C3・CA・CAM植物
TOP 車山お知らせ 車山ブログ 歴史散歩 車山高原野鳥 たびネット信州
|
 車山高原 秋10月 手や頬に あたたかい 太陽の 光線の温もり |
||||||
地球上の植物の約95%はC3植物(シーさんしょくぶつ)で、比較的温暖で湿潤な環境を好む。C4植物(シーよんしょくぶつ)やCAM植物(しーえーえむしょくぶつ)とは異なり、高温や乾燥条件下では光合成の能力が低下しやすい。
 C3植物の光合成能力は高く、短時間で大量に増殖する淡水性のクロレラなどの単細胞緑藻類やイネ・ササ・コムギ・オオムギ・ダイズ・ナタネ・ホウレンソウなど、そして樹木などその占有率は高い。樹木は、光合成の過程で最初に生成される化合物が3炭素化合物(炭素原子が3つ結合している化合物)であることからC3植物に属す。
C3植物の光合成能力は高く、短時間で大量に増殖する淡水性のクロレラなどの単細胞緑藻類やイネ・ササ・コムギ・オオムギ・ダイズ・ナタネ・ホウレンソウなど、そして樹木などその占有率は高い。樹木は、光合成の過程で最初に生成される化合物が3炭素化合物(炭素原子が3つ結合している化合物)であることからC3植物に属す。C3植物がC4植物よりも多い理由は、 C3植物は、光合成の過程で二酸化炭素を直接カルビン回路に取り込むことにある。進化の歴史:の中で地球上で最も古い光合成の形態であり、約3億年前には登場していた。そのため、C3植物は先駆的に広範囲にわたって進化し、多様な環境に適応する能力を獲得した。しかも、 C3植物は温帯地域や湿潤な環境により高度に適応した。その環境こそが地球上に広く分布しているため、C3植物の占有率が高まった。更にC3植物の光合成の効率は、低光強度や低温条件下でも優位に働いた。これにより、さまざまな環境で生育する耐性を獲得した。
C3植物はC4植物に比べて光合成に必要なエネルギーや器官が少ないため、より少ない資源で生育できた。 一方、C4植物は高温・乾燥地域に適応し、その光合成効率が高いが、特定の環境に限られるため、C3植物ほど広範囲には分布しなかった。
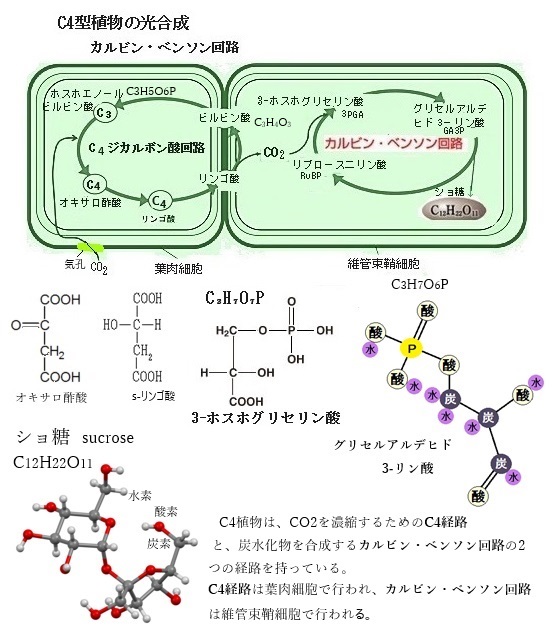
C3植物が、光合成の過程で最初に生成される安定した化合物が3-ホスホグリセリン酸(3-PGA)であることからその名が付けられている。これに対し、光合成のCO2初期固定をC4ジカルボン酸回路により行う植物をC4植物、そしてベンケイソウ型有機酸代謝を行う植物をCAM植物と呼ぶ。
この3-PGAは炭素が3つ含まれているため、「3炭素化合物」と呼ばれる。C3植物の光合成は、カルビン・ベンソン回路を利用して行われる。この回路では、二酸化炭素CO2がリブロース-1,5-ビスリン酸(RuBP)と結合し、3-PGAが生成される。このプロセスがC3植物の特徴である。C3植物の光合成の場所は葉肉細胞で、常の環境下では効率的に光合成を行うが、乾燥や高温の環境ではC4植物やCAM植物に比べて効率が低下しやすい。
CAMはベンケイソウ型有機酸代謝Crassulacean Acid Metabolism (CAM)の略で、CAM型光合成を行う植物をCAM植物と呼ぶ。ベンケイソウ型有機酸代謝(CAM)は、ベンケイソウやサボテン、アロエなどの多肉植物や乾燥地帯に生息する植物が採用する光合成の一形態である。CAM植物は、昼間は気孔を閉じて水分の蒸散を防ぎ、夜間に気孔を開いて二酸化炭素CO₂を取り込み、その夜間に取り込んだCO₂を、昼間なると光合成を行うための材料として使う。
CAMはベンケイソウ科植物に限らず多くの植物にみられる代謝であるが、19世紀にベンケイソウ科植物の葉の酸含量が夜間に増し、昼間に消失する現象が、この科の植物に特有な代謝として認識・研究されたことに由来して慣用的に用いられている。
CAM植物はシダ植物3科5属、裸子植物1科1属(イチョウ科のイチョウ属)、単子葉植物8科98属、双子葉植物33科239属に分布し、ラン科での推定種数7,000種を含め約16,000種にみられる。この数は地球上の維管束植物の約6%に相当する。このように多数の植物にみられるCAMの進化は、環境条件によって誘発された適応によるものと考えられている。CAM植物はCAM型光合成と、葉や茎に水分を蓄える能力を備え、乾燥した環境に適応する多汁性という形態学的特性を併せもつこで、砂漠などの水分ストレスが多発する環境下で生育する植物として進化した。従来、CAM植物のバイオマス生産力(動植物などの生物資源から得られる有機物)はきわめて低いとされてきたが、適切な農業的管理下ではC3植物をしのぎ、C4植物に次ぐ高い能力を示す種もある。
適切な農業管理下では、CAM植物の中にはC3植物を上回り、C4植物に次ぐ高いバイオマス生産力を示す。例えば、リュウゼツランやウチワサボテンは、メキシコやチリの砂漠条件下で高いバイオマス生産を達成している。このようなCAM植物の高い生産能は、昼間は気孔を閉じて水分の損失を最小限に抑えながら、夜間に気孔を開いてCO2を吸収し、昼間になると気孔を閉じたまま光合成を行う光呼吸抑制機能によるものであり、エネルギー効率の面でも優れている。これにより、乾燥地帯や塩害地などの厳しい環境下でも安定したバイオマス生産が期待される。
ニコチン酸を植物に与え、NAD(ニコチンアミドアデニンジヌクレオチド)を合成する経路の代謝物であるニコチン酸を高蓄積させることにより、植物の乾燥耐性およびバイオマス生産性を高められることが発見された。この成果を応用すれば、将来的に農作物の乾燥ストレスを強くする肥料や農薬が開発され、それに伴う収量増産などに貢献できる可能性が高まっている。
また、常にある一定程度のCAM型光合成を示すものを偏性CAM植物、ある要因によりC3光合成からCAM型光合成に切り替わるものをf通性CAM植物と呼ぶ。
目次へ
2)光合成の多様で精妙な進化
夜間は、気孔を開いてCO₂を取り込み、ホスホエノールピルビン酸カルボキシラーゼ(PEP-C)によってリンゴ酸として液胞に蓄える。 暗→明への移行期には、気孔が開いたままリンゴ酸の脱炭酸反応が進行し、CO₂が再放出される。CAMはベンケイソウ科植物に限らず多くの植物にみられる代謝であるが、19世紀にベンケイソウ科植物の葉で酸含量が夜間に増し、昼間に消失する現象が、この科の植物に特有な代謝として認識・研究されたことに由来して慣用的に用いられた。
 CAM植物の進化は、環境条件によって誘発された収斂進化によるものと考えられている。CAM植物は、CAM型光合成という生理機能と多汁性という形態学的特性を併せもつのは、砂漠や水分ストレスが多発する着生環境下で生育する植物に多くみられることである。
CAM植物の進化は、環境条件によって誘発された収斂進化によるものと考えられている。CAM植物は、CAM型光合成という生理機能と多汁性という形態学的特性を併せもつのは、砂漠や水分ストレスが多発する着生環境下で生育する植物に多くみられることである。パイナップルは、パラナ川 と パラグアイ川 の流域が原産地で、熱帯から亜熱帯にかけてのアメリカ原産の多年草のCAM植物である。地下茎からロゼット状に硬い剣のような形の長い葉を多数出し、高さは60~100cm程度に成長する。 パイナップルも、夜間に気孔を開いてCO2を吸収し、リンゴ酸として蓄え、昼間に気孔を閉じて光合成を行うことで水分の損失を最小限に抑える特性を持っている。
光合成を行う単細胞緑藻類は、一般的にC3植物とは異なる。C3植物は、陸上植物の中で最も一般的な光合成経路を持つ植物群で、光合成の最初の生成物が3炭素化合物(3-ホスホグリセリン酸)であることからその名がついている。 単細胞緑藻類は、光合成を行うものの、C3植物とは異なる光合成経路やメカニズムを持つことが多い。緑藻類の中には、光合成の際に二酸化炭素を効率的に取り込むための特別な機構を持つものもある。
例えば、クロララクニオン藻という藻類は、他の緑藻を細胞内に取り込んで共生することで、独自の光合成能力を持つ。このような共生関係は、藻類の多様性を生み出す重要な要素となっている。
南極に生息するナンキョクカワノリは、赤外線を利用して光合成を行うことができ藻類で、通常、植物や藻類は可視光を利用して光合成を行うが、この藻類は赤外線を利用することで、南極の厳しい環境に適応している。植物や藻類が行う光合成は、太陽光に含まれる可視光(350~700nm)のエネルギーを使って水を酸素と水素と電子とに分解し、そこで得られた還元力が二酸化炭素の同化に利用されている。700nmより長波長の光は赤外線であるが、赤外線は可視光に比べてエネルギーが低いため、通常では水を分解することはできない。ただ、一部のシアノバクテリア(最も原始的な酸素発生型光合成生物で葉緑体の祖先と考えられている)では、赤外線を利用した光合成を行うことが知られいる。
C4光合成を行う植物もたくさんあり、しかも単子葉植物に多く、イネ科(サトウキビ・トウモロコシ・モロコシ・ヒエ・アワ・ハトムギ・メヒシバ・ススキ)やカヤツリグサ科(ハマスゲ・テンツキなど)、少数だが双子葉植物でもアカザ科(ホウレンソウやアカザなど)、トウダイグサ科コニシキソウなどがある。
それらは光合成中の二酸化炭素CO2の損失を減らすために進化し、二酸化炭素濃度が低く、また高温・乾燥状態でも光合成が可能となった。そのC4植物の光合成の効率はC3植物よりも高く、それは進化の過程で獲得したCO2濃縮機構や素早い気孔応答に起因する。 C3植物の光合成の最初の生成物がC3化合物(3-ホスホグリセリン酸)であるが、C4植物では、まず二酸化炭素を葉肉細胞でC4化合物(オキサロ酢酸C4H4O5)に固定する。その後、オキサロ酢酸はリンゴ酸に変換され、維管束鞘細胞に運ばれる。
 維管束鞘細胞でリンゴ酸が脱炭酸され、再び二酸化炭素が放出され、カルビン・ベンソン回路に取り込まれる。 葉肉細胞は、葉の内部に位置し、光合成を行う主要な細胞で、その葉肉細胞には多くの葉緑体が含まれており、光合成を行うための主要な場になる。一般的には「葉緑体細胞」という用語はあまり使われないが、葉緑体を含む細胞、つまり葉肉細胞や孔辺細胞など、葉緑体を持つ細胞を総称してこう呼ぶこともある。
維管束鞘細胞でリンゴ酸が脱炭酸され、再び二酸化炭素が放出され、カルビン・ベンソン回路に取り込まれる。 葉肉細胞は、葉の内部に位置し、光合成を行う主要な細胞で、その葉肉細胞には多くの葉緑体が含まれており、光合成を行うための主要な場になる。一般的には「葉緑体細胞」という用語はあまり使われないが、葉緑体を含む細胞、つまり葉肉細胞や孔辺細胞など、葉緑体を持つ細胞を総称してこう呼ぶこともある。葉緑体は光合成を行う細胞小器官で、葉肉細胞や孔辺細胞に存在し、クロロフィルを含み、光エネルギーを化学エネルギーに変換し、光合成に必要なエネルギーを供給する。 これらの違いを理解することで、植物の光合成の仕組みがより明確になる。 葉緑体細胞は、光合成を行うために必要な葉緑体を含む細胞で、光エネルギーを化学エネルギーに変換し、植物が成長するためのエネルギーを供給する。葉緑体細胞は主に葉の表皮細胞や葉肉細胞(ようにくさいぼう)に存在する。
その「葉肉細胞」は、葉の表皮細胞に守られた細胞全体のことを指す。葉肉細胞は葉の緑色を保持し、太陽光を受けて二酸化炭素と水から糖を生成する。このプロセスは、光エネルギーを植物が生きるために必要なエネルギーに変えて植物全体に供給する。
(通常、葉の表皮細胞には葉緑体は含まれていない。ただ、例外として気孔を形成する孔辺細胞には葉緑体が含まれているが、孔辺細胞にある葉緑体は、気孔の開閉に使うカリウム・イオンを孔辺細胞に運び込むのに必要なエネルギーを生産する役割を持っているため、光合成産物の生産には殆ど使われていないようだ。
気孔が、大部分の植物で葉の裏側の表皮細胞にあるは、葉の表側の方が、裏側に比べ、雨が降れば葉の表側は水に濡れ、気孔を通してのガス交換の進行が妨げられるなど、いろんなことが想定されるが、孔辺細胞の葉緑体は光合成よりも気孔の開閉に役立っているため、これには余り照度の高い光を必要としないためでもある。
また表皮細胞が葉緑体を持たない理由は、葉の表面が常に物理的・化学的・生物的な阻害要因が多いためであり、例えば大切な光合成装置である葉緑体が表皮細胞にあると、せっかく合成した光合成産物が昆虫によって食べられる、病菌感染などによる障害を受けやすくなる。むしろ、植物は傷ついた際に再生能力を高め、新しい表皮細胞を復元する方が急務となる。この再生能力は、植物が持つ高い生命力の証(あかし)であり、激変する環境変化に適応するための重要なメカニズムになっている。
通常、表皮細胞には色素の含量は少なく、表皮細胞によって太陽光は余り遮られないようなっている。その一方、いくつかの植物では表皮細胞に光合成に有効でないばかりでなく光合成を阻害する紫外線を吸収してしまう成分を含む種もある。
それでも、いくつかの例外があり、例えば、シダ植物や水生植物、そしてシソ科オドリコソウ属などの表皮細胞には葉緑体が存在する。
最近の研究では、植物の表皮細胞に存在する微小な葉緑体が、病原菌の侵入に対して防御機能を持っていることが明らかになっている。これらの葉緑体は、光合成にはあまり関与しないものの、植物の免疫応答に重要な役割を果たす。シソ科オドリコソウ属の表皮細胞に存在する葉緑体は、主に植物の防御機能に関与し、病原菌の攻撃に応答して細胞内を移動し、この過程で、複数の免疫因子が集積し、病原菌の駆逐している。生物進化の過程は種によりさまざまで、一見真逆な進化を遂げながら、それぞれの種の進化を積み重ねていく。)
目次へ
3)多様化する葉肉細胞の役割
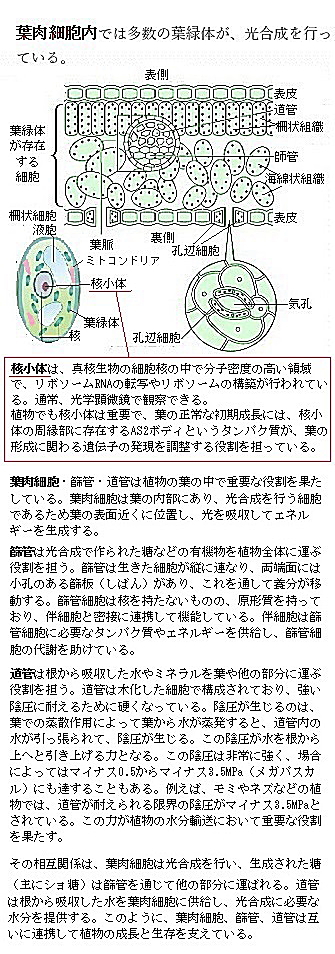
「葉肉細胞mesophyll cell」は、上下表皮組織間に存在する細胞の総称で、葉の上部には柵状(縦)に並んだ葉肉細胞が存在し、これを柵状組織細胞と呼ぶ。また、その下部には海綿状の葉肉細胞が存在し、これを海綿状組織細胞と呼ぶ。柵状組織細胞と海綿状組織細胞の2つ葉肉細胞は、光合成に特化しており、葉の内部で効率的に光を吸収する。
葉肉細胞には、維管束組織は含まないから、一般に被子植物では、主に柵状組織(向軸側)と海綿状組織(背軸側)を形成されている。ただし、イネ科植物などでは柵状・海綿状組織の細胞とまったく異なる、表面が入り組んだ突起armをもつ「有腕細胞armed cell」と呼ばれる葉肉細胞が発達している種がある。特に、稲Oryza sativa( オリュザ・サティブァ)やトウモロコシZea maysなどのイネ科植物において、このような細胞が発達している。
稲は、イネ科イネ属の植物、属名Oryza は古代ギリシア語由来のラテン語で「米」または「イネ」を意味する。種小名 sativa は「栽培されている」といった意味である。
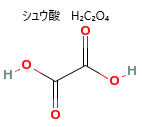 |
| シュウ酸は、多くの植物に含まれる成分で、特にほうれん草やタケノコなどに多く含まれている。 シュウ酸は、えぐ味や苦味の原因となる。 水を多く飲むことで、尿のシュウ酸濃度を薄めることができる。 品種改良でシュウ酸の少ない品種を育種する努力がされており、サラダ用ホウレンソウはその一つ。 |
葉肉細胞には、これらの主要な組織だけでなく、異形細胞や貯水細胞なども含まれる。異形細胞は、特定の環境条件や植物の種類によって異なる形態を持つ細胞で、貯水細胞の方は水分を貯蔵する役割を持ち、乾燥した環境での生存を助ける。例えば、異形細胞は、他の細胞と形や大きさ、含有物が著しく異なる特殊な細胞であって、例えば、タンニンを含むタンニン細胞や、シュウ酸カルシウムの結晶を含む結晶細胞などがこれに該当する。タンニンは植物に広く存在する化合物で、特に葉や果実に多く含まれている。柿の果肉では黒い粒子としてタンニン細胞が見られる。タンニンを含むことで、昆虫や動物の食害を防ぐ。タンニンはタンパク質と強く結合し、消化酵素の働きを阻害する。これにより、捕食者の消化能力を低下させ、栄養吸収が妨げる。また、消化管の内壁に作用し、炎症や損傷を引き起こし、消化不良や下痢などの誘発する。植物のタンニン細胞は、主に液胞にタンニンを蓄積する。これにより、タンニンが植物の他の細胞や組織に影響を与えることなく、効果的に防御機能を果たすことがでる。
タンニン自体は、植物に広く存在する水溶性のポリフェノール化合物の一種で、ワインやお茶には多く含まれており、風味や色に関与する。
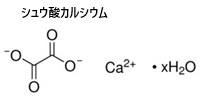
シュウ酸は、多くの植物に含まれる有機酸の一種で、化学式はH₂C₂O₄、植物の成長過程で生成され、カルシウムの輸送や貯蔵、有害物質の解毒などの役割を果たす。その一方シュウ酸はカルシウムと結びつきやすく、過剰に摂取すると腎臓結石のリスクが高まる。特に、腎臓結石の原因となることが多い。シュウ酸カルシウム CaC2O4の結晶を含む結晶細胞は、ほうれん草やレタス、ルバーブやなど植物の葉肉細胞で多くで見られる。摂取すると口や喉に灼熱感を引き起こし、場合によっては消化器障害や呼吸困難を発症する。「シュウ酸を多く含んだ食べ物はできるだけ茹でて食べる」。その結晶自体は、葉や茎などの細胞内に含まれ、植物の防御機構の一部として機能する。
 貯水細胞は、水分を蓄えるための細胞で、特に乾燥地に生育するサボテンやアロエなど多肉植物によく見られる。貯水細胞は、葉肉細胞の一部として含まれることもあるが、貯水細胞は、植物の葉の中で葉肉細胞とは異なる役割を持つ細胞で、水を貯蔵する役割を持ち、特に乾燥した環境で植物が水分を保持するのに役立つ。また貯水細胞の細胞壁は薄く、柔軟性があり、水分を効率的に吸収・保持することができる。
貯水細胞と葉肉細胞は、植物が効率的に光合成を行い、必要な水分を保持するために協力している。貯水細胞は大きな液胞を持ち、多量の水分を蓄えることができ、これにより、長期間にわたって乾燥した環境でも生存が可能になる。貯水細胞は多糖類からなる粘液を含んでおり、この粘液が水分を保持するのに役立っている。
貯水細胞は、水分を蓄えるための細胞で、特に乾燥地に生育するサボテンやアロエなど多肉植物によく見られる。貯水細胞は、葉肉細胞の一部として含まれることもあるが、貯水細胞は、植物の葉の中で葉肉細胞とは異なる役割を持つ細胞で、水を貯蔵する役割を持ち、特に乾燥した環境で植物が水分を保持するのに役立つ。また貯水細胞の細胞壁は薄く、柔軟性があり、水分を効率的に吸収・保持することができる。
貯水細胞と葉肉細胞は、植物が効率的に光合成を行い、必要な水分を保持するために協力している。貯水細胞は大きな液胞を持ち、多量の水分を蓄えることができ、これにより、長期間にわたって乾燥した環境でも生存が可能になる。貯水細胞は多糖類からなる粘液を含んでおり、この粘液が水分を保持するのに役立っている。
目次へ
4)葉緑体細胞の機構
葉緑体細胞は、葉緑体を含む植物細胞であり、光合成を行うために特化しており、葉や茎などの緑色の部分に多く存在している。ここが光合成の主要な場となる。 一方、葉緑体は、葉緑体細胞内に存在する小器官で、光合成を行う場所で、その葉緑体は外膜と内膜の二重膜で囲まれており、その葉緑体の内部には光合成色素である葉緑素(クロロフィル)が含まれている。葉緑体は光エネルギーを利用して二酸化炭素と水から酸素と糖を生成する役割を果たす。
葉緑素は、葉緑体の中のチラコイド膜に存在する。チラコイドは、葉緑体内で円盤状に積み重なった構造で、これが集まってグラナと呼ばれる部分を形成している。 葉緑体と葉緑素は、どちらも植物の光合成において重要な役割を果たしているが、それぞれ異なる特徴と役割を持っている。葉緑体は、葉緑体細胞内に存在する小器官で、光合成が行われる場所であり、その葉緑体は二重膜で囲まれており、その内部にはチラコイド膜と呼ばれる構造があり、葉緑素(クロロフィル)で光合成が行われる。
葉緑素は、具体的には、チラコイド膜に埋め込まれた光化学系の中に含まれる。光化学系は、光エネルギーを吸収して電子を励起させる役割を持つ酵素複合
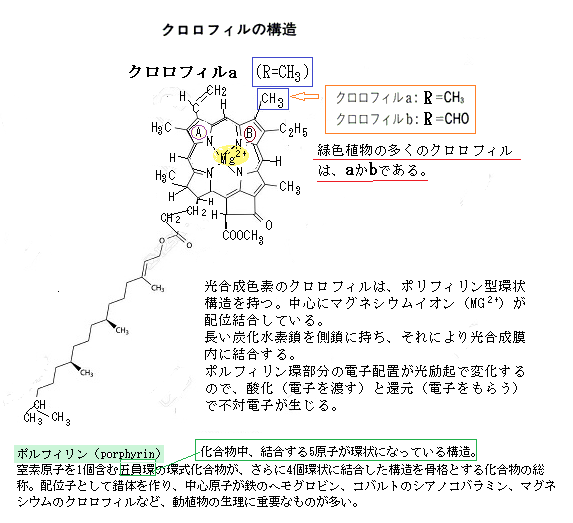 体で、クロロフィルはその中心的な成分になる。チラコイド膜は、光合成の光化学反応が行われる場所であり、その葉緑体内に存在する緑色の色素・葉緑素が、光エネルギーを吸収することで電子を励起し、そのエネルギーを光化学反応に導く働きをする。
体で、クロロフィルはその中心的な成分になる。チラコイド膜は、光合成の光化学反応が行われる場所であり、その葉緑体内に存在する緑色の色素・葉緑素が、光エネルギーを吸収することで電子を励起し、そのエネルギーを光化学反応に導く働きをする。
要約すれば、葉緑体細胞は光合成を行うための葉緑体を含む細胞全体を指し、葉肉細胞はその中でも特に光合成に特化した細胞で、葉の内部で効率的に光を吸収する。葉緑体はその細胞内で光合成を行う小器官として働く。
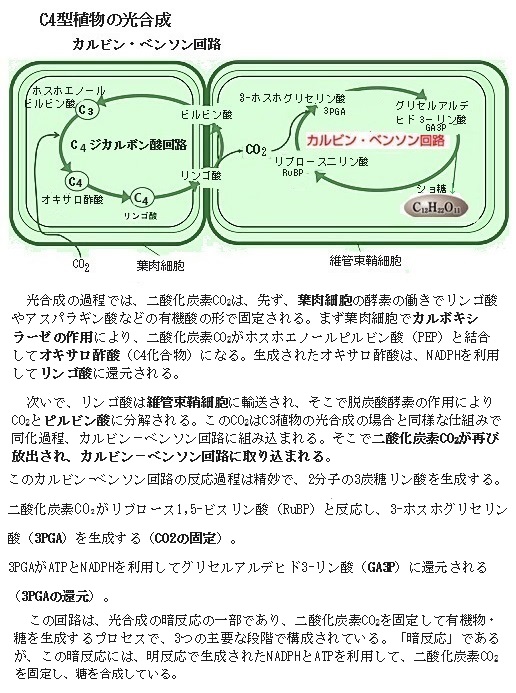
二酸化炭素の固定では、C4植物は、まず葉肉細胞で二酸化炭素CO2をホスホエノールピルビン酸PEPと結合させ、オキサロ酢酸C4H4O5(C4化合物)を生成する。この反応は、ホスホエノールピルビン酸カルボキシラーゼPEPCという酵素によって触媒される。特にC4光合成を行う植物では重要で、このC4化合物オキサロ酢酸は維管束鞘細胞に移動し、そこでCO2を再び放出し、カルビン回路に取り込まれる。このプロセスにより、C4植物は葉肉細胞内のCO2濃度を高め、葉緑体のストロラに局在する光合成酵素であるルビスコの効率を向上させることができる。その結果、C4植物はC3植物に比べて高い光合成効率を持ち、乾燥や高温の環境でも優れた生育を示す。C4植物は、効率的にCO2を固定し、高温や強光下でも高い光合成能力を維持することができるようになる。
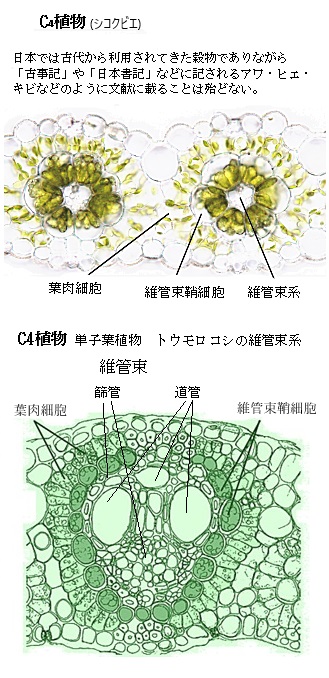 生成されたオキサロ酢酸は、リンゴ酸C4H6O5やアスパラギン酸に変換され、維管束鞘細胞に運ばれる(C4回路)。維管束鞘細胞で、これらの化合物は再びCO2を放出し、カルビン・ベンソン回路に取り込む。維管束鞘細胞で放出されたCO2は、カルビン・ベンソン回路により炭水化物に変換される。その特徴は、C4植物はCO2を効率的に濃縮するため、光合成速度が高く、光呼吸が抑制される。
生成されたオキサロ酢酸は、リンゴ酸C4H6O5やアスパラギン酸に変換され、維管束鞘細胞に運ばれる(C4回路)。維管束鞘細胞で、これらの化合物は再びCO2を放出し、カルビン・ベンソン回路に取り込む。維管束鞘細胞で放出されたCO2は、カルビン・ベンソン回路により炭水化物に変換される。その特徴は、C4植物はCO2を効率的に濃縮するため、光合成速度が高く、光呼吸が抑制される。その利点は高温や乾燥条件下でも効率的に光合成を行うことができる環境適応により、トウモロコシやサトウキビなど多くの作物の栽培地を拡大させてきた。そのC4植物の光合成は、特に高温や乾燥した環境での生育に適しており、これがC3植物との大きな違いである。この仕組みにより、C4植物は水分利用効率が高く、乾燥した環境でも高い光合成速度を維持することができる。生育する過酷な環境に適応するための進化の成果と言える。
C3植物では、光合成を行う緑色細胞は基本的には1種類であるが、C4植物の光合成経路の細部は種によって異なる。C4植物では、光合成組織が維管束のすぐ外側を取り囲む維管束鞘細胞とその外側の葉肉細胞に分化しているが互いに密接に協働している。このC4植物の特徴的なクランツ型葉構造は、主にC4植物に見られる特徴的な葉の構造で、維管束鞘細胞が維管束を取り囲み、その外側を葉肉細胞が放射状に取り囲む。特に「クランツ型葉構造(ドイツ語の「Kranz【花冠、リース】」)」と呼ばれる。このクランツ型葉構造(クランツがたようこうぞう)が、C4光合成において重要な役割を果たす。
C4植物は、CO2を効率的に固定するために、その特殊な葉構造を進化させ、維管束鞘細胞にも葉緑体を多く含ませ、ここでもCO2が濃縮され、カルビン・ベンソン回路で再固定される。
「クランツ型葉構造」での維管束鞘細胞は、植物の水や栄養を運ぶ組織の維管束を取り囲む細胞であるが、これらの細胞は、光合成の一部を行うためにも特化している。しかも維管束鞘細胞の外側に存在する葉肉細胞は、光合成の初期段階を担当することになる。
この配置により、C4植物は効率的に二酸化炭素CO2を固定し、高い光合成効率を維持する。C4植物では、まず葉肉細胞で二酸化炭素がC4化合物(例えばオキサロ酢酸)に固定される。葉肉細胞で固定された二酸化炭素は、C4化合物として維管束鞘細胞に運ばれ、そこで再び二酸化炭素が放出され、この過程により維管束鞘細胞内の二酸化炭素濃度が高く保たれカルビン回路に取り込まれる。維管束鞘細胞内で高濃度の二酸化炭素が一旦放出されることで、光呼吸(酸素がカルビン回路に干渉する現象)が抑制され、カルビン回路の光合成の効率がより向上する。
具体的には、C4植物ではまず葉肉細胞で二酸化炭素がC4化合物(例えばリンゴ酸)として固定される。このC4化合物は維管束鞘細胞に輸送され、そこで二酸化炭素と初に生成されるC3化合物(3-ホスホグリセリン酸PGA))に分解される。このようにして、維管束鞘細胞内で高濃度の二酸化炭素が維持されることで、光呼吸を抑制し、光合成の効率を高めることができる。この過程で放出された二酸化炭素は、維管束鞘細胞内でカルビン・ベンソン回路に取り込まれ、光合成産物に変換される。
光呼吸は、植物が光の下で酸素を消費し、二酸化炭素を放出する過程であり、これは光合成の効率を低下させる要因となっている。C4植物、例えば、トウモロコシやサトウキビは、光呼吸をほとんど行わないため、光合成効率が高い。これらの植物は、CO2を効率的に固定し、光呼吸を抑制する「カルビン・ベンソン回路」と呼ばれる特別な経路を持っている。C4植物は、CO2を高濃度で維管束鞘細胞に供給するため、光呼吸を抑制する。これは、RuBisCO酵素が酸素ではなくCO2と反応する確率を高めるためである。
 |
| 明反応と暗反応は、光合成の過程における2つの主要な段階を示す。 明反応では、光エネルギーを利用してATPとNADPHを生成する。どちらもエネルギーの運搬や変換に重要な役割を果たす分子である。この反応は葉緑体のチラコイド膜で行われる。
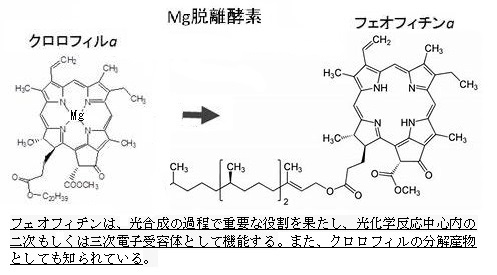
水が分解されて酸素が放出される。 光合成の明反応、つまり光エネルギーを利用してATPとNADPHを生成する反応は葉緑体のチラコイド膜で行われる。その光合成の過程で重要な役割を果たす光化学系I(PSI)と光化学系II(PSII)の2つの異なる光化学系で光エネルギーが吸収される。 光化学系Ⅰ(PSⅠ)と光化学系Ⅱ(PSⅡ)の活性については、光化学系(PSⅠ)は、主に700nmの光を吸収し、NADP+を還元してNADPHを生成する役割がある。 一般的には光化学系Ⅱの方が強い酸化力を持ち、水を分解する能力が高いとされている。光化学系II(PSII)の反応中心クロロフィル(P680)の「P680」の名前は、その吸収極大波長が680 nmであることに由来する。また、P680は非常に高い酸化還元電位(約1.1 V)を持ち、水分解反応を可能にするためのエネルギーを提供する。主に680nmの光を吸収すると、そのエネルギーによってP680の電子が励起され、隣接するフェオフィチンPheと言う分子に電子が移動する。 このフェオフィチンは、クロロフィル分子からマグネシウムイオンMg2+がとれて水素原子2つと置き換わったものの総称である。 クロロフィルが酸性下に置かれるなどしてマグネシウムが脱落すると、代わりに水素原子が結合する事が知られており、これをフェオフィチン化と呼ぶ。 この電子はさらにプラストキノンを経由して電子伝達鎖を通じて移動し、最終的にはNADP+(ニコチンアミドアデニンジヌクレオチドリン酸)を還元して(電子を伝達して)NADPHを生成する。 プラストキノンPlastoquinone(PQ)は、光合成の光化学反応において重要な役割を果たす分子で、具体的には、チラコイド膜を通過する可動性電子伝達体として機能する。 光化学系IIからの2つの電子と葉緑体のストロマから2つのプロトンH⁺を受け取ると、プラストキノンは還元されてプラストキノールPQH₂を形成する。このプラストキノールは、シトクロムb6f複合体を介して電子をプラストシアニンに移動させる役割を担う。 このプロセスにより、チラコイド膜を挟んだプロトン勾配が形成され、最終的にATP合成酵素がADPと無機リン酸Pi(オルトリン酸H₃PO₄を指す)からATPを生成する。 光化学系II(PSII)の反応中心クロロフィルP680が電子を失うと、非常に酸化されやすくなり、水分子が酸化されて酸素分子O2とプロトンH⁺が生成される。つまり、PSIIの反応中心クロロフィルが光を吸収し、水が分解され、酸素とプロトンH⁺、および電子が生成される。この過程で酸素が放出される。 ATPの生成は、電子の移動に伴ってプロトンH⁺がチラコイド内腔に蓄積され、プロトン勾配が形成される。 このプロトン勾配を利用してATPシンターゼがATPを合成する。 NADPHの生成は、PSIで再び光エネルギーが吸収され、電子がフェレドキシンを経由してNADP⁺に渡され、NADPHが生成される。 光合成の明反応では光エネルギーを化学エネルギーに変換し、ATPとNADPHを生成する。これらのエネルギー分子は、次の暗反応で二酸化炭素を固定して糖を合成するために使われる。 暗反応は、カルビン・ベンソン回路とも呼ばれる。明反応で生成されたATPとNADPHを利用して二酸化炭素をショ糖に変換する。 この反応は葉緑体のストロマで行われる。 光が直接必要ないため「暗反応」と呼ばれる。実際には光がないと進行しない。 ATP(アデノシン三リン酸)とNADPH(ニコチンアミドアデニンジヌクレオチドリン酸)は、どちらもエネルギーの運搬や変換に重要な役割を果たす分子で、 ATP 役割は細胞内のエネルギー通貨として機能し、エネルギーを必要とする様々な生化学反応にエネルギーを供給する。 主にミトコンドリアでの呼吸過程や葉緑体での光合成の明反応で生成される。主に、筋肉の収縮・タンパク質の合成や細胞分裂など、多くの細胞活動に利用される。 NADPHは還元力を提供し、脂肪酸やコレステロールの合成、グルタチオンの生成などの生合成反応に関与する。その生成は、光合成の明反応やペントースリン酸経路で生成される。 光合成の暗反応(カルビン・ベンソン回路)で二酸化炭素を固定し、糖を生成する際に利用される。 |
 |
これらの生態系は、二酸化炭素CO2を吸収し、炭素を貯留する能力があり、地球温暖化対策や生物多様性の保全に重要な役割を果たしている。海草や海藻は光合成を通じてCO2を吸収し、その炭素を海底に蓄積している。
カルビン・ベンソン回路は、光合成の暗反応の一部で、二酸化炭素を有機物(ショ糖)に変換するプロセスは、葉緑体のストロマで行なわれる。このプロセスで、酸素がカルビン回路に干渉する現象を「光呼吸」と呼び、この光呼吸では、リブロースビスリン酸カルボキシラーゼ/オキシゲナーゼRuBisCO(ルビスコ)が、酵素として働き二酸化炭素の代わりに酸素を固定してしまうことがある。
現在の地球大気中の酸素濃度は約21%、これを大幅に超える高酸素環境では、植物の生育や光合成に影響を与えることが知られている。この高酸素濃度下では、光合成速度が低下することが多い。これは、光合成の初期段階で働く、葉緑体のストロマに局在する酵素ルビスコRubiscoが酸素に対しても反応するためである。この現象が「光呼吸」と呼ばれる。結果として光合成の効率が低下する。
その一方で、水耕栽培などで水中に酸素を供給することは、植物の生産量を高める。植物の根が十分な酸素を吸収できれば、根の成長が促進され、全体的な生育が向上する。これは、根の呼吸が促進され、栄養吸収が効率化するためである。例えば、トマトの実験では、根付近の酸素濃度が低いと根の伸長量や重量が抑制されることが確認されている。
植物の環境ストレス応答に関して、酸素濃度の変化は重要な要素となる。その一方では、高酸素環境は、植物にとって一種のストレスとなり、その環境に適応するためにさまざまな生理的変化を引き起こす。
その「光呼吸」は、特に高温や乾燥条件下で顕著になるため、時には、植物の成長や生産性に悪影響を与える。光呼吸は、植物が光の下で酸素を消費し、二酸化炭素を生成する過程である。特に高温や乾燥条件下では、光呼吸の速度が光合成の速度を上回ることがあり、これが植物の成長や生産性に悪影響を与えることがある。つまり、光呼吸が顕著になる理由の一つは、光合成酵素であるRubisCOが酸素と二酸化炭素の両方と反応するためで、高温条件下であれば、RubisCOと酸素との反応が促進され、光呼吸が増加する。
RuBisCOの酸素添加反応によって生じた2分子の2-ホスホグリコール酸は、その後、ペルオキシソームとミトコンドリアにおいて代謝されて、最終的に、1分子のCO2と1分子の3-ホスホグリセリン酸に変換される。そして、再びカルビン・ベンソン回路の基質として用いられる。この経路では、光エネルギーを利用する明反応で生じたATPやNADPHのエネルギーを用いることで、酸素O2を消費して二酸化炭素CO2を生成することから「光呼吸」とも呼ばれている。
C4植物の葉肉細胞と維管束鞘細胞の間での二酸化炭素のやり取りは、光合成の効率を最大化するための重要なプロセスと言える。
リブロース-1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼRuBisCOは、光合成のカルビン-ベンソン回路において二酸化炭素を固定する酵素で、この酵素は、リブロース-1,5-ビスリン酸RuBPに二酸化炭素を結合させて、2分子の3-ホスホグリセリン酸を生成する。
リビスコRuBisCOは、植物の葉緑体に大量に含まれ、地球上で最も多いタンパク質ともいわれる。また、この酵素はカルボキシラーゼ反応だけでなく、「オキシゲナーゼ反応」も行う。この光呼吸では、RuBisCOが酸素を基質として使用し、2-ホスホグリコール酸を生成する。この過程で、同化されたCO2が再放出されるため、光呼吸と呼ばれれる。この光呼吸は、特に高温やCO2濃度が低い条件下で顕著に見られるから、植物の成長効率に悪影響を与えることになる。
C4植物では大気中のCO2が 気孔から取り込まれると、まず葉肉細胞でホスホエノールピルビン酸phosphoenolpyruvate(C3H5O6P)カルボキシラーゼphosphoenolpyruvate carboxylase (PEPC)の作用によりホスホエノールピルビン酸phosphoenolpyruvate (PEP) に固定されてオキサロ酢酸となり、直ちにリンゴ酸C4H6O5やアスパラギン酸などのC4化合物 となる。ただ、どのようなC4化合物となるかは種によって異なる。
ホスホエノールピルビン(PEP)は、生化学的に非常に重要な有機化合物で、PEPは解糖系や糖新生の経路で重要な役割を果たす。主な特徴は、PEPは生体内で最もエネルギーの高いリン酸結合を持つ(高エネルギーリン酸結合)。また解糖系では、PEPはピルビン酸キナーゼによってピルビン酸に変換され、この過程でATPが生成される。
糖新生では、PEPはオキサロ酢酸から生成され、グルコースの合成に利用される。これらのC4化合物は維管束鞘細胞に輸送され、そこで脱炭酸酵素作用によってCO2と、ピルビン酸C3H4O3やPEPなどのC3化合物に切り離される。解糖系では、1分子のグルコースが2分子のピルビン酸に分解される過程で、3-ホスホグリセリン酸(3-PGA)が生成されが、カルビン回路では、リブロース-1,5-ビスリン酸(RuBP)に二酸化炭素CO2が結合して2分子の3-PGAが生成される。
ここではCO2は、リブ ロース二リン酸(RuBP)カルボキシラ―ゼの働きにより、RuBPに固 定され2分子のホスホグリセリン酸(PGA)となり、そのPGAがカルビン回路でデンプンなどの炭水化物が合成される。残りのC3化合物は葉肉細胞へ戻り、 ピルビン酸リン酸ジキナーゼ(PPDK)の働きによってホスホエノールピルビン酸PEPとなり、再度、CO2固定の受容体になる。
C4植物の名前の由来は、CO2固定の初期産物がC4化合物のオキサロ酢酸であることによるが、一方 、C3植 物では、大気中のCO2はリブロース二リン酸RuBPカルボキシラーゼによって直接カルビン回路に取り込まれる。C3では2種類の光合成細胞 を必要としないので、C3植物の名前は、CO2固定の初期産物がC3化合物のホスホグリセリン酸PGAであることに由来する。
このC4化合物は維管束鞘細胞に移動し、そこでCO2が再び放出し、カルビン回路に取り込まれる。このプロセスにより、この高濃度のCO2は、カルビン回路で効率的に利用されるため、光合成の効率が向上する。C4植物の維管束鞘細胞のCO2濃度を高め、葉緑体のストロマに局在する光合成酵素であるルビスコの効率を向上させることができる。その結果、C4植物はC3植物に比べて高い光合成効率を持ち、乾燥や高温の環境でも優れた生育を示す。
目次へ
C3植物は、光合成の過程で最初に生成される安定な化合物が3-ホスホグリセリン酸(3-PGA)であることからその名が付けられている。これに対し、光合成のCO2初期固定をC4ジカルボン酸回路(C4回路)により行う植物をC4植物、ベンケイソウ型有機酸代謝を行う植物をCAM植物と呼ぶ。
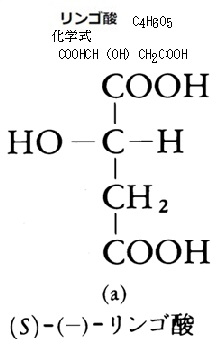 |
| リンゴ酸malic acidは、ヒドロキシ酸に分類される有機化合物の一種、リンゴやブドウ、バナナ・トマトなどの果物に多く含まれ、爽やかな酸味が特徴。リンゴ酸の和名は、リンゴから見つかったことに由来する。天然に存在するのは(S)-(-)-リンゴ酸である。 動物に比べて、植物の果実はクエン酸やリンゴ酸などの有機酸を多量に蓄積している。光合成植物は太陽エネルギーを取り込んで二酸化炭素から糖を合成する。糖や脂肪は炭素・水素・酸素といった元素によりエネルギーを保有する。 生物は、これらのエネルギーを生存、成長のために利用できるように取り出す生化学反応系をもっているが、同時に炭素・水素・酸素からなる中間産物を自身の体を作る材料として利用している。 つまり糖、脂肪の酸化によるいろいろな物質変化の流れと、それに付随するエネルギーの流れがある。それにより炭素は二酸化炭素に、水素は水になり、そのエネルギーはATPと呼ばれる生体エネルギーの通貨のような物質に移される。 具体的には、クエン酸は柑橘類、例えばオレンジやレモンなどに多く含まれ、リンゴ酸はリンゴやモモなどに多く含まれている。これらの酸は果実の成熟過程で蓄積され、熟すとともに分解されていく。 また、クエン酸やリンゴ酸は、動物のエネルギー代謝にも重要な役割を果たしている。これらの酸は、動物が食物からエネルギーを取り出すための代謝経路、クエン酸回路において重要な中間体として機能する。しかも、果実の酸味は動物にとって魅力的であれば、植物が進化の過程で種子散布を効果的に行うための戦略の一つとなる。 |
葉肉細胞でCO2がホスホエノールピルビン酸カルボキシラーゼPEPCによってオキサロ酢酸OAAに固定され、オキサロ酢酸はNADPHによりリンゴ酸に還元される。
リンゴ酸は、葉肉細胞から維管束鞘細胞に輸送され、そこで再びCO2を放出する。このCO2はカルビン・ベンソン回路に取り込まれ、最終的に糖などの有機物に変換される。
C4光合成はC3光合成よりもエネルギー効率が高く、乾燥や高温環境に適している。)
この3-PGAは炭素が3つ含まれているため、「3炭素化合物」と呼ばれる。C3植物の光合成は、カルビン・ベンソン回路を利用して行われる。この回路では、二酸化炭素CO2がリブロース-1,5-ビスリン酸(RuBP)と結合し、3-PGAが生成される。このプロセスがC3植物の特徴である。C3植物の光合成の場所は葉肉細胞で、通常の環境下では効率的に光合成を行うが、乾燥や高温の環境ではC4植物やCAM植物に比べて効率が低下しやすい。
CAMはCrassulacean Acid Metabolism (ベンケイソウ型有機酸代謝)の略で,CAM型光合成を行う植物をCAM植物と呼ぶ、ベンケイソウ型有機酸代謝Crassulacean Acid Metabolism(CAM)は、多肉植物や乾燥地帯に生息するサボテンなどの植物が活用する光合成の一形態である。
CAM植物は、夜間に気孔を開いて二酸化炭素CO₂を取り込みリンゴ酸として液胞に蓄える。昼間に気孔を閉じて水分の蒸散を防ぎながら、太陽光エネルギーに依存しながら夜間に取り込んだCO2を効率的に再固定し、その一方では、酸素を取り込む光呼吸を抑制することで高い光合成能を達成する。C4植物は、葉肉細胞と維管束鞘細胞の2種類の細胞を使ってCO2を濃縮し、効率的に炭素同化を行う。
その主な特徴を要約すると、そのCO2の前固定では、葉肉細胞でCO2がホスホエノールピルビン酸カルボキシラーゼ(PEPC)によってオキサロ酢酸(OAA)に固定する。その後、維管束鞘細胞でOAAが脱炭酸され、CO2が再固定される。このC4光合成はC3光合成よりもエネルギー効率が高く、乾燥や高温環境に適している。C4植物には、トウモロコシやサトウキビなどが含まれるが、特に乾燥地帯や高温環境で優れた適応能力を発揮している。
この代謝経路では、夜間に気孔を開いてCO₂を取り込み、ホスホエノールピルビン酸カルボキシラーゼ(PEP-C)によってリンゴ酸にして液胞に蓄える。昼間には、気孔が開いたままリンゴ酸の脱炭酸反応を進め、その際CO₂を再放出する。
このベンケイソウ型有機酸代謝CAMはベンケイソウ科植物に限らず多くの植物にみられる代謝であるが、19世紀にベンケイソウ科植物の葉の有機酸の含量が夜間に増し昼間に消失する現象が、この科の植物に特有な代謝として認識され研究されたことに由来する。
CAM植物はシダ植物3科5属、裸子植物1科1属、単子葉植物8科98属、双子葉植物33科239属に分布し、ラン科での推定種数7,000種を含めて約16,000種にみられる。この数は地球上の維管束植物の約6%に相当する。このように多数の植物にみられるCAMの進化は、環境条件によって誘発された適応進化によるものと考えられている。
ベンケイソウ型有機酸代謝(CAM)を持つ植物は、高温・半乾燥地帯に適応し、熱帯林や亜熱帯林の着生植物、高山植物や塩生植物(海浜や岩塩地域などの塩分に富む土壌に生育する植物、アッケシソウ・ハママツナなど一般に多肉のものが多い)、あるいは水生植物で確認されいる。日本においても、岩場・海岸・山草地・畑・路傍・極相林(日本では、ブナやミズナラなどの陰樹が多く見らる)・貧栄養湖(栄養塩が乏しく生産活動の低い湖沼、摩周湖・十和田湖など。)などにCAM植物は生育している。
栽培種を含め国内に分布するCAM植物は、ヒカゲノカズラ植物門ミズニラ科の5種、シダ植物門ウラボシ科3種、イノモトソウ科1種、マツ門(裸子植物門)ウェルウィッチア科1種、被子植物ではモクレン類コショウ科1種、単子葉類7科25属86種、真正双子葉類11科53属140種で、合計23科83属237種あり、そのうち栽培種は185種と全体の約8割を占め、在来種も56種の分布が確認されている。そのほとんどの種は、岩場や海岸など限界に近い水分環境下で生育している。帰化種も33種確認さている。
CAM植物はCAM型光合成という生理機能と多汁性という形態学的特性を合わせ持つ。砂漠や着生環境などの水分ストレスが多発する環境に適応した植物に多くみられる。着生植物は、土壌に根を下ろさず、他の木や岩などに根を張って生活する植物、一般的には、樹木に付着して生活する植物を指すが、岩に着生するものも含まれる。塩生植物のアッケシソウ(厚岸草or珊瑚草)は、塩分を含む汽水に生育する1年草で、北海道のサロマ湖や能取湖、瀬戸内海沿岸などに分布している。アッケシソウは中でも特に強い耐塩性を示し、むしろ塩の存在に依存する植物である。生育過程が進むにつれて、塩を蓄積することにより耐塩性を獲得する強塩生植物である。過剰な塩類の液胞内への蓄積と連動して、浸透圧を調整する強い耐塩性を示し、細胞内にグリシンというアミノ酸を基にした化合物を蓄積し、浸透圧を調整する。また、秋になると茎や枝が紅紫色に紅葉するのは、ベタシアニンという色素が関与している。
(グリシンは、植物や動物、微生物など多くの生物に広く存在している。浸透圧調整や植物が高温や乾燥、塩分ストレスなどの環境ストレスに耐えるのを助ける。)
ハママツナは、海水が影響する砂地に生育する一年草、春に発芽して秋に開花結実する。高さ15-35cmの茎を持ち、葉は肉質で線形、葉を噛むと塩辛い味がする。CAM植物として、露点が低い夜は、気温が下がり空気中の水蒸気が少なく乾燥しているが、気孔を開いて空気中のCO2を取り入れて、それを水和させ炭酸水素イオンHCO3-にし、これを葉肉細胞の酵素の働きホスホエノールピルビン酸(PEP)とPEPカルボキシラーゼを用いてオキサロ酢酸(C4)にする。そしてNADPHとリンゴ酸デヒドロゲナーゼを用いてリンゴ酸(C4)にする。そして、このリンゴ酸は昼まで有機酸として液胞中に蓄える。昼は気孔を閉じて水の蒸散を防ぎながら、有機酸を分解してCO2を生成し、これを通常の光合成により、糖などの有機物の原料とすることで成長する。
アフリカ西部にもわずかに分布するが、ほとんどがアメリカ大陸・西インド諸島の熱帯・亜熱帯の原産パイナップル科(アナナス科)は、砂漠から森林まで分布する。パイナップルゲノムは、全ゲノムの重複事象以前から存在する7本の染色体に基づ核型があり、このパイナップル系統によりC3型光合成からCAMに移行したことが示される。そのCAM関連遺伝子が光合成組織で日周的な発現パターンを示している。CAM経路の遺伝子には、概日周期の時計遺伝子による制御を受けるシス調節配列が豊富に存在する。これはCAMと概日時計制御を結びつけるシス調節の最初の報告であった。パイナップルのCAM光合成は、C3植物が持つ経路を再構成することで進化した。つまり既存の遺伝子が調節性の新機能を獲得したことによって進化したものであり、全ゲノム重複や縦列遺伝子重複によって獲得した新機能によるものではない。
(全ゲノム重複は、染色体全体が重複する現象で、これにより、すべての遺伝子が二重に存在するようになる。例えば、酵母のゲノムは約1億年前に全ゲノム重複を経験したと見られている。このような重複は、新しい遺伝子機能の進化を促進する可能性がある。
縦列遺伝子重複は、特定の遺伝子や遺伝子群が連続して重複する現象で、通常、同じ染色体上で隣接して重複する現象で、これにより、同じ遺伝子が連続して並ぶことになる。この重複は、遺伝子の機能や発現パターンに変化をもたらすことがある。 これらの重複現象は、進化の過程で新しい遺伝子機能を生み出す重要なメカニズムとされている。
人類のDNAは30億の塩基対からなるが、遺伝子を含むDNAのある領域が重複して存在するという現象をDNA上の諸々で見られる。これを遺伝子重複と言う。多くの場合、まったく同じというわけではなく、異なっている部分がある。明らかに、元々はまったく同じで あったものが長い年月で変異が加わり、異なってきたと推定できる。)
ラン科は単子葉植物の中で最も種の数が多く、種はなお分化を続けているため「種の爆発の時」といわれている。ランは樹木の樹皮や岩の上などに根を張る着生種と、林の中や草原などの地面に生育する地生種に分けられる。ランは分化を続けており、その分布は植生の限界にまで広がり地生ランは世界のほぼ全域にみられる。一方、着生ランの分布域は赤道を中心に南緯と北緯それぞれ30~40度が限界になっている。
パイナップル科やラン科植物など熱帯気候雨林に生息するCAM植物の場合は、着生環境で水が得にくい環境に生息するが、夜間にCO2を吸収することは、C3植物との競合しないためCO2を吸収しやすいと言っ利点もある。CAM型光合成は砂漠などの水分が慢性的に不足しており、かつ昼夜の温度差が大きい環境に適応したものだと考えられる。通常の植物は昼に気孔を開け、CO2を取り込む。ただし、このような環境下の場合、同時に大量の水分を失ってしまう。しかし、CAM植物は涼しい夜に気孔を開け、CO2の取り込みを行い、昼は気孔を閉じることで水分の損失を最小限に抑えることができる。
このようにCAM植物は、ベンケイソウ科Crassuraceaの一部植物のように、光合成で特殊な有機酸代謝Acid Metabolism行うことで、砂漠や乾燥地帯などに自生するサボテンやベンケイソウなどの典型的なCAM植物や、他の植物体に着生するものが代表的で、水分の供給が不十分なところに生育する植物に多く見られる。昼は気孔を閉じて水の蒸散を防ぎ、気温が下がり露点が低い夜に気孔を開いて空気中のCO2を取り入れて、これをリンゴ酸やアスパラギン酸などの有機酸として液胞中に蓄える。
昼は有機酸を分解してCO2を生成し、これを通常の光合成により、糖などの有機物の原料とすることで成長する。つまり、貯蔵されたリンゴ酸がNADP+と反応し、NADPHとピルビン酸(C3)とCO2が生成される。このように濃縮されたCO2はカルビン - ベンソン回路に入り、ピルビン酸はデンプンとなり、ホスホエノールピルビン酸(PEP)の原料となる。PEPはピルビン酸からATPとピルビン酸オルトリン酸ジキナーゼを用いて合成される。
C3植物が優位にあるのは、光合成の過程で二酸化炭素を直接カルビン回路に取り込むことにある。C3植物の光合成経路が、そのカルビン - ベンソン回路のみの場合と比べて、CAM経路はエネルギーを余計に消費する。通常の光合成と比較すると、CO2同化に加えて有機酸の蓄積と分解に余分なエネルギーを必要とするため、エネルギー変換の効率は低くなるが、水の損失を防ぐことができるというのが最大の効用がある。CAM型光合成生物は、系統的にも多様で、同じ科に属するものでもC3型とCAM型が混在している例も沢山ある。また、水の供給が不十分な時はCAM型光合成を行い、十分あるときは通常の光合成植物(C3型)と同様に、昼間に気孔開いて光合成をおこなう種もある。つまりCAM型植物のエネルギー変換効率は悪いが、水が得にくい環境に何とか適応しようとして発達した光合成方式と言える。
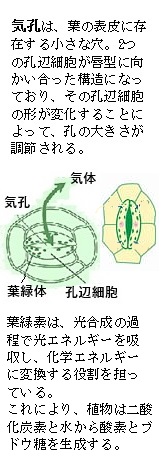 気孔の開閉は、通常の植物では、主として青色光を吸収するホトトロピンという色素タンパク質の光応答によって制御されており、また、乾燥した条件では、植物ホルモンのアブシジン酸が合成されて気孔の開閉を妨げる。
気孔の開閉は、通常の植物では、主として青色光を吸収するホトトロピンという色素タンパク質の光応答によって制御されており、また、乾燥した条件では、植物ホルモンのアブシジン酸が合成されて気孔の開閉を妨げる。ホトトロピンは、植物の気孔の開閉において重要な役割を果たす青色光受容体で、青色光を受け取ると、細胞膜のプロトンポンプ(H+-ATPase)が活性化される。このATPのエネルギーを利用して水素イオンH+を輸送する細胞膜ポンプが活性化され、水素イオンH+を細胞外へ能動輸送する事により膜電位が過分極し、ついで、同じく細胞膜にあるカリウムチャンネル がその過分極に応答して開き、孔辺細胞内にカリウムイオンK+が取り込まれると考えられている。
(カリウムの単体金属は激しい反応性を持つため、電子を1個失って陽イオンK+になりやすく、自然界ではその形でのみ存在する。カリウムイオンK+は気孔の開閉を制御し、二酸化炭素の取り込みや酸素の放出、水分の蒸散を調整する。また.カリウムイオンK+は光合成に関与する酵素を活性化し、光合成の効率を高める。
地殻中では2.6 %を占める7番目に存在量の多い元素であり、花崗岩やカーナライトなどの鉱石に含まれる。)
細胞は一般に内部が負、外部が正に分極しており、その分極が定常状態より大きくなることを過分極、小さくなることを脱分極と言う。つまり、細胞内の水素イオンH+を細胞外に輸送し、膜電位を過分極させる。これにより、電位依存性のカリウムチャネルが開き、カリウムイオンK+が孔辺細胞内に取り込まれる。結果として、孔辺細胞の体積が増加し、気孔が開く。気孔が開いた状態の孔辺細胞には、閉じた状態の数倍濃度のカリウムイオンK+が蓄積しており、カリウムイオンK+の蓄積により浸透圧が上昇し、水が取り込まれ、孔辺細胞の体積が増加する。孔辺細胞は内側に厚い細胞壁、外側に薄い細胞壁を持っており、孔辺細胞の体積が増加すると外側の薄い細胞壁が押され、孔辺細胞間の孔が開くと考えられている。
一方、乾燥条件下では、植物内で合成された植物ホルモン・アブシジン酸が応答して気孔の閉鎖が促進させる。植物ホルモン・アブシジン酸は気孔の閉鎖・種子や芽の休眠・成長抑制を行う。また凍結耐性や乾燥耐性などの多くのストレスに対しても抵抗する。
(青色光は植物において、胚軸の伸長抑制・光屈性・葉緑体定位運動・気孔開口などの様々な反応を引き起こす。これを「青色光効果」と呼ぶ。胚軸の伸長抑制では青色光受容体タンパク質クリプトクロムが、光屈性、葉緑体定位運動。気孔開口では、植物の青色光受容体タンパク質の一種フォトトロピンが青色光受容体として機能している。)
 |
| 「チャツボミゴケ」は、硫黄泉などの酸性泉に限って生育する特殊な苔で、鉄を溶かし込む鉄泉が流れ込む川原の岩にびっしり付着し、川の流れの中に、ビロードの「チャツボミゴケ」を敷き詰める独特の景観をつくりあげるのです。 逆川橋のもう一つの景観は、樹皮に付着して懸垂する糸状の地衣類「サルオガセ」です。地表、岩の上、樹皮上などに着生し、霧のかかるような所では種類が多いようです。他の植物が生育できないような厳しい環境に進出できるのも地衣類の特徴で、都会でもコンクリートの表面に懸垂するといいます。 苔に近い仲間の地衣類に属し、長く木の枝から垂れ下がり、空気中の水蒸気を吸って自立して育ち、決して寄生植物ではありません。 霧藻(きりも)とも呼ばれ、水分と光合成だけで成長し、栄養を他から奪い取ることはないそうです。 |
ティランジア属エアプランツは、パイナップル科チランジア属の一部の植物の総称で、主に北アメリカ南部からメキシコの乾燥地帯に自生しており、そのほとんどが樹木や岩場に着生している。大気中の水分を吸収して生きる植物で、樹木や岩に着生する。
CAM型光合成を行うことで、乾燥した環境でも生き延びられる。エアプランツ(チランジア)は土を使わずに育てられるので、飾り方の幅も広く、インテリアグリーンとして人気の植物で、エアプランツの葉には、「トリコーム」という繊細な繊毛が全体を覆うことで、大気中のわずかな水分を吸着し、CAM型光合成を行うことで、エアプランツ全体の養分として成長している。
エアプランツの光合成では、直射日光を避け、明るい日陰や半日陰の環境が最適で、または植物育成LEDライトを使用することで、室内でも光合成を促進することができる。
サルオガセ(猿尾枷)は、奥蓼科の渋川上流などの森林で見られる着生植物である。サルオガセは地衣類の一種で、樹皮に付着して糸状に垂れ下がる特徴がある。この植物は、藻類と菌類が共生しているため、サルオガセ科の地衣類である。我が国では亜高山帯の針葉樹林に多く見られる。
地衣類は、菌類の仲間で、必ず藻類と共生している。菌類は、藻類と共生することで 「地衣体」と呼ばれる特殊な生体を作り出す。地衣類を構成している菌と藻は、互助関係にあり、菌類は藻類に安定した住み家と生活に必要な水分を与えるかわりに、藻類が光合成で作った炭水化物を栄養として共生菌へ輸送する.。
両者の共生関係は非常に密接で、地衣体の形態・生理機能・繁殖方法などは単独の生物と同じように遺伝する。共生藻には光合成関連遺伝子が共生関連遺伝子に含まれ、また光合成産物の炭化水素化合物を輸送する共生菌(リビトールトランスポーター遺伝子) も共生関連遺伝子に含まれてる。これらの遺伝子は共生による光合成の活性化と共生藻から共生菌への光合成産物の輸送の働きを担っている。
さらに共生関連遺伝子には、リン酸化合物と窒素化合物の輸送に関わる遺伝子も複数含まれており、これらは共生菌から共生藻へのリン酸化合物と窒素化合物の輸送の働きを担っている。このように栄養授受を通して共生関係は維持されていることから、栄養授受に関与する遺伝子の発現量の増加は、地衣類の共生において最も重要な役割を果たしている。また、光依存的に炭化水素化合物の代謝活動を制御する FTR 遺伝子や二酸化炭素の輸送や固定に関わる炭酸脱水酵素遺伝子が共生関連遺伝子に含まれることから、地衣体内の光条件や二酸化炭素濃度が、共生時の藻類の代謝活動を制御している可能性も示している。つまり、あたかも独立した生物体のようである。
地衣類は世界中に広く分布し、世界で約2万種、日本では約1800種が知られている。実は、身近に普通に生息しているが、大気汚染や環境の変化には抵抗力が弱い種類が多いために、都市部ではあまり多くの種類は見られない。しかし、 都市部から離れた里山や高山では、地表や岩の上、木の幹などに、地味であるが清やか垂れさがる光景は涼やかである。
その一方、アメリカ合衆国アリゾナ州の南西部からメキシコのカリフォルニア湾周辺にかけて広がっているソノラ砂漠や、チリのアンデス山脈と太平洋の間に広がる海岸砂漠であるアタカマ砂漠など、その過酷な環境下でも地衣類が見られる。これらの地衣類は、干ばつの時期には休眠状態になり、雨が降ると再び活動を再開する。砂漠のような厳しい環境でも地衣類はわずかな水分を利用して繁茂している。
地衣類は非常にゆっくりと成長し、1年に1mm以下しか成長しないこともあるが、数千年も生きることができる。そのため、砂漠や北極、南極などの過酷な環境でも長期間にわたって生き続ける。
目次へ