| 目次 |
| 1)地磁気逆転 チバニアン期のシベリア高気圧 千葉セクション |
| 2)チバニアン期の古生物 日本海の形成 チバニアン期以前の古生物 北太平洋亜熱帯循環と代表的な海流および水塊 |
| 3) 第四紀氷河時代 ナウマンゾウの時代 |
下層雲は、その名の通り、上空の最も低い位置、約2,000m付近の青空に浮かぶ白いわたのような「青天積雲」に代表される。雨や雪などの降水をもたらす雲は、中層雲である「乱層雲」を除けば、基本的にこの下層雲が主体となる。その青天積雲が発達し、大きくなると中層や高層にまで広がる「雄大積雲」となり、雲の下では激しい雨に突風が伴う。特に「積乱雲」は発達すると、高さ13,000mまで上昇することもあり、他の雲と比べて非常に分厚く大きな雲となる。積乱雲は壮大で美しいが、ゲリラ豪雨や落雷などの激しい気象現象をもたらす。
 中層雲は、上空の真ん中付近 2,000m~7,000mに発生する雲、この雲は小さな水の粒が集まってできており、青空に白い羊の群れがいるように見える。中層雲は、上層雲の巻雲や下層雲の晴天積雲とは異なり、水平に広がっていることが多い、これは、中層雲が気温の不連続面に沿った広範囲の弱い上昇気流により生じるためと言う。
中層雲は、上空の真ん中付近 2,000m~7,000mに発生する雲、この雲は小さな水の粒が集まってできており、青空に白い羊の群れがいるように見える。中層雲は、上層雲の巻雲や下層雲の晴天積雲とは異なり、水平に広がっていることが多い、これは、中層雲が気温の不連続面に沿った広範囲の弱い上昇気流により生じるためと言う。地磁気geomagnetismは、地球自体が持つ磁性と、その地球によって生じる磁場を言う。地磁気の大部分は、地球内部の外核といわれる部分で発生する。ここでは鉄とニッケルが主成分となっており、巨大な圧力と高温(約4400℃~約6100℃)のバランスの中で溶融状態にあり、地球磁場はこの導電性の高い鉄の流体運動・対流により生じている。
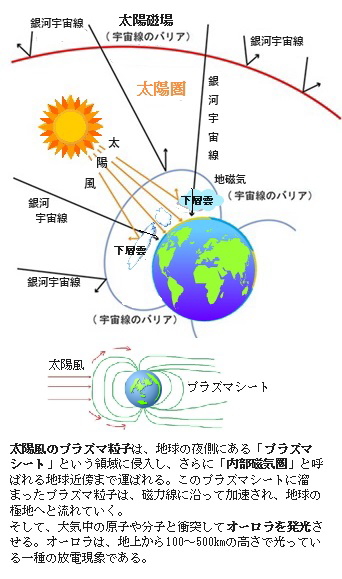
宇宙空間にも広がる地球磁場は、太陽から放出された高エネルギー粒子の流れ、太陽風の圧力を受け、太陽と逆側に圧縮され、また地球の反対側では吹き流されるようになる。地球の磁気圏は、太陽風のため昼側では地球半径の10倍ぐらいまでに圧縮され、夜側では細長い尾を引く。こうした地球磁場が支配できる領域を磁気圏と呼ぶが、地球は磁気圏を持つことで太陽風の高エネルギー粒子に直接さらされずに守られ、生命が宿る美しくも苛烈な惑星となった。
磁場はN極・S極からなる磁極で、正電荷・負電荷からなる電荷とは異なり、それぞれ単独では存在できない。磁極の強さが「磁気量」と呼ばれる。この磁場は、空間それぞれで向きと大きさを持つ物理量で、通常はテスラTという単位で表されるが、地球の磁場はとても弱いため、ナノテスラnTが用いられる。地球は、一つの大きな磁石とみなすことができる。北極は地球の北極点であり、南極は地球の南極点であるが、北極は磁極とは異なり、南極は磁極と一致している。地磁気の観点から言えば、北極は「N極」であり、南極は「S極」、つまり、地磁気の磁場の向きに基づいて命名されている。方位磁針の針は、北を指すN極と南を指すS極を示すが、方位磁針の指す北(磁北)は、地理的な北とは異なる。地理的な方向を知る際の「正確な真北」とは、ある地点を通過する子午線が示す北、つまり北極点の方向を指す。その地図上の真上とは、地球の自転軸の北端(北緯90度地点)であり、コンパスが指す北は「磁北」と呼ばれる。地球の磁力を地磁気と呼ぶ。
太陽の表面は「コロナ」と呼ばれるセ氏 100万度もの極めて高温の大気で覆われている。コロナは、時には爆発的に膨張し、陽子と電子の粒子からなる「プラズマ」が太陽の外へ飛び出し地球まで達する。これが太陽風である。地球に降り注ぐ宇宙線や太陽からの紫外線を和らげる役割も果たす地磁気は、地球の大気や水の宇宙空間への拡散を防ぐ。
地磁気逆転geomagnetic reversalは、地球の地磁気の向きが南北逆になる現象で、この逆転は、地球の磁場が数万〜数十万年ごとに反転することを意味する。最後の磁場の逆転は約78万年前に起こっている。逆転時には地磁気の強度が最大約1/10まで減少した。
地磁気逆転途中の5,000年間、中国の黄土高原では冬の季節風が運ぶ砂塵の粒が大きくなり、堆積速度が3倍となった。冬の風がそれだけ強まっていた。その5,000年間は地磁気強度の減少と、銀河宇宙線の50%以上の増加期間と一致する。冬の季節風の強化は、銀河宇宙線の増加に伴い増えた下層雲の日傘効果で、大陸が太平洋に比べより強く冷え、シベリア高気圧がより強まったことによる。そのシベリア高気圧は、主にシベリア内陸部特有の厳しい寒冷化を産み、大気を冷却し、その密度を凝縮させた。また、銀河宇宙線がその気候変動の大きな要因に加わった。
78万年前の地磁気逆転途中に、銀河宇宙線が増加したことによる下層雲の日傘効果がシベリア高気圧をより強化し、冬の季節風をより激しくした証拠が見つかっている。この現象は、銀河宇宙線が地球の気候変動に影響を及ぼす証拠を示すものとなった。冬季の季節風が強化されると、日本海上で北西季節風が卓越し、日本海側は厳冬となり豪雪になる。季節風は、夏は海洋から大陸へ、冬は大陸から海洋へ吹く。それは大陸と海洋の比熱差によって起る。その冬の海洋への季節風強化と同じ期間に、78万年前の地磁気逆転途中の約5,000年間にわたって黄土粒度が粗くなり、堆積速度が3倍以上増加する「冬の季節風の強化」の痕跡が大阪湾堆積物コアで発見された。この風の強化期間は、地磁気逆転に伴い地磁気強度が1/4以下に減少し、それが銀河宇宙線が50%以上増加した期間と一致する。
これは「スベンスマルク効果」により、銀河宇宙線の増加は、大気中の水蒸気やエアロゾルをイオン化し凝結核(ぎょうけつかく)を増やして下層雲量を増加させ、その雲の日傘効果で大陸がより低く冷却され、シベリア高気圧が強化されたことを示唆していた。
ちりやほこりを含まない清浄な空気中では、相対湿度(その温度における飽和水蒸気量に対するその時の空気中の水蒸気量の比率)が100%を超えても水滴はできない。大気中を浮遊する微粒子(エアロゾル)は、水滴の半径を大きくし、低い過飽和度でも水滴ができるようにする。エアロゾルは、水蒸気を凝結 させるための核の役割を担う。
地磁気逆転途中の現象として、大阪湾堆積物コアで痕跡が見つけられている2−3℃の年平均気温の低下と気温年較差の増大に加え、今回新たに冬の季節風の強化の痕跡が見つかった。これらの痕跡の原因が下層雲の日傘効果によることがほぼ確実となった。
 (海洋観測船や深海掘削船による研究航海の際に掘削・採取される連続する柱状の試料marine core samples【コアcore】は、全地球規模における過去の地球環境変動、例えば海水面変位・地磁気の変移・海流系の変動・水温変化などの記録を留める。これらのコアは、海底から採取され、物質循環プロセスや海洋環境変動の実態を復元するために詳細に解析される。海洋コアは、地球環境システムの理解や近未来の気候変動予測において重要な情報試料にる。
(海洋観測船や深海掘削船による研究航海の際に掘削・採取される連続する柱状の試料marine core samples【コアcore】は、全地球規模における過去の地球環境変動、例えば海水面変位・地磁気の変移・海流系の変動・水温変化などの記録を留める。これらのコアは、海底から採取され、物質循環プロセスや海洋環境変動の実態を復元するために詳細に解析される。海洋コアは、地球環境システムの理解や近未来の気候変動予測において重要な情報試料にる。また、大阪湾地域の更新世チバニアン期(約77.4万年前~12.9万年前)の地磁気逆転当時の海成粘土層の特徴や形成過程についても調査されている。この湾内の特徴は、海成粘土層に粗粒な部分が挟まっていることにある。
この現象は、新生代第四紀後氷期の約5,000年前の縄文海進により、海面が高くなった時期に潮流や沿岸流による運搬作用が強くなり形成された。
後氷期の急激な海進について、大阪湾の状況は非常に興味深いです更新世チバニアン期の約21,000~約19,000年前の最終氷期の最盛期に、北半球や南極大陸に大陸氷床が広く発達し、それにより海面が120~130m低下した。
後氷期海進は、地質学的には完新世海進、または日本では縄文海進と呼ぶ。最終氷期の最寒冷期後(約19,000年前)から始まった温暖化に伴う海水準上昇を指す。日本では縄文時代が始まる約16,000年前に近い時期にこの海進が発生した。
後氷期気温が上昇し始めると、海面も上昇し始めたこの時期、大阪湾では縄文海進による急激な海面上昇が起った。 特に約9,850年前~5,000年前にかけて、海面は現在の高さを超え、最大で約2.2m上昇した。この期間に、古河内湾を誕生させた。
深海底より5倍以上速い堆積速度を持つ大阪湾海底堆積物について国立の産総研が1997年にボーリングした堆積物コアから、100年スケールの高解像度の地磁気逆転データを取得し2006年に発表した。
このコアから78万年前に起こった最後の地磁気逆転において数百年単位の小反転が少なくとも4回起こったことを発見し、インドネシアのジャワ島、中国の黄土高原レスでも同様の小反転が見つかっている。
千葉セクション【千葉県市原市の養老川河岸に露出する地層断面】では、2010年にボーリング調査を行い、10年単位の超高解像度の古環境データを取得した。
上総層群がある千葉県の房総半島沖は、海洋プレートの太平洋プレートとフィリピン海プレートが、大陸プレートの北米プレートにぶつかる沈み込む地点にあたる。
同半島は第四紀更新世後期に隆起して陸地化したが、初期には海底の沈降域にあった。基盤の沈降速度が早く、その上に堆積した地層の堆積速度が世界最速級であったため、世界でも稀な高解像度の古海洋環境記録が取得できた。大阪湾、北大西洋の記録とあわせて、3地域で同時に起こる数百年スケールの急激な温暖化と寒冷化を多数確認された。 )
銀河宇宙線が増えれば下層雲が増え、逆に銀河宇宙線が減れば下層雲も減るので逆日傘効果で温暖化が起こる可能性もある。銀河宇宙線が下層雲の生成を誘起するというスベンスマルク効果は、銀河宇宙線が気候に影響を及ぼす可能性を示唆した仮説であるが、かつてのほとんどの気象観測データを使った検証では、銀河宇宙線量や雲量とも変化が微少なため、気候への影響を示す明確な証拠を得ることが困難であった。しかし、銀河宇宙線が大幅に増加した地磁気逆転期には、雲量の増加も大きく、日傘効果もより強くなるため気候への影響はより高感度で検出できる。
春先に見られる黄砂は、中国内陸部の砂漠地帯で巻き上げられた砂塵が偏西風に乗って日本列島にまで運ばれたものである。モンゴルとの国境付近に位置するゴビ砂漠のすぐ南の中国黄土高原では、この砂塵が過去260万年の間に厚さが最大で200mにもなる風積土 (風で運ばれ堆積した細粒物質層) として堆積している。
(砂漠や氷河で生成された岩粉が風に運ばれ堆積した黄土をレスloess 【Löß】と呼ぶ。元々がドイツ語で、氷河起源のものを指したが、日本では砂漠起源の黄砂が堆積した風積土をレスと呼ぶ。)
風が強くなれば粗い粒子がより遠くまで運ばれ、運搬量も増える。このことに着目し、地磁気逆転時に増えた雲の日傘効果で、冬の季節風が強化されたする仮説を立て黄土高原中央部の2か所のレス層の砂塵の粒度と堆積速度の変化を詳細に調査した。
その結果、両地点ともにおいて78万年前の地磁気逆転途中の約5,000年間にわたって粒度が粗くなり、堆積速度が3倍以上増加する「冬の季節風の強力化」の痕跡が発見された。この風の強力化期間は、逆転に伴い地磁気強度が1/4以下に減少し、銀河宇宙線が50%以上増加した期間と一致した。これは銀河宇宙線の増加とともに下層雲が一段と増加し (スベンスマルク効果)、その雲の日傘効果で大陸がより強く冷却されてたことを示唆している。
目次へ
チバニアン期のシベリア高気圧
偏西風Westerliesとジェット気流は、どちらも中緯度の上空で西から東へ吹く風であるが、偏西風は、北緯35~65度あたりの中緯度帯の上空1万m付近で吹いている西風のことである。日本は中緯度帯にあるため、日本の上空にも偏西風が吹いている。日本の天気が西から東へ変化するのも、この偏西風が影響している。
ジェット気流Jet Streamは、偏西風の中でも特に強い風を指す。夏よりも冬のほうが強く吹く、冬には風速が100mを超えることもある。このジェット気流は、高度9000m(300hPa)付近で観測される。
偏西風は南北方向に波打って流れており、南に張り出した部分が「気圧の谷」で、地上では低気圧が発生する。逆に北に張り出した部分は「気圧の尾根」と呼ばれ、高気圧が発生する。
地球の自転によって、コリオリの力が働き、風が向かう方向を右側に偏向させる。南から北に向かう気流は、このコリオリの力によって、どんどん右にそれていくことになり、最終的に西から東へ移動するようになる。地球の自転により、北半球では偏西風は右に曲がり、南半球では左に曲がりる。また、太陽によって赤道付近が暖められ、上昇気流が発生する。この上昇気流により、低気圧が形成され、周囲の空気が流れ込む。
シベリア高気圧Siberian Highは、冬の季節風を強め日本列島に寒気をもたらす。気圧が高いことが特徴で、現代では、冬季には1050-1070 hPa程度まで発達することもある。
シベリア気団は、地球規模で寒気が蓄積されることにより形成される。シベリアの冬季の太陽の位置は低く、ほどんど登らない。大陸の高緯度できるシベリア気団は、太陽の恩恵が乏しく気温が低い、さらに夜は放射冷却の効果でさらに冷えていき、冷たい空気は、暖かい空気と比べると密度が高いので重たい(冷たい空気の分子は集まりやすい。暖かい空気は温度が高くなるほど分子が広がる。暖かい空気が上昇して冷たい空気が下降する仕組みは、 冷たくて重い空気が暖かくて軽い空気を押しのけるから)。地表面に寒気がどんどん溜まっていく。その重たい空気が高気圧を形成する。 シベリア高気圧は背の低い高気圧と呼ばれる。
大陸で溜まりに溜まった寒気が、日本に流れ込んでくるのが寒波で、それがシベリア気団と呼ばれるものの実態で、冷たい空気は、暖かい空気と比べると重たい、 重たい空気が高気圧を形成するので、背の低い高気圧気団になる。
気圧は相対的で高気圧か低気圧かは、その気圧差の比較で決められるが、シベリア気団の気団とは、広い範囲で温度や湿度のほぼ一様な性質をもった大陸でできた空気の塊を指す。 シベリア気団では、放射冷却に加えて地表面の重たい寒気が蓄積され、その冷え切った空気の塊が、特段に冷たくて重たい高気圧を形成するから「シベリア高気圧」と特に呼ばれる。
中緯度上空を流れる偏西風が、ときおり大きく北に持続的に蛇行することがあり、それがシベリアの移動性高気圧の移動を阻害する状態が1週間程度かそれ以上にわたって続くことがある。この時、高緯度側に蛇行した偏西風は高気圧性の渦(北半球では時計回り)をつくる。これを「ブロッキング高気圧」と呼ぶ。
通常、移動性の高低気圧はジェット気流上を東進するが、「ブロッキング高気圧」が発生すると移動性の高気圧が東進を妨げられる(ブロックされる)ため、異常気象が発生し易い。この高気圧と対になって赤道側に低気圧性の渦ができることもあり、「ブロッキング高気圧」は様々な異常気象をもたらす。その予測は長期予報にとって非常に重要であるが、同時に非常に難しくなる。
「ブロッキング高気圧」は、下層から上層まで周囲より温暖になっており、長期間停滞することが多く、しかもゆっくりと西進することがある。ブロッキング現象によってシベリア上空に高気圧が停滞すると、極端な高温を引き起こし、周りの大気の流れに影響してさらなる異常気象を引き起こすこともある。2010年、ロシアの西部やシベリアを中心に猛暑となり、同国気象庁によると、1日の平均気温が平年より9~10度も高い状態が長く続き、「130年の観測史上最も暑い年になる」(フロロフ同庁長官)。緊急事態省によると、水死者は全土で昨年より倍増、6月は1244人、7月も891人に達し、計2千人を超えた。干ばつや自然火災で26の連邦構成体が非常事態を宣言した。またブロッキング現象は、気象の変化のスピードを遅くする要因にもなっている。
2010年夏には、ロシア付近の高気圧のブロッキングは、1ヶ月以 上に亘って持続した。上空では等圧線に沿って風が吹 くので、ブロッキングの西側では暖かい南風が、東側では冷 たい北風が持続する。この普段とは異なる異常気象が、なぜそんなに長く持 続するのかというメカニズムは依然として分かっていない。「20世紀最後の難問」と言われていたが、21世 紀になっても「難問中の難問」である。
「ブロッキング高気圧」にはいくつかのパターンがあり、主なものでは、高緯度側のやや東側にブ「ブロッキング高気圧」、低緯度側のやや西側に低気圧が形成され場合、偏西風が2つに分断され、高気圧側では暖かい南風が、低気圧側では冷たい北風が持続する。また、帯状に流れていた偏西風が2つに分かれ、北の分枝は高気圧性に「ブロッキング高気圧」の周りを流れ、南の分枝は低気圧性に曲がって寒冷な低気圧の周りを流れる場合があり、、この型がブロッキングの典型的なパターンである。
シベリアでは「ブロッキング高気圧」が西進する様子がみられ、シベリア高気圧の 発達と関連している可能性が考えられている。この上層の「ブロッキング高気圧」が関与して、シベリア高気圧が、日本列島に一段と厳しい寒気をもたらす冬の季節風として猛威を振るう。
 チバニアン期の境界が世界で最もよく分かる地層として認められて「国際境界模式層断面とポイント(GSSP)」と呼びれる「千葉セクション」の近くで掘削した海成層のコア試料から、10年間隔の超高解像度の古海洋環境記録が得られている。この記録と大阪湾や北大西洋中緯度の記録から、中期更新世チバニアン初期の、数mの海水準変動を伴う氷床の拡大と縮小を、500年~2000年間隔でくり返す激しい気候変化が明らかになった。北太平洋と北大西洋で同時に起こるこの気候変化の一部には、北大西洋の大量の氷山流出が関係している証拠も発見された。
チバニアン期の境界が世界で最もよく分かる地層として認められて「国際境界模式層断面とポイント(GSSP)」と呼びれる「千葉セクション」の近くで掘削した海成層のコア試料から、10年間隔の超高解像度の古海洋環境記録が得られている。この記録と大阪湾や北大西洋中緯度の記録から、中期更新世チバニアン初期の、数mの海水準変動を伴う氷床の拡大と縮小を、500年~2000年間隔でくり返す激しい気候変化が明らかになった。北太平洋と北大西洋で同時に起こるこの気候変化の一部には、北大西洋の大量の氷山流出が関係している証拠も発見された。地磁気逆転途中の5000年間、中国黄土高原では冬の季節風が運ぶ砂塵の粒が大きくなり、しかも堆積速度が3倍に増加していた。つまり冬の風が強大化していた。 上記の5000年間は地磁気強度が減少し、銀河宇宙線が50%以上増加した期間と一致する。
この冬の風の強大化は、銀河宇宙線の50%以上の増加が下層雲の増加を誘発し、その日傘効果により太陽熱が遮られ、冷えやすいユーラシア大陸が太平洋に比べより強く冷え、シベリア高気圧が強まった証拠でもある。つまり、銀河宇宙線は気候変動の要因になり、冬の季節風強化と同じ時期に、大阪湾堆積物コアの分析から寒冷化や気温の年較差の増大も起きていたことから、雲の日傘効果がこれらの主因であることがほぼ証明された。
これらの気候事象は、新生代更新世後期の7万年前から2万年前にかけての最終氷期に起こったダンスガード・オシュガー・サイクルDansgaard-Oeschger cycle(D-O)事象に特徴が似ている。D-Oイベントは数百年~3000年ほど続く温暖期(亜間氷期)と寒冷期(亜氷期)の20回に及ぶ繰り返しであった。寒冷期の事象として北大西洋における大量の氷山流出が同時に起こったこと
もあった。この寒冷期には、日本列島を含む東アジアでは、気温低下と夏季降水量の減少を併発した。
銀河宇宙線が増えれば下層雲が増え、銀河宇宙線が減れば下層雲が減るという正の相関(スベンスマルク効果)は、銀河宇宙線が地球全体の気候に多大な影響を及ぼすことを示した。主に正の電荷をもつ銀河宇宙線は地球と太陽の磁場によってシールドshieldされており、磁場が弱まれば大量の銀河宇宙線が地表に降り注ぐ。更に磁場が弱まれば大量の銀河宇宙線が地表にばら撒かれ、生命絶滅の危機を迎える。このように銀河宇宙線量を制御している地磁気と太陽磁場も、気候への影響は大きい。
チバニアン期直前の約78万年前から約77.6万年前にかけての間氷期(MIS19)では、温暖期が約1万年で終了しているので、既に温暖期が1万年以上続く完新世~現在でも、人為的な影響がなければそろそろ寒冷化して氷期に入る可能性が考えられる。
大阪湾の海底堆積物コアで見つかった酸素同位体ステージMIS19の最高海水準期付近の寒冷化は約5,000年間続いており、その期間は地磁気強度が40%以上減少し、銀河宇宙線量が40%以上増加した期間にぴったり一致した。このことから、当時の寒冷化は銀河宇宙線の増加により増えた下層雲の日傘効果が原因だと考えられている。本来なら、最高海水準期に合わせて気温もピークを取るが、この寒冷化が起こったことで少なくとも中緯度域の最温暖期は、地球磁場逆転geomagnetic reversal後、地磁気強度が40%以上に回復するまで遅らされている。それは約78万年前の最高海水準期の4,000年後の約77万6,000年前である。同様の遅れは北大西洋中緯度域の海表面水温でも起こっている。
約78万3,000年前から約77万7,000年前にかけての間氷期は、気候リズムが乱れていた。最温暖期は、約780,000年前の最高海面期から約4,000年遅れで約77万6,000年前発生しており、それも約77万7,000年前の地磁気逆転直後に関連した寒冷化が原因でこの遅れが生じた。
地磁気逆転は、約78万3,000年前から約77万7,000年前にかけての間氷期における最高海面から低海面に向かう途中で発生している。この地磁気逆転のタイミングは深海底堆積物や中国の記録と一致しており、地磁気逆転と関連している可能性が高い。地磁気逆転に伴う地磁気強度減少期と寒冷化がリンクしていることが示唆されてる。
寒冷化期間に地磁気強度が40~20%ほど減少し、銀河宇宙線量は40~90%増加したと推定されている。この重なりは、銀河宇宙線が下層雲の形成を有利にし、その日傘効果を介して寒冷化している。約77万7,000年前の地磁気逆転直後から地磁気強度が急激に回復したため、銀河宇宙線増加とそれに伴う下層雲の日傘効果が低下し、本来の温暖な間氷期が戻ったと考えられている。
チバニアン初期の、数mの海水準変動を伴う氷床の拡大・縮小を500年~2000年間隔でくり返す激しい気候変動を経た直後、突然、銀河宇宙線増加による寒冷化により日本列島を含む東アジアで、気温低下に加え、夏季降水量の減少が起こっている。それが約200年の周期で振動しながら急激に温暖化して800年後にピークに達した直後、最温暖期の最中、大量の大氷山流出が北大西洋中緯度域まで到達する大寒冷イベントを契機に、わずか50年で突然、寒冷期に戻った。太陽活動に起因すると思われる周期性を伴う急激な温暖化が、忽然と停止する現象であった。
周期性を伴う温暖化と急激な寒冷化は地磁気逆転直後に2度繰り返し、その約1万年後にもう1度繰り返した。いずれも地磁気強度が元の強さに回復した後であった。この間氷期の後半が、数mの海水準変動を伴う氷床の拡大縮小を500年~2000年間隔でくり返す激しい気候変化の時代だったことを明らかにした。
(「千葉セクション」における地磁気の逆転境界付近の地層には、ツガ・モミ・ブナ・ツツジ・イヌマキの仲間など花粉の化石が多数含まれ、現在よりも少し寒い気候が認識されている。今では日本列島で見られなくなった種、ハリゲヤキやカリヤグルミなども見つかっており、これらの花粉化石をもとに、地磁気逆転の時代、カラブリアン期末からチバニアン期初頭にかけての陸上環境を詳細に復元する研究が進められている。)
目次へ
千葉セクションChiba section
「千葉セクション」は、千葉県市原市田淵の養老川沿いに露出する地層で、約77万年前の地層を示している。この地層は、更新世の前期のカラブリアンと中期のチバニアンとの境界を示すものであり、それは地球史上最後の地磁気逆転が起きた際の事象の痕跡を留めている。
田淵の露頭で見られる地層は、海底で堆積したもので、この地層は2017年11月に「国際標準模式層断面及び地点」に内定され、2018年には「養老川流域田淵の地磁気逆転地層」として国の天然記念物に指定された。
しかも、77万年前に起こった御嶽山噴火による火山灰層(白尾火山灰層)が、地磁気逆転の時期の目印となり、その地層は「チバニアン」とも呼ばれる、中期更新世の地質時代を画している。養老川沿いの露頭では、地磁気の逆転境界付近の「白尾火山灰層」に含まれる火山灰が、現在の御嶽山付近から飛来したもので、地磁気が逆転した時期と白尾火山灰が堆積した時期が比較的近いため、地磁気逆転が起きたときの地層の位置を確認するのに適し、また、この地層からは当時の海中に生息していた微生物の化石や植物の花粉の化石も産出されており、地磁気逆転時の環境変化を解析する試料が取得できた。
「千葉セクション」および周辺の地層を対象として、地層に含まれる花粉や海洋微生物化石の分析を行い、千葉複合セクションが堆積した約80万年前から約75万年前までの6万年に及ぶ気候と海洋環境の変化が詳細に調査された。
千葉複合セクションには、この時期の、寒冷(氷期)→温暖(間氷期)→寒冷(氷期)と激しい気候と海洋環境の変動の軌跡が遺されていた。その結果、温暖期中で最も暖かかった時期の長さが約1万年間であることが明らかになった。また、最後の地磁気逆転が約77万年前に起こっていたが、千葉複合セクションでは、地磁気逆転の際、生物の明瞭な絶滅現象や気候・海洋環境の特異な変化は確認されていない。
この時代の千葉県には、ゾウやサイ、超大型のトドなど、現代では想像できない大型の動物たちが暮らしていた。
https://www.nhk.or.jp/shutoken/chiba/article/014/61/
「チバニアン」を生きた古生物 県立中央博物館で企画展
目次へ
日本海の形成
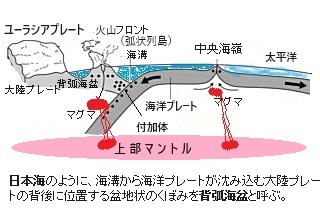
新生代古第三紀漸新世中頃〜新第三紀前期中新世中頃の約3,000万年前〜1,800万年前、海洋プレートが沈み込む海溝の西側に背弧海盆として日本海の形成が始まった。その地下深部から流動性のある高温のマントルが上昇することによって、大陸プレートの東縁の大陸縁辺地殻の一部が引き延ばされ、正断層が伸展し大陸地殻が裂け、その地下深部の割れ目から大量のマグマが噴き出して火山活動が激化した
 少なくとも漸新世(約3,400万年前~約2,303万年前)の約2,400万年前、東アジア大陸の縁辺地殻に深い溝ができ、湖や湿地が形成された。その後、裂け目が拡がり、中新世の2000万年前~1500万年前にかけて、背景海盆ができ、さらに深度を深め、2000~3000m程度の水深をもつ日本海ができた。中新世の半ば約1,500万年前に現在の位置に移動し、日本列島の骨格・弧状列島ができたと言われている。
少なくとも漸新世(約3,400万年前~約2,303万年前)の約2,400万年前、東アジア大陸の縁辺地殻に深い溝ができ、湖や湿地が形成された。その後、裂け目が拡がり、中新世の2000万年前~1500万年前にかけて、背景海盆ができ、さらに深度を深め、2000~3000m程度の水深をもつ日本海ができた。中新世の半ば約1,500万年前に現在の位置に移動し、日本列島の骨格・弧状列島ができたと言われている。中新世は、約2,303万年前から約530万年前にあたる新第三紀の第一世の地質時代である。気候は比較的温暖であったが、中新世後期(約790〜530万年前)には、東アジアの気候が夏季モンスーンから冬季モンスーンに移行し徐々に寒冷化が進行し、中新世の終わり頃には南極大陸の氷床が拡大し大陸のほとんどを覆うようになった。また、海と陸の生物相は現代に近づいていた。
(南極氷床Antarctic ice sheetは、地球上に2つある海氷polar ice packs【北極海の北極氷パックと南極海の南極氷パック】の内の一つ。南極大陸の98%を覆う氷床が、地球で一番大きい一個の氷の塊である。面積はおよそ1400万km2、体積は3000万km3、地球上にある淡水の内およそ61%を占める。
東南極の氷床は大きな陸塊に載っているが、西南極では載っている岩盤は海面下2500mよりも深い所にある。)
その後も日本海は拡大していくのだが、日本海の海洋底の拡大の仕方は少し変わっている。一般には大西洋に見られるように、大陸が分裂し、地溝帯が中央海嶺の拡大に伴って離れていく。大西洋中央海嶺は、大西洋の真中を南北に連なる海嶺で分かれているが、南北アメリカの海岸線とヨーロッパ・アフリカのそれとが合致している。アイスランドもこの海嶺の一部で、大西洋を拡大させたホットスポットのひとつである。ナンセン・ガッケル海嶺は、その大西洋中央海嶺の延長で、アイスランド北方から北極海へ延びる海底山脈で、大部分が北極圏に属するグリーンランドと北極圏のスピッツベルゲン島(ノルウェー領スヴァールバル諸島最大の島)の間を延伸する。その長さは約1800㎞で、ロシア東部の東シベリアの海岸に達している。このホットスポットとは、リソスフェアより下のアセノスフェアを生成源とする、マグマによる火山活動が起きている場所を指す。マントル上昇によって温度を保ったまま圧力が減少するため、マントルに部分的な溶融が起こり、マグマが発生する。
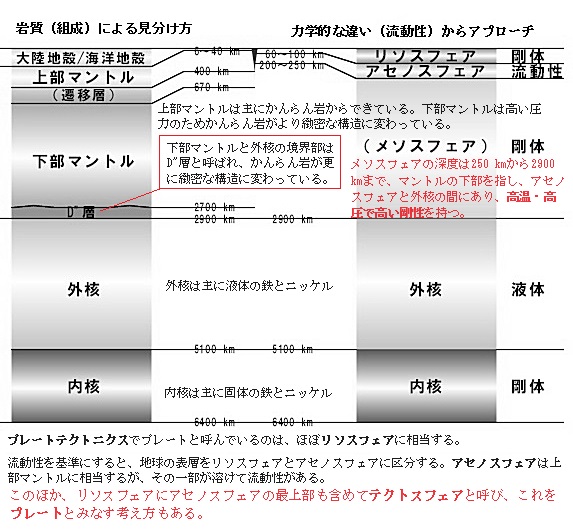 地球の内部構造を理解するには、どんな物質でできているかという岩質 (組成) による見分け方と、剛体であるかないかという力学的な違い (流動性) からアプローチする。
地球の内部構造を理解するには、どんな物質でできているかという岩質 (組成) による見分け方と、剛体であるかないかという力学的な違い (流動性) からアプローチする。上部マントルは主にかんらん岩からできていると考えられている。下部マントルは高い圧力のためかんらん岩がより緻密な構造に変わっていると考えられ、上部マントルと下部マントルの間は漸移帯 (遷移層) がある。
下部マントルと外核の境界部はD"層と呼ばれ、かんらん岩が更に緻密な構造に変わっている。
外核は主に液体の鉄とニッケルから、内核は主に固体の鉄とニッケルからできている。
アセノスフェアは、リソスフェア(プレート)の下位にあって、部分的に融解している物質であるため、比較的流動性に富んだ軟らかい層なので、岩流圏とも呼ばれる。
アセノスフェアは、リソスフェアと違い流動しやすい特性があるため、リソスフェアと対比される。アセノスフェアの下部で接する上部マントルには、地震波の速度が深さと共に減少するいわゆる低速度層として存在するが、アセノスフェアはこの層とほぼ同一のものと考えられている。
低速度層は、大陸下よりも海底下でより顕著に存在することが地震波の観測から知られている。この層は海底下では深さ70~250kmくらいに存在するが、大陸下ではほとんど認められない。
しかし、日本などの島弧下では低速度層は異常に発達しており、地殻直下(深さ約30km)にまで及んでいる。このようなアセノスフェアの発達状況の違いは、上部マントルとリソスフェア(プレート)の間に位置する層であり、上部マントルは主にかんらん岩からできており、アセノスフェアはその一部が溶けて流動性を持っていると考えられている。マントルは、高温のために部分溶解しているか、それに近い軟弱な状態にあると考えられている。アセノスフェアは、リソスフェアよりかなり軟らかいためリソスフェア(プレート)が運動するのを容易にしていると考えられている。リソスフェアは、地球表面を覆う硬い層で、地震発生や火山噴火のメカニズムに重要な役割を果たしているが、そのリソスフェアはアセノスフェアの上を流動している。
 日本海は、大陸縁辺部の地殻の伸長と薄化によって始まった海盆で、日本海の東側を画する大規模な横ずれ断層から拡大が開始された。その後も伸長を続ける大陸性地殻の東縁で西方へ伸長していく新生代古第三紀漸新世の約2800万年前の日本海の拡大開始は、ユーラシア大陸縁辺部の薄化と島弧地殻の伸長よるものであった。
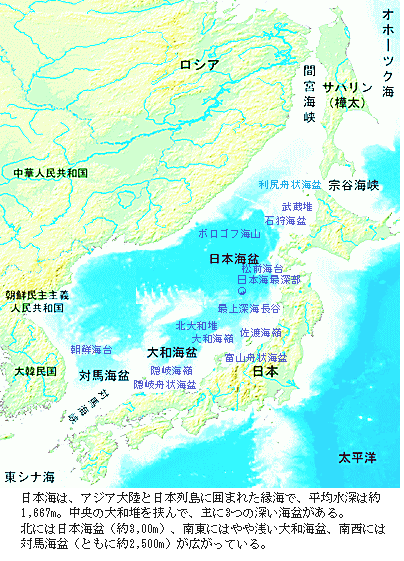
日本海は、大陸縁辺部の地殻の伸長と薄化によって始まった海盆で、日本海の東側を画する大規模な横ずれ断層から拡大が開始された。その後も伸長を続ける大陸性地殻の東縁で西方へ伸長していく新生代古第三紀漸新世の約2800万年前の日本海の拡大開始は、ユーラシア大陸縁辺部の薄化と島弧地殻の伸長よるものであった。日本海は、大陸と日本列島に取り囲まれた縁海である。ユーラシア大陸と樺太の間の間宮海峡(タタール海峡)、樺太と北海道の間の宗谷海峡でオホーツク海と繋がり、北海道と本州の間の津軽海峡では太平洋と、九州と対馬の間の対馬海峡東水道、対馬と韓国の間の対馬海峡で東シナ海と繋がっている。
海底地形は、日本列島沿岸海域の大陸境界地、すなわち小規模な海底山脈(海嶺)と海盆とが複雑に組み合って分布する海域と、大陸性地殻からなる大和海嶺、朝鮮海台などの高所、そして日本海北部に代表される深海盆に大別できる。
日本海の北半分を占める日本海盆の最深部は約3,796mで、その日本海盆底は主に玄武岩からなり、約3,000万年前に形成されたと推定されている。周りの陸地から海底谷を通って流れ込んだ陸性堆積物が厚さ2000m も積もり、著しく平坦であるが、西部はやや浅くなっている。
約2800万年前に日本海の東縁を画する大規模横ずれ断層では、海洋地殻と上部マントルの一部も含むリソスフェアの最初の断裂が起こり、そこから海底拡大が開始された。この海底拡大は新第三紀中新世の約1800万年頃まで西および南西に伝播し、日本海盆の東半分を海洋性地殻で形成した。一方、日本海南西部では地殻伸長・薄化により陸性地殻の断片からなる海嶺・海台群と海盆群が形成された。海嶺は海洋底で海盆を分ける細長い山脈状の地形で、海嶺により新しい海洋地殻が形成される場所であるばかりか、海底が生まれる場所でもある。日本海盆には海嶺・海台群と陸性地殻の伸長・薄化により生じた海盆群が存在する。
 日本海が形成され、これに伴う海底火山活動で日本各地にグリーンタフgreen tuffと呼ばれる凝灰岩層が発達した。緑色凝灰岩とも呼ばれる、その緑色や緑白色や淡緑色などは、産出場所によって、その色調は異なる。重要なのは、グリーンタフが緑色を呈する理由が、岩石に含まれる輝石・角閃石などの造岩鉱物が、熱水による化学反応で粘土鉱物の1種の緑泥石に変化したことにある。しかも、グリーンタフは、日本列島以外にもアジア東部の大陸地殻にも存在し、約2600万年前から約500万年前にかけて「グリーンタフ造山運動」と呼ばれる造山運動が起きたことによる。この造山運動は、日本列島を含む太平洋周辺地域で特に顕著で、地殻の沈降と堆積が初期段階で支配的であったが、後半には褶曲と隆起が著しくなり、日本列島の形成に大きく寄与した。
日本海が形成され、これに伴う海底火山活動で日本各地にグリーンタフgreen tuffと呼ばれる凝灰岩層が発達した。緑色凝灰岩とも呼ばれる、その緑色や緑白色や淡緑色などは、産出場所によって、その色調は異なる。重要なのは、グリーンタフが緑色を呈する理由が、岩石に含まれる輝石・角閃石などの造岩鉱物が、熱水による化学反応で粘土鉱物の1種の緑泥石に変化したことにある。しかも、グリーンタフは、日本列島以外にもアジア東部の大陸地殻にも存在し、約2600万年前から約500万年前にかけて「グリーンタフ造山運動」と呼ばれる造山運動が起きたことによる。この造山運動は、日本列島を含む太平洋周辺地域で特に顕著で、地殻の沈降と堆積が初期段階で支配的であったが、後半には褶曲と隆起が著しくなり、日本列島の形成に大きく寄与した。グリーンタフのグリーンは緑色を指す。タフとは火山灰が固まってできる凝灰岩を指す。つまり造山運動による変質作用によって生成された緑色を示す変成鉱物である。
一口に緑色とは言っても、緑色や緑白色や淡緑色など、産出場所によって、その色調は異なる。グリーンタフが緑色を呈する理由は、岩石に含まれる輝石・角閃石などの造岩鉱物が、熱水の影響による変質により粘土鉱物の1種の緑泥石に変化したことによる。
輝石は火成岩および変成岩で見つかり、角閃石は主に火成岩で見られる。特に苦鉄質の火成岩によく含まれ、マグマが固化してできた岩石であることを示す。代表的な石材としては、栃木県宇都宮市北西部の大谷町付近一帯で採掘される大谷石、秋田県大館市で採掘されてきた十和田石、伊豆で採掘されてきた伊豆石、福井県で採掘されていた笏谷石(しゃくだにいし)や滝ケ原石などがある。
目次へ
 そこから先、日本のゾウはどうなったのかは、実は、よく分からない。中新世後期になる地球寒冷化が進行し、約1600万年前から600万年前までの間、ゾウに限らず陸上生物の化石記録がほとんど産出しない。中新世の終わりには氷床が大陸のほとんどを覆うようになる。生物相も現代に近づき、ヒト科もこの時代に現れた。中新世の終わり(
そこから先、日本のゾウはどうなったのかは、実は、よく分からない。中新世後期になる地球寒冷化が進行し、約1600万年前から600万年前までの間、ゾウに限らず陸上生物の化石記録がほとんど産出しない。中新世の終わりには氷床が大陸のほとんどを覆うようになる。生物相も現代に近づき、ヒト科もこの時代に現れた。中新世の終わり(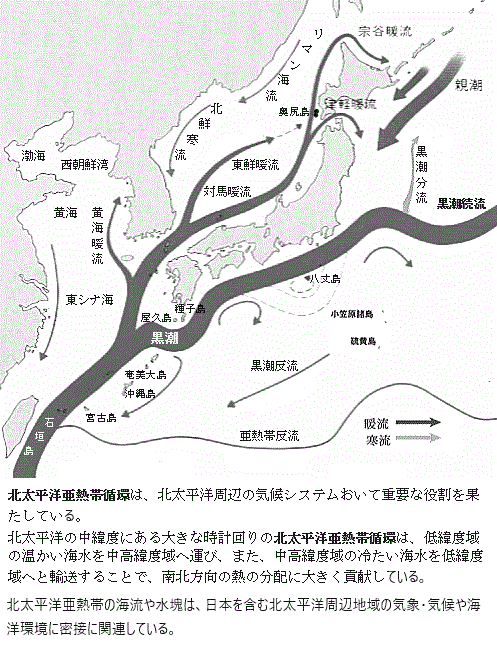 黒潮続流の北側は、この低温・低塩の海水と亜熱帯起源の高温・高塩の海水の両方が存在する海域のため混合水域と呼ばれる。黒潮続流は、その混合水域の南の境界側と見られる。
黒潮続流の北側は、この低温・低塩の海水と亜熱帯起源の高温・高塩の海水の両方が存在する海域のため混合水域と呼ばれる。黒潮続流は、その混合水域の南の境界側と見られる。 中緯度上空を流れる偏西風は、ときおり大きく南北に持続的に蛇行することがある。この蛇行が持続すると、気温や降水が平年とは異なる状態が続くことで異常気象をもたらす重要な要因の一つになる。この蛇行が高緯度側にひときわ大きく起こっている場所ではブロッキング高気圧が形成される。
中緯度上空を流れる偏西風は、ときおり大きく南北に持続的に蛇行することがある。この蛇行が持続すると、気温や降水が平年とは異なる状態が続くことで異常気象をもたらす重要な要因の一つになる。この蛇行が高緯度側にひときわ大きく起こっている場所ではブロッキング高気圧が形成される。