の非交雑個体とともに生息してい た。具体的に、3 つの地点では交雑個体とマルミミゾウのみが確認され、それらはマルミミゾ ウに分類され、6 つの地点では交雑個体とサバンナゾウのみが確認され、それらはサバンナゾ ウに分類された。国際自然保護連合(IUCN)のレッドリストの評価では、2015年の分布図と 2019年による最近のデータに基づき、これらの地点をサバンナゾウまたはマルミミゾウのいずれかの分布域に指定している。
アフリカゾウは世界最大の陸生哺乳類で、体長は最大6.5m、体重は10tにもなり体も大きい。アフリカゾウは、大きな耳と長い牙、そして長い鼻が特徴で、大きな耳は、表面積が広く、血管がたくさん通っているため、体温調節に役立つ。
アフリカゾウのオスとメスの牙には、機能的にも著しい違いがある。オスの長い牙は、土を掘ったり、木を倒したり、敵と戦ったりする時に使う。一般的に、オスのゾウの牙は長くて太く、より湾曲していることが多い。この長く、湾曲した牙を持つ個体が遺伝されるのは、草や木を引き抜いたり、他のオスと戦うことで、自然と湾曲するように進化し、またその長くて強い牙ほど、他のオスに対する威嚇力が向上し、それがメスをより引きつける魅了となったため「自然選択」され、その特性が次世代に引き継がれ、その特定の遺伝子が定着し、しかも発現しやすくなったためである。
つまり、アフリカゾウのオスの牙の形が、長年の多くの試練を克服し適応するため形成を積み重ね統合されたのは、「遺伝子の応え」による。一方、メスの牙は短くて細め、どちらも木の皮を剥ぐためや、食物を掘り出すために使うには都合がよい。また、牙を使って、地面を掘って根っこを食べたりすることもある。それは集団の中の子を中心にした「種の保存」を最優先にするメスゾウの進化の過程における光景となった。
長い鼻は、呼吸しながら匂いをかぐ、水を飲む、物をつかむなど、様々な用途に使われ、また、コミュニケーションにも有効で、挨拶をしたり、感情を表現したりする時に、鼻を絡ませたり、高く上げたりする。さらに様々な方法でコミュニケーションをとるが、諸々な音を出し、また、体を使って、挨拶をしたり、感情を表現したりする。時には、足の裏を使って、地面の振動を感知し、遠く離れた場所にいる仲間とコミュニケーションをとることもできる。
サバンナゾウの前歯が発達してできた牙は、マルミミゾウに比べて湾曲しており、一生伸び続けまる。 牙はオスのほうが大きく、60歳の個体ではオスが約110㎏、メスが約40㎏ほど、長いものは3.5mにもなります。サバンナゾウの耳は大きく、縦2m、横1.5mのもなる。
サバンナゾウは、アフリカの森林地帯にもいるが、主に標高2,500mまでのサハラ以南のサバンナ・山地林・低木林・草原・半砂漠などに定住している。サバンナとは、熱帯や亜熱帯域で、1年の中で雨季と乾季が存在し、年間降水量のあまり多くない。その「サバンナ気候」と呼ばれる土地に形成される草原地帯の年間降水量があまり多くない。大きな樹木ほど豊富な水を必要とするため、サバンナ気候のような降水量の少ない土地では育ちにくい。しかしながら、雨季には雨が降るため、まったく植物が生えない、という土地でもない。乾燥に比較的強いサバンナを広く覆うイネ科の植物が主体となり、サバンナという草原が成立する。サバンナゾウが生息するアフリカ大陸の広大な草原地帯サバンナは、主にアフリカの東部から南部にかけて広がっている。
サバンナゾウはかつてアフリカ全土に生息し、現在は 23 カ国に生息する。サバンナゾウの分布に関しては場所や時期により違いはあるが、大陸全体を通して 見ると、本種の分布域が縮小し、分断が進んでいることは明らかである。現在のサバンナゾウ の分布域は、農耕が始まる以前の分布域の 15%と推定される。ブルンジとモ ーリタニアでは国内のサバンナゾウは絶滅したと考えられている。
現生の在住は、アンゴラ・ボツワナ・カメルーン・中央アフリカ共和国・チャド・コン ゴ共和国・コン ゴ民主共和国(ザイール)・エリトリア・エチオピア・ケニア・マラウイ・マリ・モザンビーク・ナミビア・ ナイジェリア・ルワンダ・ソマリア・南アフリカ・南スーダン・タンザニア連合共和国・ウガ ンダ・ザンビア・ジンバブエとなっている。
アフリカのサバンナ地域の年間降水量は、地域によって異なるが、一般的には500mmから1,500mm程度である。雨季も地域によって異なりるが、多くのサバンナ地域では3月~5月と9月~11月が雨季となる。東京都の年間降水量は約1,482mmで、東京都は四季がはっきりしており、特に梅雨の時期や台風のシーズンに多くの降水が見られる。
サバンナゾウLoxodonta africanaが最も多く生息している国は、ボツワナで、約13万頭のサバンナゾウを抱えており、これは世界で最も多い。その理由の一つは、ボツワナが広大なサバンナに保護区を設けたことにより、サバンナゾウたちは、安全な生息地と十分な食料を確保できる恵まれた環境が保障された。
ところが「ボツワナのオカバンゴ・デルタには1万8000頭のサバンナゾウが定住していて、このゾウの主要な定住地の1つと考えられている。2020年から2021年にかけて、この地域のゾウの少なくとも450頭が謎の死を遂げた。」
「ゾウが目まいを起こしているようにクルクルと回り、突然、倒れて死ぬ。顔から倒れることもある。ボツワナではこの数カ月、数百頭のゾウが命を落としており、一部のゾウは前述のような症状を示していた。なぜかは誰もわからない。」
「(CNN) アフリカ南部のボツワナで野生のゾウ 300頭以上が死んでいるのが見つかった問題で、政府の野生生物保護当局は2020年9月21日に、毒素を生成するシアノバクテリア(藍色細菌)が水飲み場で繁殖していたことが原因だったと発表した。」
それに納得しない専門家は、政府の調査結果の全面的な開示を求めている。ゾウを専門とする保全生物学者のキース・リンゼイ氏は、「水飲み場が原因だったとすれば、なぜゾウだけが影響を受けたのか」と疑問を投げかけている。
イギリス連邦加盟国であるボツワナの政情は、1966年の独立以来、複数政党制の下、ボツワナ民主党(BDP)が政権を維持してきている。南部アフリカ諸国の経済的統合を目的とする南部アフリカ開発共同体(SADC)事務局の所在地にもなっている。非常に安定し、民主的な制度が整い、治安も良好、観光客にとって安心して訪れることができる国と評価されている。
ボツワナは、ダイヤモンドでは、世界有数な生産国であり、その採掘が主要な産業となっている。鉱物資源も豊富で、特に金やニッケルなどが採掘されている。民主的な制度が整っており、治安も良好で、観光客にとって安心して訪れることができるようだ。
サバンナの特徴としては、広大な草原に原生するサバンナ・アカシアやバオバブ(生息地はアフリカやインド、オーストラリアやマダガスカル)の高木と、アカシア属(アカシア・セネガルやアカシア・ターティリス、そしてアカシア・カラツラなど)の灌木が叢生する典型的なブッシュ風景で、これらの植物は、厳しい環境条件に適応しながら、サバンナの野生動物たちに重要な食料源や隠れ家を提供する。アフリカゾウは、広い行動範囲を持ち、食料や水を求めて長距離を移動し、草・木の葉・果実・樹皮・根など、1日に約200~300kgの植物を食べ、水も100ℓ以上飲む。
中国科学院武漢植物園や英ロンドン大クイーンメアリー校などの国際研究チームが、アオイ科バオバブ属の8種の全遺伝情報(ゲノム)を解析した成果を、2024年5月15日付の英科学誌ネイチャー電子版に発表した
「現存する8種のうち1種はアフリカ大陸の広域、1種は豪州北西部に分布し、残り6種はマダガスカル島西部に広がる。解析の結果、8種の共通祖先は新生代古第三紀始新世の約4,100万年前に出現し、新第三紀中新世の約2,100万年前にアフリカ大陸の種と豪州の種が分岐。残り6種が中新世の約1,200万年前までに分かれたと推定された。」
ゾウやキリンなどの大型の動物は、バオバブの巨大な幹や枝を食べることで水分を補給する。バオバブには他の樹木のような年輪がなく、その木の中はスポンジのような構造で、その中に水を大量に貯めこむ。バオバブは非常に乾燥に強く、驚くことに 2年間雨が降らなくても枯れない。 その秘訣は、幹の中にたっぷりと水分を貯蔵できることと、もともと葉の数が少なく、乾季には自分で葉を落とし蒸散を防いでいる。
バオバブは乾季には貴重な水源となる。アフリカでは乾季になるとゾウが樹皮を剥がして、水分をたっぷり含んだ中身を食べている姿も見られる。
バオバブは、今も昔もとても貴重な食料として利用されている。若葉や新芽なども生、もしくは調理して様々な方法で食べられている。果実にはカルシウムやビタミン、鉄分がたっぷり含まれ、その栄養の高さから、近年はスーパーフルーツとして世界中から注目を浴びている。 バオバブの花はコウモリや蜂によって受粉される。果実は猿や鳥などの食料にもなる。バオバブの木の洞は、小動物や鳥たちの安全な隠れ家としても利用される。住民は、果肉や油を取った後の果実の外側は、食器や器としても利用し、樹皮は編めば丈夫なロープとなり、解熱剤や下痢止めの薬にもなる。
乾燥したサバンナ地帯に広く分布するバオバブは、平均すると直径が10mほどあり、高さも約30mにまで成長し、幹の中に大量の水分を蓄える。その大きな個体に、約10tもの水分を貯める。サバンナではバオバブの水量が、人々や動物たちの貴重で安定した資源となっている。その樹齢は長く、平均すると300年~500年で、樹齢が長いと約6,000年と推定される巨木もある。
サバンナゾウは、マリの熱帯北部(北緯 16 度)から南アフリカの温帯南部(南緯 34 度)まで の広い緯度範囲に生息する。山地林、ミオンボ・モパニ林、灌木林、サバンナ、イネ科植物が主体となる草地から乾燥 した砂漠に至る多様な植生環境だけでなく、山の斜面から海浜に至るまでの広い高度と領域範囲に生息している。生息地の生産力や水の利用可能性に応じて、大陸のさまざまな地域で、定住や移動、半移動や放浪に近い移動のパターンを示す。行動圏の広さには、主に多様な生態系における植物の生産性と 人間の活動にも強く関連して極端なばらつきがある。
国境を越えて生息しているため、5つの国にまたがボツワナのオカバンゴ・デルタで、アフリカゾウの子どもが仲間に鼻をすりつけている世界最大の自然保護区「カバンゴ・ザンベジ越境保護区域」では、32万頭以上のゾウが生息し ている
アフリカゾウは長距離を移動することができる。乾燥した生態系で、また、気候条件、例えば季節性 や干ばつなどに対応して自然に長い距離を移動する。サバンナゾウは、メスとその子供たちからなる群れで生活する。群れのリーダーは、最年長で経験豊富なメスが務める。通常、3~10頭の群れを作り、メスとその幼獣が中心であるが、群れが成長すると、数百頭に及ぶこともある。非常に社会的で、採餌場や水場の情報を共有する。また水浴びや泥浴びを好み、これにより体温を調整し、心地よさを得る。メスは定まった繁殖期を持たず、22か月の妊娠期間を経て通常1頭の子どもを出産する。アフリカゾウの赤ちゃんは、生まれた時には既に約100kgもあり、人間でも巨漢の重さである。赤ちゃんは、生後すぐに立ち上がり、母親の後をついて歩く。
| タイに生息するゾウ タイに生息するゾウは主にアジアゾウで、世界で最も大きな陸上動物に属し、その体重は最大で6,000kgに達する。アジアゾウは社会的な動物で、特にタイのゾウは非常に知的で、道具を使ったり、問題を解決する能力がある。 彼らは家族単位での強い絆を大切にし、母親とその子供たちを中心にした家族グループが一緒に生活し、協力して食料を探したり、子育てを行う。彼らは主に草や木の葉を食べ、1日に約150kgを消費する。また、非常に知能が高く、様々な音や仕草でコミュニケートする。 タイの歴史では象に乗る王たちの伝説が豊富にあり、特に有名なのが、アユタヤ王朝時代(1351年 - 1767年)、象は戦場で重要な役割を果たし、王たちは通常、象に乗って戦いに臨む。象に乗ることは、王の権威と権力の誇示でもあった。 現在では、象乗り(エレファントライディング)のアクティビティーは観光客に大変好評で、タイの各地の観光地が全力を挙げて取り組んでいる。現在、1,300頭以上の象が象乗りに携わっており、その収益の一部は象の餌や飼育費用に充てられている。象は1日に150kgの食料と10ℓの水を必要とするため、その負担は大きい。 タイ政府は象の保護に積極的であり、保護センターや象の病院の建設、年老いた象のための保護施設など、ゾウの保護と繁殖プログラムが進められている。これにより、適切なケアを受けながら、健全な生活を送っている。 |
 |
 |
 |
オスは、大人になると群れを離れ、単独で生活するか、他のオスたちと小さな群れを作る。繁殖期になると、オスはメスの群れに近づき、メスを巡って争うが、これにより、繁殖の機会を増やすことができる。彼らは広範囲に移動し、食物や水を探し、時折、他の雄と同盟を結び、協力して行動することもある。アフリカゾウの雄は、その大きな体と力強さを活かして、群れの安全と繁殖に貢献する。
しかしながら、アフリカゾウは、密猟や無秩序な開発による生息地の減少などにより絶滅の危機に瀕している。サバンナゾウとマルミミゾウの両方を合わせた大陸全体の個体 数を 415,428頭と推定し、その2006 年以降、大陸全体でおよそ 111,000 頭減少 したと2021年に報告されている。同様の期間に、分布域の約 90%を調査すると、サバンナゾウが 30%減少していることが報告された。
マルミミゾウとサバンナゾウは、かつては同じ種だと考えられていたが、最近の研究によって、別々の種であることが分かり、彼らは、遺伝的に異なっており、そのため、マルミミゾウとサバンナゾウは、互いに異なる独自の進化を遂げてきたと考えられている。
耳の外縁が丸みを帯びて比較的小さな耳が名前の由来となっている。マルミミゾウは湾曲しない短い牙を持ち、体長が約4〜6m、肩高が約2.4mで、体重は約2,700〜6,000㎏、サバンナゾウ(全長6~7.5m、肩高はオスが3~4m、メスが2.2~2.6m、体重はオスが4.5~7トン、メスが2~3.5トン、尾長は1~1.5mで、性的二型が顕著)の亜種とされていたが、近年では別種として扱われいる。アフリカ中部の熱帯雨林に生息し、ミネラルを補うために岩塩や泥を食べることもある。また、シロアリ塚やミネラル・リックmineral lickと呼ばれる場所で塩などのミネラルを補う。
ミネラル・リックは、家畜が舐める(lick)ことができるように農民が牧草地に置く塩の塊であることもあるが、自然界のリック lickは普通にあり、 ここでは基礎的な必須元素 が得られる。 例えばリンとナトリウム、カルシウム・鉄・亜鉛などの微量元素 trace elementsがあり、 象や鹿などの野生動物の骨、筋肉などの成長期の春には欠かせない。このようなリックは、必須元素が取り込められない一般に貧弱な生態環境ではとりわけ重要である。 激しい天候が塩分を含んだミネラルの埋蔵ヵ所を露頭させ、 これが何マイルも離れた動物を引き寄せる。ある種の動物はソールト・リック salt lickの中のカルシウムを検知できると思われている。
マルミミゾウの牙は細く短く、下向きに曲がっている。これは、密林の中を移動する際に、邪魔にならないように適応した結果だと考えられている。主に果物・樹皮・葉などを食べ、森林の中を歩き回り、地面に落ちた果物や、木の葉、樹皮などを探す。長い鼻を使って、器用に食べ物を口に運ぶ。
マルミミゾウは、数頭のメスとその子供たちからなる最大10頭前後の小規模な群れで生活する。群れは、血縁関係のあるメスとその子供たちで構成され、リーダーは、最も年上で経験豊富なメスが務める。群れのメスがほかの群れに移動するなど、交流は非常に柔軟と言う。
オスは、大人になると群れを離れ、単独で生活するか、他のオスたちと小さな群れを作る。群れからはぐれた若い個体は、ライオンやブチハイエナからの捕食の危険性があるが、生息地が被ることは余りなく捕食は稀と言う。
繁殖に季節性はないが、オスは繁殖の準備ができるとムストmusthと呼ばれる状態になり、この時期にはホルモンの変動が起こり、非常に攻撃的になったり、行動が変わったりする。他のオスとの争いや、メスへの関心が高まるため非常に活発になり、メスに積極的にアピールをする。この間、オスの側頭腺からは強烈なにおいがする分泌物が流れ出し、オスはそれを木にこすりつけたり、耳をはためかせたりして拡散させる。 また、尿も垂れ流し状態となり、攻撃性が高くなる。ムストは若齢個体だと数日から数週間で終わるが、年を増すにつれて長くなり、数か月になることもある。オスはメスの群れに近づき、メスを巡って争う。
繁殖形態は胎生で、1回に1頭の幼獣を産む。繁殖スピードは、マルミミゾウのほうがサバンナゾウよりもやや遅いようである。メスは約22か月の妊娠期間ののち、100㎏の赤ちゃんを1頭産む。出産間隔は5~6年でメスは生涯に約4頭の子供を産む。 赤ちゃんはほかの群れのメスからも育てられ、4~5歳で完全離乳するが、その後も数年周りの世話を受ける。性成熟にはオスもメスも11~14歳ごろであるが、実際の繁殖は23歳ごろから始まるようだ。また、オスは性成熟に達するころ、群れを離れる。
マルミミゾウは、喉の奥にある特殊な構造を使って低周波音を発信する。この低周波音は「インフラサウンド」と呼ばれ、人間の耳には聞こえないほど低い周波数(20Hz以下)で、この音を使って、遠く離れた仲間とコミュニケーションを取る。具体的には、ゾウの喉の奥にある「喉頭嚢(こうとうのう)」という空洞が共鳴器として機能し、低周波音を増幅する。この音は地面を伝わり、最大で10km離れた仲間まで届けることができる。この低周波音は、仲間の位置を知らせたり、危険を警告したり、繁殖期のメスを引き寄せるために使われる。ゾウの社会生活において非常に重要な役割を果たしている。
さらに、ゾウは足の裏でもこれらの低周波音を感じ取ることができ、地面を伝わる振動を通して情報を受け取ることができる。この驚くべき能力は、ゾウが広範囲にわたって情報を共有し、群れの安全を確保するのに役立ってている。
果実を主食とし、日々長距離を移動するマルミミゾウは森林の生態系において重要な役割を果たしており、その糞には、植物の種子がたくさん含まれており、森のあちこちに種子を運ぶ役割をしている。糞は、土壌を豊かにする効果もあり、マルミミゾウは、「森の庭師」と言える。マルミミゾウは、また植物の種子を遠くまで運んでくれる「種子散布者」である。 食べた果実の中の種子を遠くまで運ぶ。約40時間もの間、ゾウの体内に滞留したのち、糞とともに排泄された種子は、糞を肥料として新たな場所で芽吹きする。果実には外殻が固く、ゾウにしか食べられないものもある。そういった植物にとっては、ゾウはかけがえのない存在となり、このような種はキーストーン種Keystone speciesと呼ばれる。
(キーストーン種は、生態学において、それが生息するコミュニティに不釣り合いに大きな影響を与える種の多くが、頂点捕食者となり、天敵や敵がいない状況に置かれる。このような種は、コミュニティを支配する他の種の個体数を制御することにより、コミュニティ内の地域の生物多様性を維持するのに役立つが、 やがて幅広い種に重要な資源を提供することによって生物群集の他の種もキーストーン種の存在に依存して、群集の構造を維持するようになる。
キーストーン種と要種は、生態系において非常に重要な役割を果たす種であるが、 キーストーン種は、その存在が生態系全体の構造と機能に大きな影響を与える種を指す。例えば、オオカミがキーストーン種である場合、オオカミがいることで草食動物の数が制御され、それによって植生や他の生物の多様性が維持される。
要種は、特定の生態系や生物群にとって特に重要な種を指す。要種がいなくなると、その生態系全体に大きな影響が及ぶことが多い。 例えば、アフリカプラムの果実を食べるマルミミゾウは、アフリカの森林生態系において要種と言えるが、マルミミゾウが果実を食べることで種子が分散され、新たな植物が生える機会が増え、新たな生態系が確立されるためキーストーン種と呼ばれる。
昆布の森の生態系の要種であるラッコは、ウニを消費することで昆布の森の生態系のバランスが維持される。キーストーン種とは、特定の生態系にその個体数に対して極めて大きな影響を与えるが、生態系の全体的な構造と機能にとっても重要となり、その生態系を構成する他の種類の植物や動物全体に影響を与える。)
アフリカの森林地帯には、いくつかのプラムの木が自生している。その中でも特に有名なのは、アフリカプラムPlumieria albaで、この木は美しい白い花を咲かせ、果実も食べられる。このキーストーン・チェリーは、プラムの木から生産される果実で健康にも良い。アフリカプラムの木は、森の動物たちにも食料や住処を提供する。また、森林再生の貢献も大きい。また、アフリカン・プラムは熱帯樹木の一種で、薬用植物としても重宝されている。南・中央アフリカ原産で、主にカメルーン山地や高地に生育している。そのアフリカン・プラムの樹皮は、何世紀にもわたってサハラ以南のアフリカの医療に用いられてきた伝統的な薬草資源である。現地の人々にとっては、膀胱や腎臓の問題、衰弱・炎症・感染症・発熱・性機能障害など、多くの病気の治療薬になっている。このように、アフリカプラムの果実はキーストーン種として、森林生態系のバランスを保ちながら重要な役割を果たしている。
特に森林に適応しているマルミミゾウは、主にアンゴラ・ガーナ・ガボン・カメルーン・コンゴ共和国などの西アフリカから中央アフリカにかけて、標高2,000mまで熱帯雨林にも生息する。野生下では50〜60年生きる。
しかしながら、密猟や生息地の破壊により、絶滅の危機に瀕している。IUCN(国際自然保護連合)のレッドリストでは「絶滅危惧種IA類」に指定されている。
軟質なサバンナゾウの象牙に対し、マルミミゾウの象牙は硬質でより上質とされている。 その象牙のために、多くのマルミミゾウが殺されてきた。また、生息地である森林の伐採により、彼らの生息地はかつての4分の1に限定されている。 さらに、マルミミゾウの個体数の約7割に当たる約9万5千頭が定住するガボンでは、気候変動や降水量の減少などの影響で、ゾウが食べる果実が8割も減少した。このためマルミミゾウが菜園などの農作物を荒らすこともあり、人間との軋轢が高まった。 かつては何百万頭もいたマルミミゾウは、現在サバンナゾウと合わせても約41万5千頭しかいないと推測されている。
過去50万年にわたり、アフリカゾウとマルミミゾウの間で交配した形跡が見られない。アフリカゾウとマルミミゾウは異なる生態系に適応し、それぞれの環境で独自の進化を遂げてきた。また、アフリカゾウはLoxodonta属、アジアゾウはElephas属と異なる属に属しており、しかもアフリカゾウやマルミミゾウは56本、アジアゾウは58本と染色体の数も違うため自然界での交配は確認されていない。
最近の研究によると、密猟の影響で、アフリカゾウの中で牙がない個体が増えている。象牙を狙った密猟が激化する中で、牙を持たないゾウの方が生き残りやすくなり、その結果、牙のない個体が増えている。
モザンビークは、ポルトガルの植民地から独立を果たした1975年以降、不安定な政情が続き、特にモザンビークの内戦中(1977年~1992年)では、象牙を資金源とするために多くのゾウが殺された。この期間中、牙のないメスのゾウの割合が急増し、内戦後もその傾向が続いている。牙はゾウにとって重要なツールであり、木の皮を剥いたり、地下水を掘り当てたりするのに使われる。牙のないゾウが増えることで、これらの行動が変わり、生態系全体に影響を及ぼすことが懸念されている。
ナミビアに生息する象はサバンナゾウで、ナミビアの砂漠地帯に適応した特徴を持っており、そのため「サバンナの巨人」とも呼ばれている。
ナミビアは年間300日が晴天で、6月から8月の冬期はる乾燥し、9月から11月が小雨季、2月から4月が大雨季である。降水量は場所によりさまざまで旱魃も多い。
2024年、過去数十年間で最悪の干ばつに直面。政府は放牧や水供給への負担を軽減するとともに、干ばつの影響で飢えに直面している人々に食肉を提供する目的で、2024年8月28日、野生動物の駆除に着手。カバ・バッファロー・インパラ・ゾウ・オグロヌー・エランド・シマウマなどが犠牲となった。
「ナミビア政府が、干ばつに苦しむ同国で野生動物を食用として捕獲すると決定した。対象は、ゾウ・カバ・バッファローなどの723頭。」
「ハンター企業が既に157頭を捕獲し、56,800キログラム以上の食肉が生産された。ナミビア政府は「国民のため、憲法に定める天然資源の利用だ」と説明している。」
「ナミビアでは食糧備蓄量の84%が枯渇し、半分以上の人口が深刻な食糧不足に直面する可能性が高まっている。政府は「飢饉」への対策として野生動物の捕獲を決めた。」
アジアゾウの牙は、上顎の門歯(前歯)が変化したもので、主に防御や食物の掘り起こし、樹皮の剥ぎ取りなどに使われる。また、インドではオスのアジアゾウを牙の長さや太さで分類することもある。ゾウの個体の識別やその牙の大きさが、そのゾウの社会的地位や年齢を示すことがあり、群れ内での役割や行動に影響を与えることもある。また、ゾウの生態や行動を研究するために、牙の特徴を記録することが重要であり、密猟防止のための監視にも役立つ。
インドゾウの体長は最大6.5m、体重は6.7tで、アフリカゾウに比べて耳が小さく、牙も短いのが特徴で、アフリカゾウの肩高は約3~3.9m、インドゾウの肩高は約2.5~3.2mで、一般的にアフリカゾウの方が大きい。インドゾウは、インド亜大陸や東南アジアに生息している。野生では60〜70年生きることができる。
野生の場合、おとなは立って寝るが、子ゾウはおとなが守ってくれるので横になって眠る。動物園では、まわりに危険がないので、おとなのゾウも横になって眠る。この行動は、飼育環境に慣れていることの証拠にもなる。
 |
 |
| 古代のバリ島は、多くの歴史的な遺産と文化的な遺物で知られている。バリ島はBC1世紀には既に存在していたとされ、インドや中国との交易が盛んであった。特に、バリ・ヒンドゥー文化の影響が強く、多くの寺院や神殿が建設された。 インドネシアの古代ゾウは、主にスンダランドSundalandと呼ばれる地域に生息していた。これらのゾウは、約1万年前に絶滅した。 インドネシアの古代ゾウは、現在のインドやスリランカに生息していたゾウと同じ種であると考えられている。 スンダランドは、現在のインドネシア・マレーシア・フィリピンなどにかつて存在した広大な大陸の一部である。約1万年前の氷期には、現在の海面よりも約100m低く、広大な陸地が広がっていた。この地域は、現在のインドネシアの島々がつながって一つの大きな大陸を形成していた。 スンダランドは、現在のインドネシアの中央部から東部にかけて広がっており、多くの古代文明が栄えた場所でもある。この地域は、氷期の終わりに海面が上昇し、現在のような多島海洋地帯に変わっていった。 ナウマンゾウは、約3万年前にはスンダランドから日本列島に移動し、その後絶滅しました。この移動は、当時の地続きであった東南アジアと日本の間の陸橋を利用して行われたと考えられている。 ナウマンゾウは、日本の更新世の遺跡からも発見されており、特に九州地方や四国地方で多くの化石が見つかっている。このことから、ナウマンゾウが、西日本から環境変化に適応しながら日本列島を北上し広く分布していたことがわかる。 ナウマンゾウPalaeoloxodon naumanniは、約1万5,000年前までは日本列島に生息していた象で、ナウマンゾウは、現生のアジアゾウElephas maximusと近縁であり、特にインドゾウElephas maximus indicusと遺伝子上の関連が深い。 ナウマンゾウは、約30万年前に、ユーラシア大陸からと言うよりも、スンダランド陸橋から日本列島に渡来し、次第に寒冷な気候に適応しながら進化した。 現在でも、インドゾウの本流は、インドや東南アジアに生息しており、熱帯と温暖な気候に適応しながら進化し分化を重ねた。ナウマンゾウとインドゾウは、共にその進化の過程で分岐したと考えられている。 ナウマンゾウの化石は日本各地で発見されており、特に長野県の野尻湖周辺では、長期的に連続する多数の化石が出土し、 ナウマンゾウとインドゾウは共通の祖先を持ち、それぞれ異なる環境に適応して分化し進化してきた。 |
アジアゾウは、アフリカゾウと並ぶ、2種類のゾウの一種であるが、アフリカゾウと比べて、体が小さく、耳も小さいのが特徴である。また、アジアゾウのメスは、牙を持たないか、持っても非常に小さい。その一方、非常に知能が高く、しかも社会性も発達している。彼らは、複雑なコミュニケーションを取り、群れで協力して生活する。彼らは、古くから、人間と、深く関わってきた。彼らは、労働力として、木材の運搬や、農作業などに、利用されてきた。また宗教的な儀式や、祭りにも、重要な役割を果たしてきた。しかし、近年、アジアゾウは、絶滅の危機に瀕している。彼らの数は、生息地の破壊や、密猟によって、減少し続けている。アジアゾウを守るためには、彼らの生息地を守り、密猟を阻止することが、重要である。
アジアゾウは草食動物で、草・木の葉・果実・樹皮などを食べる。1日に、約150kgもの植物を食べるので、食べることに、多くの時間を費やす。彼らは、長い鼻を使って、地面の草や、木の葉を、器用に巻き取って食べる。また、牙を使って、木の皮を剥がしたり、地面を掘って、根っこを食べたりすることもある。さらにアジアゾウは、農作物を食べることもあり、農家の人々との間で、トラブルになることもある。
アジアゾウは、様々な方法で、コミュニケーションを行う。彼らは、ラッパのような大きな声で鳴いたり、鼻を鳴らしたり、足で地面を stamping したりすることで、様々な音を出し、コミュニケーションを取る。彼らは、耳を動かしたり、頭を振ったり、鼻を上げたり、体同士を触れ合ったりすることなど、体の動きで、意思の疎通を図る。また、尿や糞の匂いで、コミュニケーションを図る。例えば、尿にはホルモンが含まれている。それで繁殖期の情報や健康状態などが伝えられる。また、糞の匂いは個体識別の役割も果たす。どの象がその地域にいたかを示す。こうした匂いを嗅ぎ分けることで、インドゾウたちはお互いの位置や状態を把握している。
これらの方法を、組み合わせて、複雑なコミュニケーションをとっていると考えられている。
アジアゾウは、IUCN(国際自然保護連合)のレッドリストで、「絶滅危惧種(EN)」に指定されている。
彼らの数は、生息地の破壊や、密猟によって、減少し続けている。アジアゾウを守るためには、彼らの生息地を守り、密猟を阻止することが、重要である。
アジアの森林を、悠然と歩くアジアゾウは、知能が高く、社会性も発達した動物として知られている。古くから人間と共に生きてきた彼らは、アジアの文化と密接に関係している。
インドゾウは、アジアゾウの亜種の一つ。古くから人間と共に生きてきた彼らは、インドの文化に深く根付いている。肩高は約2.7〜3.5m、体重は3,000〜5,000kgほどで、アジアゾウの中では中くらいの大きさである。オスはメスよりも大きく、立派な牙を持っている。野生下では60〜70年生きる。アジアゾウの亜種は、インドゾウの他に、スマトラゾウ・セイロンゾウが現存し、さらにボルネオゾウは別亜種とする学説もある。
熱帯雨林や乾燥落葉樹林、草原など、様々な環境に適応しているインドゾウは、アジアゾウの亜種の一つで、インド・ネパール・ブータン・バングラデシュに生息している。彼らは、アフリカゾウに比べて、体が小さく、耳も小さい。また、額には2つのコブがあり、オスの牙は、アフリカゾウほど大きくはない。インドゾウは、古くから人間と深く関わってきた。彼らは、労働力として、農業や林業、輸送などに利用されてきた。また、宗教的な儀式や祭りにも、重要な役割を果たしてきた。
インドゾウは草食動物で、草、葉、木の枝、樹皮、果実などを食べる。彼らは、1日に約150kgもの植物を食べるので、食べることに多くの時間を費やす。
また、塩分を補給するために、地面を掘ってミネラルを含む土を食べることがある。農作物を食べることもあり、農家の人々との間で、トラブルになることもある。
インドゾウは、メスとその子供たちからなる群れで生活する。群れのリーダーは、最も年上で経験豊富なメスが務め、群れをまとめ、安全な場所へ導く。
オスは、大人になると群れを離れ、単独で生活するか、他のオスたちと小さな群れを作る。繁殖期になると、オスはメスの群れに近づき、メスを巡って争う。
インドゾウは、ラッパのような大きな声で鳴いたり、耳をパタパタさせたり、鼻を上げて匂いを嗅いだり、体同士を触れ合ったりして、コミュニケーションを取る。彼らは、様々な音や動作を使い分けて、仲間とコミュニケーションをする。
インドゾウは、知能が高く、穏やかな性格なので、訓練することができる。彼らは、古くから、木材の運搬や、農作業、そして観光客の乗馬など、様々な仕事で活躍している。
しかし、近年では、動物福祉の観点から、ゾウを労働力として利用することに対する批判が高まっている。
インドゾウは、IUCN(国際自然保護連合)のレッドリストで、危急種に指定されている。彼らの数は、生息地の破壊、密猟、そして耕作地の拡大・植林など人間との衝突などによって、減少している。インドゾウを守るためには、森林の保護、密猟の取り締まり、そして人間との共存のための取り組みが必要となっている。
インドゾウは、インド・ネパール・ブータン・バングラデシュなどの南アジアに分布している。ネパールとブータンに生息するインドゾウは、主に森林地帯や草地に生存するが、広範囲に移動する。これらの地域は、彼らにとって重要な生息地となっている。
ネパールでのサファリSafari(スワヒリ語で「旅」の意味。狩猟・探検のための旅行を指す)ではチトワン国立公園へ行くのが一般的で、カトマンズ→チトワン国立公園→ポカラと周遊するのがネパール旅行の定番となる。チトワンで見られる動物はサイ・アジアゾウ・ベンガルトラ・ヒョウ・ナマケグマ・ワニなどでシカは4種類、サルは2種類生息している。チトワンでは、ゾウに乗ってサイを探すエレファントサファリがハイライトで、このサファリは世界的にみても珍しい。
インドゾウは母系社会を形成し、雌とその子供たちが群れを作る。雄は成熟すると群れを離れ、単独で生活する。草・葉・果実・樹皮などを食べる。1日に大量の植物を摂取し、その活動を通じて生態系に大きな影響を与える。「生態系エンジニア」とも呼ばれ、植物の種を運んだり、森林の構造を変えたりすることで、他の動物の生息環境を整える役割を果たす。インドゾウは絶滅危惧種に指定されており、個体数の減少が懸念されていが、互いの生息地の破壊が人間との衝突の主な原因となっている。
インドゾウはその知恵と力強さで知られ、人間社会とも深い関わりを持っている。彼らの保護と共存が重要な課題となっている。
| スリランカゾウCeylon elephan アジアゾウElephas maximusは、新生代第四紀更新世ジェラシアン期の約200万年前に登場したとされている。この時期に、彼らの先祖は徐々にアフリカからアジアへ移住し、それぞれの環境に適応して特有の形態に進化させてきた。 スリランカゾウはアジアゾウの亜種であるが、他のアジアゾウよりも小型で、特異な鼻の形状をしている。約2,500年前から人間と共存してきた。 |
 |
 |
 |
 |
 |
|
カンボジアゾウなどの野生のアジアゾウは、主に夜行性で、草・樹皮・果実などを食べるために長い距離を移動する。象は非常に社交的な動物で、母親が子象をしっかりと守る。 カンボジアゾウもアジアゾウであるため、比較的温厚で人に慣れやすいため、アジアの国々と同様に古くからアジアゾウを飼いならし共に生きてきた。 カンボジアの遺跡建設には、ゾウが重要な役割を果たしていた。特に、アンコール・ワットやバイヨンなどの古代都市の建設には、ゾウが使われていたと言われている。ゾウは、重い石材を運ぶために使われ、建設現場での重要な労働力として活躍していた。 |
 |
 |
 |
スリランカゾウの肩高は約2.5〜3m、体重は3,000〜5,500kgほどで、アジアゾウの中でも、大型である。オスはメスよりも大きく、立派な牙を持つことが多い。メスは、牙を持たないか、または、非常に小さい。スリランカ島のみに生息する、固有種で、かつては、スリランカ島全域で生存していたが、現在では、その数は激減し、生息地は限られる。彼らは、乾燥した森林地帯や、草原地帯を好むが、水辺に近い場所を、住処とすることもある。野生では約60〜70年生存する。
牙は、象牙として、高値で取引されるため、密猟の対象となっている。そのため、牙を持つオスは、密猟によって、殺されることが多く、その結果、スリランカゾウの性比は、メスに、偏っている。そのため繁殖に、悪影響を及ぼす。密猟を阻止することが、スリランカゾウの保護にとって最大の課題になっている。
スリランカゾウは、様々な方法で、コミュニケーションを行う。ラッパのような大きな声で鳴いたり、鼻を鳴らしたり、足で地面を stamping したりするなど、様々な音を出し、コミュニケーションを取る。また、耳を動かしたり、頭を振ったり、鼻を上げたり、体同士を触れ合ったりするなど、体の動きで、コミュニケートする。尿や糞の匂いでも、コミュニケーションをする。これらの方法を、組み合わせることで、複雑なコミュニケーションを取っている。
スリランカゾウは、古くから、人間と、共存してきた。労働力として、木材の運搬や、農作業などに、利用されてきた。また、彼らは、宗教的な儀式や、祭りにも、重要な役割を果たしてきた。しかし、近年、スリランカでは、人口増加や、開発が進み、スリランカゾウの生息地が、狭められている。そのため、彼らは、食料を求めて、人里に近づくことが増えて、その結果、人間とスリランカゾウの軋轢が絶えない。
スリランカゾウでも現在、絶滅危惧種に指定されている。かつては4万頭以上が生息していたが、植民地時代の狩猟や生息地の開発.による減少により、1970年には約2,000頭まで減少した。現在は保護活動の成果もあり、約5,000頭まで回復しているが、密猟や人間との衝突により年間約100頭が犠牲になっている。
スリランカでは、ゾウと人間の共存が大きな課題となっている。ゾウが人間の生活圏に侵入しないように電気柵が設置されているが、それでも完全に防ぐことは難しい。また、ゴミの不法投棄が原因で、ゾウが病気になるケースも増えている。
スマトラ島の緑深い森に、静かに暮らすスマトラゾウは、アジアゾウの中でも最も小型で、その愛らしい姿には魅了される。観光客はスマトラゾウに餌をあげたり、ブラッシングをして「お世話を体験」もすることができる。また、スマトラゾウと一緒に川で水遊びを楽しみ、彼らの自然な行動を間近で観察できる。一部の保護区では、ゾウと一緒にジャングルをパトロールするツアーもある。これはゾウの保護活動の一環で、観光客も参加することにより、スマトラゾウの保護活動や現状について学び、より理解を深めることができ、しかも観光客にとっても貴重な体験となる。
今、アジアゾウの中でスマトラゾウは、その名の通りインドネシアのスマトラ島にのみ生息するアジアゾウの亜種の1つ、しかし最も高い絶滅の危機に直面している。
インドネシアのスマトラ島に生息するアジアゾウの亜種スマトラゾウも、絶滅の危機に瀕している。スマトラゾウの推定個体数は1,700頭。
その危機の最大の原因は、今も続く島の熱帯林の消失です。IUCN(世界自然保護連合)も、絶滅が心配される世界の野生生物のリスト「レッドリスト」の中で、このスマトラゾウの危機ランクを最高位の近絶滅亜種(CR)へと引き上げ、その未来を警告、生息環境である森の保全が急がれる。
豊かな生物多様性に恵まれた、世界的に貴重な熱帯林が今も残るインドネシアのスマトラ島。その森は、人々の生活の基盤となるとともに、アジアゾウ・トラ・スマトラサイ・オランウータンなど、多くの希少な大型野生動物の生息地でもある。
しかし、急速な森林破壊と生息地の減少で、1985年以降、スマトラ島の森林は半減し、ゾウの生息地が大幅に減少した。熱帯林は製紙企業による森林の伐採と、アカシアやユーカリといった紙の原料となる木の植林地へと換えられた。また、パーム油を生産するためのアブラヤシの大規模農園の開発や、違法な居住地や農地の開発も、熱帯林を大規模に劣化・減少させる原因になっている。これにより、ゾウたちは食料や水を求めて人間の居住地に近づくことが増え、結果として人間とゾウの軋轢が増加した。
ボルネオゾウ 約30万年前のボルネオ島(インドネシア、マレーシア、ブルネイ)の環境については、現在のような熱帯雨林が広がっていたと考えられている。この時期は新生代第四紀更新世の氷河時代であれば、ボルネオ島も氷期と間氷期の影響を受けており、気候は現在よりも涼しく乾燥していた可能性が高い。そのため、森林の構成や動物の分布も現在とは異なっていた。
しかもボルネオ島は長い間、豊かな生態系を維持しており、古代から多様な動植物が生息していた。インドネシアの中部に位置する大きな島インドネシアのスラウェシ島だけでも、2021年現在、その過去70年間におよそ300の洞窟で壁画が確認されている。その中には、2番目に古い具象的な洞窟壁画も含まれている。4万4000年以上前に描かれたこの壁画は、5~10cmほどの小さな人間らしき姿がイノシシや小型の水牛の仲間を追う場面が描かれ、太古の狩りの様子が生き生きと表現されている。
ボルネオ島の洞窟にも古代の壁画が残されており、これらの壁画は約4万年前のものとされている。これらの壁画は、当時の人々がどのような動物と共に生活していたかを示す貴重な史料となる。解析が進めば、インドネシア各地で発見された洞窟壁画の価値をさらに高めることになった。
約30万年前のチバニアン期(約77万年前から12万年前まで)のボルネオゾウの環境は、現在とは大きく異なっていた。この時期は、地球の気候が比較的温暖で、多雨の環境が続いていた。ボルネオ島は、現在よりも広い陸地が存在し、森林が広がっていたと考えられている。
約30万年前のボルネオゾウの環境も、現在のようにボルネオ島の一部でした。当時の気候は比較的湿潤で、熱帯雨林が広がっていた。ボルネオゾウは、この豊かな森林環境の中で生息していた。彼らは主に草や木の葉を食べて、他の大型動物とも共棲していた。生態系は非常に多様であった。
ボルネオゾウElephas maximus borneensisは、アジアゾウの亜種とされている。遺伝子研究によれば、ボルネオゾウは約30万年前に、ボルネオの独特の環境に適応して、小型化し、特徴的な行動や形態を持つようにいたった。ボルネオゾウが小型化したのは、主に環境の変化と食料資源の競争によるもので、ボルネオ島の熱帯雨林は、他の動物たちと共に生息しているため、食料資源が限られている環境で生き残るために、小型化が進んだと考えられている。
約30万年前のボルネオ島には肉食動物が存在していた。特に、サーベルタイガーやマカイロドゥスなどの大型肉食動物が生息していた。ボルネオゾウは、このような肉食動物から身を守るために、群れで行動するなどの戦略を取っていた可能性がある。この時期の生態系は非常に複雑で、しかも多くの生物が共棲していた。小型化はエネルギー効率の向上や運動能力の増強にも寄与し、生存率を高める効果があった。ボルネオゾウは環境に適応するために進化し、小型化してきた。
遺伝的には、他のアジアゾウの亜種のインドゾウ・スマトラゾウ・スリランカゾウと非常に近い関係にあるが、いくつかの特徴的な違いがある。ボルネオゾウは、他のアジアゾウに比べて体が小さく、耳が大きく、尾が長い。また、ボルネオゾウの生息地はボルネオ島の北東部に限られており、現在では約1,000頭しか残っていないとされている。そのため、保護活動が極めて重要になる。
他のアジアゾウとは遺伝的に異なることが確認されている。ボルネオゾウの雄の牙は、長さが0.5〜1.7m、重さは約15kgにもなる。他のアジアゾウよりも約30%小さい、大人のオスは身長1.7〜2.6m、メスは1.5〜2.2mで、体重は平均2,500kgほどで、小型で、丸みを帯びた体つきをし、顔が小さく、尾が長く、牙はまっすぐに伸びている。
主にボルネオ島の北東部に限られた地域に生息しており、その生息地は急速に減少している。生息地の減少は、アブラヤシ農園の拡大や大規模な開発が原因である。もう1つは、象牙目的の密猟で、ボルネオゾウの象牙は、高値で取引されるため、密猟の対象となる。彼らの未来を守るためには、生息地の保全と、密猟の取り締まり強化が不可欠である。
群れで行動し、通常は家族単位で移動する。 ボルネオゾウは非常におとなしい性格を持ち、長い尾と比較的大きな耳が特徴です。2003年のDNA調査により、他のアジアゾウとは遺伝的に異なることが確認された。野生下では50〜60年生きることがでる。
性格は比較的穏やかで、草食動物で、人間を襲うことは稀、草・果物・樹皮・若芽などを食べ、1日に約150kgの植物を食べるので、食べることに多くの時間を費やす。また、塩分を補給するために、地面を掘ってミネラルを含む土を食べることがある。また川や沼などの水辺を好み、水浴びをしたり、水を飲んだり、木陰で休んだり、果実を食べたりする。
ボルネオゾウのメスは、約22ヶ月の妊娠期間を経て、1頭の赤ちゃんを産む。赤ちゃんゾウは、生まれた時は、体重が約90㎏もある。ボルネオゾウの赤ちゃんは、生まれてから数時間以内に立ち上がり、歩き始める。これは自然界で生き延びるために重要な能力となる。母親ゾウは非常に献身的で、赤ちゃんをしっかり守り育てる。母親は赤ちゃんの世話をするために、群れの中でも特別な役割を果たし、群れ全体で赤ちゃんを保護することもある。赤ちゃんは、母親や他のメスゾウに守られながら、成長していき、そして、約2年間、母親のミルクを飲んで育つ。
ボルネオゾウは、様々な方法でコミュニケートする。彼らは、ラッパのような大きな声で鳴いたり、耳を動かしたり、鼻を上げて匂いを嗅いだり、体同士を触れ合ったりして、感情や情報を伝達する。
また、彼らは、足の裏を使って、地面の振動を感知し、遠く離れた場所にいる仲間とコミュニケーションを図る。
ボルネオゾウが絶滅の危機に瀕している原因は、主に2つ、森林伐採と農地開発による生息地の破壊である。
目次へ
| マーチソン フォールズ国立公園 ウガンダ北部地域の美しい山と言えばエルゴン山、ウガンダとケニア国境に跨がる楯状火山 。東アフリカ大地溝帯に位置し、雄大な景観が広がる火山。その円錐形の山容は、多くの登山家や観光客を魅了する。 エルゴンという名は南麓の洞窟に住んでいたとされるエルゲヨ族(ケイヨ族)に因む。 |
 |
 |
 |
ウガンダはアフリカ大陸の東部に位置し、北に南スーダン、西にコンゴ民主共和国、南はタンンザニア・ブルンジ・ムワンザ、そして東にケニアと接している。国土すべては内陸にあり、それでも、南部にビクトリア湖、西部にアルバート湖、中部にキオガ湖などの大きな湖があり、水資源は豊富である。一般にはナイル川はヴィクトリア湖を源流とする約5,760 kmの大河と思われているが、ヴィクトリア湖には多数の流入河川が存在し、一方でヴィクトリア湖からの流出河川はナイル川しか存在しないため、アルバート湖や
一方、ウガンダの北部地域では、20年に亘る反政府組織「神の抵抗軍(LRA)」の蹂躙によって治安が不安定であったが、近隣国と共同の軍事掃討作戦や米国の支援も背景にLRAはその勢力を縮小し、拠点を国外に移した。北部地域の治安回復に伴い、一時は200万人近くに達した国内避難民の大半が帰還し、復興・開発に取り組んでいる。ただし、2016年以降は南スーダン、近年はコンゴ民主共和国の政情不安・治安悪化などにより、特に北西部・南西部に多くの難民が流入しており、2021年6月現在、ウガンダは約150万人の難民の受け入れ国となっている。
(神の抵抗軍Lord's Resistance Army【LRA】は、1987年にジョゼフ・コニー Joseph Kony【1962年 - 】によって結成された。ジョゼフ・コニーは、 ウガンダの反政府勢力神の抵抗軍 【LRA】の指導者。霊媒であると主張し、聖書と十戒に基づく神政政権の樹立を掲げ、ウガンダ北部を中心として1987年以降、少年兵を使って殺人や奴隷化、性奴隷化・強姦など残虐行為を伴うゲリラ闘争を続けていた。主にコンゴ民主共和国東部、ウガンダの北部地域と南スーダンの一部を侵犯していた。)
ウガンダの北部地域の人口はウガンダ4地域中最下位だが、面積は最も大きい。中心都市はグルGuluであるが、他の3地域と同じく行政機能を持たない。主にアルバート湖の北東側に位置し、地形は山地と高原が特徴である。この地域は、豊かな自然資源があり、特に農業と牧畜が盛んで、いくつかの重要な河川が流れており、水資源も豊富である。
ウガンダ北部地域では、20年に亘る内戦の後、ウガンダ政府や支援者により復興活動が行われているが、貧困率は依然として高く、国内の南北格差は深刻である。北部地域では、南スーダンやコンゴ民主共和国へ抜ける交通網も存在し、労働人口の多くが農業に従事していることから、農産物の輸出には有利な条件を備えている。その一方、農業経験や技術の蓄積が不足し生産性が低いために、その優位性を十分に活用しきれていない。現在の復興支援では、市場志向型の農業の促進と生計向上アプローチの両立を目標に掲げているが、
「グル県のチェレレマーケットには、トマトの産地であるカンパラ近郊から木箱に詰められて送られてきたトマトを、小売業者が販売単位の一つであるベースン(たらい)に荷分けする。グル県産のトマトに比べ、他地域産のものは品質が良い。」と評される。
ウガンダ北部地域の難民居住区の61%は子どもと見られている。うち5%の約36,000人の子どもには保護者がいない。さらにLRAに徴用された当時の子ども兵が「汚れた存在」として扱われ、コミュニティ側が帰還後も受け入れに難色を示し、社会統合に大きな障壁となっている。この他にも可視化されていない多くの難題が存在する。例えば、国際社会による「元子ども兵士」の社会復帰支援を行うことは、ホストコミュニティ側から当時の残虐な紛争記憶が障害となりタブー化されている。また海外からの送金を受け取らず、現物援助へ依存している姿勢も目立つ。結局、これらの構造的要因や歴史的背景を基に、北部地域の人々はIDP・難民・ホストコミュニティそれぞれの立場に加えて、女性や子ども、障碍者などに、別の難題が重なることによって、より一層脆弱な立場に置かれるようになるのが現状である。
| ウガンダ はアフリカ大陸の中央、首都はカンパラで、湖や山々が美しい国。サバンナ草原や |
 |
 |
| キオガ湖Lake Kyogaは、ウガンダ共和国の中部に位置する大きな湖である。アフリカ大地溝帯の一部で、アフリカの大地が何百万年にもわたって沈み込み、隆起してできた地形の一部である。キオガ湖は、この地溝帯の東端に位置し、周囲の地形とともに形成された。その自然の美しさは豊かで、湖畔には多くの野生動物が生息している
。 水面標高は914m、水深は平均で約2.4m、その面積は1,720㎢もある。水深は最も深い場所で約5.7m、湖の多くは深さが4m未満と浅く、水深3m未満の水域は全域がスイレンで覆われ、湖岸の大半にはカミガヤツリやホテイアオイが生育している。パピルスは小さい島々の間を漂う浮島を形成している。その湖の浅さと広がりは、生態系にも独特な影響を与えて、いろいろな水棲生物が住んでいる。地域の漁業にとって重要な役割を果たしている。多くの種類の魚や多数のナイルワニも棲息している。湖の周辺にはシタツンガ(偶蹄目ウシ科)が生息している。 (ウガンダのパピルスは古代エジプト王朝で使われてはいない。古代エジプト王朝は、主にナイル川流域で生産された) この地域は、火山活動や地殻変動が盛んで、湖の周囲には火山の噴火口や溶岩台地が見られる。地溝帯と湖の関係は、地形の形成過程や地質学的な特徴に深く関わっており、この地域の自然環境や生態系にも大きな影響を与えている。 |
 |
 |
 |
|
ヴィクトリア湖は、アフリカ大陸の東部に位置するアフリカ最大の湖、面積6万8,000㎢、ケニア・ウガンダ・タンザニアの三カ国に囲まれた世界第3位の広さをもつ国際湖沼で、その広さは九州の2倍もあり、かつては、約400種類の固有種が生息する「ダーウィンの箱庭」と呼ばれるほどの「生物多様性の宝庫」であった。またこの湖は、その美しい景観と豊かな生態系で知られていった。 ヴィクトリア湖は、その広大な面積と青い水面が特徴で、その湖の周りには、緑豊かな丘陵地帯や森林が広がり、特に湖の南端にはキヴィラ湖国立公園がある。この公園は、ヴィクトリア湖の自然の美しさを保護するために設立された。湖の周辺には多くの野生動物が生息しており、特にキヴィラ湖国立公園では、チンパンジーやシロサイ、ヒョウなどが見られる。 ヒグマとホッキョクグマ、スマトラオラウータンとボルネオオラウータンなど、多くの近縁種は交配することが知られている。500万年前から1,000万年前のアフリカにその起源を遡るとされるアフリカゾウでは、これまでその異種交配について解明されていなかった。 アフリカゾウは当初、単一の種だと考えられていたが、2010年になって2種に分かれることが確認された。現生のゾウは3種に分類されている。アジアゾウElephas maximusと、2種のアフリカゾウ、すなわちマルミミゾウLoxodonta cyclotis(シンリンゾウ)およびサバンナゾウLoxodonta africana(ソウゲンゾウ)である。 一方、約10万年前まで欧州の森林地帯に生息していたアンティクウスゾウPalaeoloxodon antiquusという古代の巨大なゾウは、化石上の証拠からアジアゾウに近縁だと考えられてきた。 スペイン南西部のウエルバにあるMatalascañas Trampled Surface(マタラスカーニャのゾウの踏み跡)で発見された連続した足跡の化石から、この地域が後期更新世(12万9000~1万1700年前)に、アンティクウスゾウが新生仔を育てるための生息地と示唆された。この知見を報告する論文が、Scientific Reports に掲載される。 今回、Carlos Neto de Carvalhoたちは、連続した足跡の化石34セットを分析した。これらの化石は、足跡が円形か楕円形かという基準やその他の基準により、アンティクウスゾウの行跡化石と同定された。 しかも、このゾウ化石種はアフリカのマルミミゾウに最も近縁であることが、遺伝子解析で明らかになった。さらに驚いたことに、コンゴ盆地に生息しているマルミミゾウは、現在のアフリカのサバンナゾウよりも、絶滅したアンティクウスゾウの方により近縁なことが分かった。 また、マンモス由来の古代ゲノムについて新たな情報が得られ、総合的な解析が可能になったことで、過去にさまざまなゾウ種やマンモス種が交雑したことも明らかになった。 |
 |
 |
 |
(青ナイルと白ナイルの名前の由来は、この地で合流するスーダン人がエチオピアから流れてくる川のより濃く暗い色を「青」と呼び、より透明度が高く泡立ちもあったビクトリア湖から来る方の川を「白」と呼んだ事に因む。)
ウガンダの地形は非常に多様で、高原・山地・湖沼地帯・熱帯雨林など、気候はそれぞれの地域によって異なり、通常、北部は乾燥しているのに対し、中部と南部は湿潤である。ヴィクトリア湖やアルバート湖、キオガ湖などの大きな湖があり、水資源は豊富で、ウガンダの自然美と生物多様性には感嘆せざるを得ない。
ヴィクトリア滝は「モシ・オ・トゥニャ」つまり「雷鳴の煙」とも呼ばれ、その壮大な水しぶきと轟音は圧倒的で、世界遺産に登録されており、絶景ポイントからの眺めや、デビルズプールでのスリリングな水遊びが魅力的である。
ウガンダの中央部のキオガ湖は、アフリカ大湖沼(アフリカ大陸の大地溝帯にある湖の総称)の一帯に位置している。水深は最も深い場所で約5.7m、湖の多くは深さが4m未満と浅くなり、水深3m未満の水域は全域がスイレンで覆われ、湖岸の大半にはカミガヤツリやホテイアオイが生育している。
大地溝帯Great Rift Valleyは、主にアフリカ大陸を南北に縦断する巨大な谷で、プレート境界の一つである。大地溝帯の谷は、幅35~100km、総延長は7,000kmに亘る、プレート境界の正断層による地殻が割れ、最深部2,211m紅海をはじめ、落差100mを超える急な崖や谷底にできた湖沼などが随所に見られる。アフリカと言えばサバンナをイメージするが、その多くは東アフリカのアフリカ大湖沼の地域にある。キリマンジャロは、タンザニアとケニアの国境に隆起する。
パピルス(カヤツリグサ科の植物の1種)は小さい島々の間を漂う浮島を形成している。北西部のクワニア湖とは増水時にはつながった状態になる。 キオガ湖には46種類の魚類が確認されており、多数のナイルワニも棲息している。湖の周辺にはシタツンガ(シカによく似た体型をしているが、ウシ科の動物)が生息しており、キオガ湖南東部の一部は2006年にラムサール条約登録地となっている。
ウガンダの人口は4,427万人(2019年;世銀)で、主要な言語は英語とスワヒリ語であるが、多くの地方言語も話されている。ウガンダの経済は農業が中心で、コーヒー・タバコ・茶・砂糖などの農産物が主要な輸出品となっている。観光業も重要な産業の一つである。
ウガンダ政府は、国内のサトウキビ農家を保護するために、砂糖委員会の運用を開始した。完全に設立されたとき、委員会は砂糖業界の問題を規制・監視・監督、および仲裁することが期待されている。これにより、農家は、サトウキビの価格の不安定さ、支払いの遅延や不履行など、現在直面している課題から守られる。
2021年のウガンダの砂糖生産量は、ブラウンシュガーが600,000t、工業用砂糖が60,000tであった。これらの量は、2022年の良好な気候条件により、2022年には822,000tに増加すると予想されている。
2021年と2022年上半期のサトウキビの農場ゲート価格は、27米ドル/tであったが、サトウキビの不足と国内および近隣諸国の両方での砂糖の需要が高いため、2024年9月の平均価格価格は42米ドル/トンに上昇した。
ウガンダ政府は「第三次国家開発計画(NDPⅢ)」において、持続的な経済成長に力点を置いて貧困削減を目指しており、その具体的な目標の一つに、良質で生産性の高い社会基盤の整備強化が掲げられている。内陸国であるウガンダにおいては、貨物及び旅客運搬輸送に占める陸上運輸の割合が全体の約96%であることから、持続的な経済成長を達成するには陸上輸送網の整備が必須である。しかし、道路舗装率は国道で20%、道路全体では約4%にすぎず、加えて、経済の中心である首都カンパラ市では深刻化する渋滞が社会経済活動を阻害する要因となっていることや既存道路・橋梁の維持管理面における課題が顕在化しており、これらの課題の解決が急務となっている。
特に東アフリカの内陸に位置するウガンダは、ケニアのモンバサ港、ルワンダ、コンゴ民主共和国東部、南スーダンなどの周辺内陸国・地域をつなぐ北部回廊における中継地として、東アフリカ地域の物流における要衝となっている。北部回廊に隣接する首都カンパラ市の人口は、2023年時点で約168万人、同国の政治・経済の中心地であれば、急速な人口増加及び経済成長に伴う交通量の増大により、主要交差点及び道路で深刻な渋滞が発生しており、同国の経済成長に重大な阻害要因になっている。
国際協力機構Japan International Cooperation Agency(JICA)は、既に2015年9月11日、ウガンダ共和国政府との間で「カンパラ立体交差建設・道路改良事業」を対象として199億8,900万円を限度とする円借款貸付契約に調印している。 本事業は、首都カンパラ市中心部において、特に混雑の激しい主要交差点の立体交差化、交差点改良及び既存道路の拡幅を行い、市内の交通改善と都市交通の促進を図ることにより、大カンパラ都市圏における物流の改善と経済活性化を促進することを目的としている。
持続的な経済成長において質・量ともに安定した電力供給も不可欠である。近年の経済成長を背景に、電力需要は年率約10%増で推移しており、ウガンダ政府は豊富な水資源を利用した水力発電を主とする電源開発を推進し、供給量で一定の成果をあげている。しかし、送変電設備の整備がそれに追いついておらず、頻発する停電や不安定な電圧などによる脆弱性は、産業振興や投資環境整備を図るうえで大きな障害になっている。
一方、地方電化率は現在も10%強であり、このことが地方部における教育や保健などの社会サービスの安定した提供が妨げられている。こうした背景から、安定的な電力供給はウガンダの開発政策の中でも最重点課題の一つとなっており、中長期的な送配電網の整備、地方電化率向上への支援が期待されている。
首都圏カンパラ市外の人口増加率は8%を超えており、無計画に都市圏が拡大する一方、スラム形成が発生している。これらの状況が継続することで、基礎インフラや公共社会サービスの不足・悪化が起きている。上下水道の不備や廃棄物の不適切な処理などに伴う劣悪な衛生環境による都市感染症リスクを含め、都市環境悪化に伴う多くの課題を克服する都市構造及び都市インフラ整備が喫緊の課題になっている。
ウガンダは農業生産に適した土壌を有している。同国の就労人口の72%は農業従事者であり、また輸出総額の33%を農産物が占める。そのため、農村部における所得向上は、ウガンダ全体の経済成長の原動力になる。2021年より第三次5か年国家開発計画(NDP-Ⅲ)にて定められた農業産業化プログラムはコメを重点作物の一つとして定めている。換金作物としてコメの生産は急速に拡大しているが、粗放的な稲作が原因で単位面積あたりの収穫量は乏しい。
需要・生産が増加傾向にある穀物の生産性の向上と品質を高めためには、脆弱層を対象とする生産支援が必要とされている。また、普及員や研究者の稲作に関する知識の練成は、これまでの取り組みの中で一定程度進んできてはいるものの、栽培研究や地域農業調査開発研究所での栽培試験に関しては継続した支援が求められている。
近年、気候変動の影響で農作物の供給が不安定なことから、灌漑施設の整備を充実させ水資源を有効利用することが焦眉の急となっている。しかし、農業関係者には灌漑施設整備やその維持管理のための技術や地域社会それぞれのノウハウの蓄積が極めて拙劣のようだ。
農村部の所得向上のために、ネリカNERICA(米穀)など優良品種の普及を含む稲作の振興を通じて、ウガンダにおけるコメの増産及び生産性の向上と支援、また、ウガンダにおける稲作関連の研究開発機関の能力強化と蓄積された開発成果の技術普及とそのための連携強化などの支援が不可欠となる。
(ネリカNERICAは、NEw RICe for Africaの略で、アフリカの食糧難を改善することを目的に開発されたイネ科の品種を総称する。
アジアイネOryza sativaを母親として、アフリカイネOryza glaberrimaの花粉を掛け合わせた種間雑種から開発された。
1992年にアジア稲とアフリカ稲の種間交雑に初めて成功し、現在では水稲60種、陸稲18種が登録されている。
アジアイネの高収量性と、アフリカイネの耐乾燥性・耐病虫性などを併せ持つ。2008年時点で、陸稲18品種が普及に移されたが、水稲についても普及が始まり、陸稲以上に生産性が向上することが期待されている。)
さらに、灌漑関連の人材育成及び灌漑施設の維持管理体制の構築を通じて、水稲作の生産性及び収益性の向上とコメの増産を通して、「サブサハラ・アフリカSub-Saharan Africa(アフリカ州、則ち、アフリカ大陸とその周辺島嶼のうちサハラ以南の呼称)のコメ生産を2030年までの12年間で倍増する」という目標が掲げられている。
独立以来、クーデターなどが繰り返されたが、1986年のムセベニ政権発足以来、政情は安定している。ムセベニ大統領は、1996年、2001年の選挙で圧勝。2005年には、複数政党制の復活と大統領三選禁止規定の廃止が決定されたことにより、2006年、2011年、2016年及び2021年の選挙でも再選され、長期政権を維持している。
北部地域では、20年に及ぶ反政府組織「神の抵抗軍(LRA)」の活動によって治安が不安定であったが、近隣国と共同の軍事掃討作戦や米国の支援もありLRAはその勢力を縮小し、やがて拠点を国外に移した。北部地域の治安回復に伴い、一時は200万人近くに達した国内避難民の大半が帰還し、復興・開発に取り組んでいる。ただし、2016年以降は南スーダン、近年はコンゴ民主共和国の政情不安・治安悪化などに伴い、特に北西部・南西部に多くの難民が流入しており、2021年6月現在、ウガンダは約150万人の難民の受け入れ国となっている。
独立以来、度重なる内乱により1980年代後半まで経済は混乱したが、1987年以降世界銀行・IMFの支援を得て構造調整政策を積極的に推進し、マクロ経済を安定させた。その後は比較的高い成長率を維持している。2010年、成長と雇用創出に重点を置いた5か年国家開発計画(NDP)を策定し、2010/11年度から実施。その後、2015年より第二次NDP(2015/16-2020/21)を実施、2021年より第三次NDP(2020/21-2024/25)を実施中。第三次NDP期間中に、中所得国入りを目指している。
マルミミゾウとサバンナゾウの交雑種の確認
ウガンダ共和国のキバレ森林国立公園は、ウガンダの南西部に位置し、具体的には、ブニョロ地域のキバレ県にある。このチンパンジーが多数生息するキバレ国立公園へのトレッキングでは、チンパンジー遭遇率がウガンダ国内で一番高いと言われている。他にもバブーンやブルーモンキーなど十数種類のサルを濃密に見ることができる。
標高1500m、緑豊かな山間部にある豊かな自然と多様な動植物が生息する地域として知られている。
キバレ森林国立公園の西方20km先のコンゴ民主共和国との国境の近くにキソロの街がある。キソロの北西にはムタンダ湖がある。その北部から東部は山岳地帯で、キバレ森林国立公園は湿潤常緑多雨林とサバンナからなる.多様な植生が混在する約 795 km2 のフィールドである。同地は本来、サバンナゾウの分布域であるが、近年、 隣国コンゴ民主共和国 からマルミミゾウ(シンリンゾウ)が移入してきている。結果として、 公園内のゾウの個体密度は2001年から2003年までに354 % 増加したことが確認されている。その移入に伴い、サバンナゾウとの交雑個体の増加が確認されている。
21世紀に入り、 アフリカゾウの二亜種と考えられてきたマルミミゾウとサバンナゾウの間に大きな遺伝的分化が確認された。 両種は長期間の遺伝的距離が存在するにも関わらず、その生息地の境界にあたる場所において、交配可能を示す交雑種が確認されている。しかし、 この交雑個体の生態学・ 行動学的な特徴については未だその多くが不明瞭なままである。 いずれも絶滅危惧種に指定されている両種の保全策を考えるために は、交雑個体群の生態を理解し、 今後の生態系に及ぼしうる影響を考えていくことが重視されている。
| キバレ森林公園:3日間のチンパンジー追跡と火口湖トレッキング チンパンジーは獰猛なので、ガイドは麻酔銃を構えて案内をする。 |
|
 |
 |
ウガンダ共和国におけるサバンナゾウとマルミミゾウの交雑については、具体的なデータが少ないが、交雑自体は確認されている。これは、両種が同じ地域に生息しているため、自然に交雑が起こった。現状としては、サバンナゾウの個体数は減少傾向にあり、またマルミミゾウも絶滅の危機に瀕している。アフリカゾウの生活領域の急激な縮小により、両種の交雑がさらに進む可能性があり、保護活動や研究が進められているが、環境の変化や人間社会の影響により、予想される先々の状況は厳しい。
| シタツンガは、ウガンダを含むアフリカ東部の湿地帯に生息する哺乳綱ウシ目ウシ科の動物でありながら、すらっとしたシカによく似た体型をしている。体長120~170cm、体重50~120kg程度の大きさ。生息する地域によって微妙に異なるがメスは明るい茶色の毛色、オスはメスよりも暗い色をしており、背中には白い筋模様がある。 オスには螺旋状の、大きいものだと90cmにもなる立派な角が生えている。 シタツンガは湿地帯や沼地、湖岸などの水辺に生息し、特にナイル川やその支流周辺でよく見られる。主に草食動物で、水辺に生える草や水草までも食べる。 シタツンガは群れを作る傾向があり、数十頭から数百頭の群れを形成することがあり、群れの中では社会的な構造があり、リーダーが存在する。 |
 |
 |
交雑が進むことで、両種の遺伝的多様性が増す一方で、個々の種の特性が薄れる。
陸上動物で最大となる体躯をほこるアフリカゾウは、 象牙を狙った密猟による絶滅のおそれが一層高まっている。1979年は134万頭とされた推定個体数は、2016年にはおよそ42万頭まで減少し、およそ31%までその数を減らしている。このままの勢いで密猟が横行し続けることは、アフリカゾウを絶滅に追い込むことになる、との認識が国際的に広がり、2013年以降、アフリカゾウの生息国だけでなく、象牙市場のある中国・アメリカ・イギリスなどの消費国においても対策強化が進められている。生息地のあるアフリカ各国では、密猟・密輸対策として、パトロールや汚職・水際の取り締まり強化を実施し、象牙市場のある複数の消費国で、需要を抑制しようと、象牙の国内取引を原則禁止とするなどの対策が講じられている。
(象牙市場で消費される象牙の主な用途は、アンティーク家具・楽器・彫刻・工芸品、そして装飾品など、象牙はその美しい見た目と加工しやすさから、長い歴史を持つ高級品に使われてきた。 現代の中国やアメリカ、イギリスなどでは、象牙製品が高級なアクセサリーや家具、楽器などとして人気がある。)
2021年3月に公表された最新版IUCNレッドリストの評価ではサバンナゾウとマルミミゾウの2種に分けられ、サバンナゾウはEN(危機)、マルミミゾウはCR(深刻な危機)とされた。2015年時点で約41万5000頭が生息しており、その70%以上がサバンナゾウのみの生息地である南部アフリカに生息していると推定されている。ボツワナ・ナミビア・南アフリカ・ジンバブエの4カ国のアフリカゾウ個体群は、国際取引を規制しないと絶滅のおそれが生じる可能性があり、国際取引が許可制となっている種のリストに掲載されている。 サバンナゾウ及びマルミミゾウの生息を脅かす要因として、IUCNレッドリストの評価では、密猟と土地利用の変化による生息環境の喪失と断片化があげられている。
この傾向は、従来、あまり影響を受けてこなかったアフリカ南部の一部の個体群においても強まっている。人間による急激な土地利用の変化によってサバンナゾウの生息地の突発的な喪失と切断化が進展しており、分布域のすべての個体群に対する切実な脅威になっている。土地の転換は、経済の発展と技術の進歩によって現在も進む人間の活動域の拡大とそれに伴う農業およびインフラ開発の産物である。このような傾向は、人間とゾウの軋轢に関する報告の増加という形で顕在化している。アフリカで予測されている人口の増加は、今後数十年の間に大陸規模で土地の転換が一気に加速することを示しており、この脅威を増大させる。
「アフリカ諸国の2050年・将来人口〈国連WPP2022〉」によれば
アフリカ諸国は、2050年に25億人へ67%急増する。
2024年のアフリカ総人口14.7億人は、2050年には24.6億人になり、世界人口ピーク年の2086年には36.4億人、そして2101年には39.3億人になると予測されている。
世界各国の2101年時点の総人口規模では、
1位:インド15.2億人(2024年比+6%)
2位:中国7.6億人(同▲47%)であり、
ナイジェリアが
3位:5.4億人(同141%)
と予測されている。
特に鼻の利くアフリカゾウは、家の中に蓄えている水のニオイさえも嗅ぎとり、夜間に人家を破壊する事態も発生している。 また、同じく国立公園周辺の農地では、農作物を食い荒らしたり、移動する際に作物を踏み荒らす被害も頻発し、加えて、それまで嗜好していなかった作物までゾウが食するようになり、新たな問題となっている。
象牙目的の密猟は現在、マルミミゾウの主な死因となるほどで、1970年代に行われた最初の調査から今日に至るまでの調査結果から、多くの場所で密猟の圧力が続いていることを示している。ワシントン条約によるゾウの違法捕殺監視(MIKE)プログラムの一環として収集されたデータによると、2008年から大陸規模で激化した密猟は、2011年にピークに達したものの、一部の地域では依然として高い水準で推移していることが知られる。
生息地の直接的な喪失と切断化を進展させる急激な土地利用は、アフリカゾウの分布域の全体において、一層の脅威になっている。土地の転換は、経済の発展と技術の進歩によって現在も進む人間の活動域の拡大とそれに伴う農業およびインフラ開発の産物である。このような傾向は、人間とゾウの軋轢に関する報告の増加という形で顕在化している。アフリカで予測されている人口の増加は、今後数十年の間に大陸規模な土地の転換を一気に加速させ、アフリカゾウを絶滅させる。
目次へ