TOP 車山お知らせ 諏訪の歴史散歩 高島藩二之丸騒動
| 目次 1)界門綱目科属種 生物の分類階級、ドメイン・界・門・綱・目・科・属・種について 2)ドメインdomain 六界説 : 動物界・植物界・原生生物界・菌界・真正細菌界・古細菌界 生物には無限と言っても誇張にならないほどの膨大な種があり、それぞれには独自の特徴や役割がある。 人類は、苛酷な生存競争の試練に耐え、漸く生物間の相互扶助を知る。 例えば、動物・植物・菌類・原生生物・細菌・古細菌のように、さまざまな界に分類されるが、 各界にはさらに細かい分類があり、それぞれが生態系の中で相互に関連し合い重要な役割を果たしている。 |
 2024年5月8日 諏訪湖 高島城と藤 2024年5月8日 諏訪湖 高島城と藤 |
6界説では、生物を次の6つの界に分類する。
① 動物界Animalia : すべての動物が含まれる。
(従属栄養生物Heterotroph【other nutritionの意味】はエネルギー源の供給を他種生物に依存する生物であり、
動物はこのカテゴリに入る。
寄生植物などは例外である.が、独立栄養生物の植物のように光合成を行うことができないため、
動物は他の生物から栄養を摂取しなければならない。
草食動物【一次消費者】は植物を食べ、肉食動物【二次および三次消費者】は他の動物を食べることで
エネルギーを得ている。
従属栄養生物としての動物は、生態系のバランスを保つ重要な役割を果たしている。
食物連鎖の中でエネルギーを伝達し、生態系全体の健全性を支えている。
従属栄養生物には多様な形態があり、微生物・菌類・寄生植物などが含まれる。
動物はこの広い中で特異な存在であり続け、その役割や特徴とその影響は、生態系の中で非常に際立つ。
特に動物である人類は、極めて破滅的である。しばしば「神の名において」人類を壊滅させようとする。)
② 植物界Plantae : すべての植物が含まれる。
(植物は基本的に自己栄養性で、光合成を行うことでエネルギーを獲得する。)
③ 菌界Fungi : キノコやカビ、酵母などの真菌類が含まれる。
(真菌類とは、カビの総称で、核膜に囲まれた核を持つ真核生物である。
真菌には、酵母型・菌糸型・二形性の3つの形状が存在する。
酵母型は単細胞性で、二分裂や出芽によって増殖する。
菌糸型は細胞が伸びて糸状や円筒状になる。菌糸型は主にカビやキノコなどの真菌に見られる成長形態。
この形態では、個々の細胞が伸びて連なり、糸のような構造を形成する。
これを「菌糸(きんし)」と呼ぶ。 菌糸は個々の細胞が連続的に成長して、長い糸状や円筒状になる。
この連続した細胞の並びは、一つの大きな構造として機能する。
菌糸の中にはしばしば隔壁と呼ばれる細胞壁を有し、個々の細胞を分けている。
この隔壁には小さな孔があり、細胞間で物質がやり取りされる。
菌糸は土壌や腐敗した有機物の中に広がり、そこで栄養を吸収する。
菌糸の表面積は大きいため、効率的に栄養を取り込むことができる。
しかも、そのクモの巣のような構造で環境中に広がり、クモの巣のように複雑なネットワークを形成する。
二形性は、条件によって酵母型と菌糸型の両方の形態をとることができる)
④ 原生生物界Protista : 単細胞または比較的単純な多細胞の真核生物が含まれる。
真核生物は核膜を持つ。核膜は、核を包み込む二重膜により、細胞質から核を隔離している。
この核の中には遺伝情報を含むDNAが含まれている。
真核生物には、動物・植物・菌類・原生生物などがあり、多細胞生物と単細胞生物の両方が存在する。
核膜の二重膜構造は、DNAを外部からの物理的な損傷や有害物質から守り、
核内の環境を細胞質から隔離し安定させる。
また、核以外の細胞質には、ミトコンドリア・リソソーム・ゴルジ体・リボソームなどの細胞小器官や
それ以外の細胞膜・細胞骨格・細胞溶液などの細胞質が、
細胞の内部を満たし、物質の輸送・細胞の形状維持・化学反応の場を提供するなど、さまざまな役割を果たす。
リソソームは、真核細胞内に存在する小さな膜で囲まれた小器官で、
その役割は、細胞内の不要な物質や壊れた細胞小器官、外部から取り込まれた異物などを分解して処理する。
リソソーム内には多種多様な酵素が含まれており、これらの酵素がタンパク質・脂質・核酸などの分解する。
これにより、リソソームは細胞内の清掃とリサイクルを行い、細胞の正常な機能を維持する。
核膜は染色体の高次構造の形成に重要な役割を果たしている。
核ラミナは、核膜の内側に形成される均一なメッシュ構造である。
核の二重膜はこの核ラミナと結びつくことで、より強固な構造になる。これが核の形状を維持する。
これにより、遺伝情報を含むDNAが核内に守られ、
二重膜にある核膜孔を通じて、RNAやタンパク質などの物質が選択的に出入りできる。
これにより、必要な物質だけが核内外を行き来できるように調整される。
また必要なときにだけ細胞質へ情報が伝達される。
核膜には、核膜孔と呼ばれる小さな穴があり、これを通じて物質の出入りが行われる。
例えば、mRNA(メッセンジャーRNA)は核膜孔を通って核外に出され、細胞質でタンパク質合成を担う。
DNAは主に細胞の核内に存在する。核はDNAを保護し、遺伝情報の保存と制御を行う中心的な場所である。
RNAは、その種類によって細胞内の異なるそれぞれの場所で活動する。
mRNA(メッセンジャーRNA)は核内でDNAから転写され、細胞質へと移動してタンパク質合成に関与する。
rRNA(リボソームRNA)やtRNA(転移RNA)は細胞質内のリボソームで働く。
つまり、DNAは核内に存在し、RNAは核内と細胞質の両方に存在し、
特にタンパク質合成の過程では細胞質で重要な役割を果たす。
(原生生物界には、多様な生物が含まれており、それぞれが独自の特性を持ってる。
原生生物は、単細胞生物や単細胞の集合体から成り、多くは湿った環境や水中に生息している。
アメーバは、単細胞の微生物で、水や土壌、人体などさまざまな環境に適応する。
仮足を伸ばして移動し、細胞質を流動的に変形させることで、摂食や移動を行う。
アメーバは非常に多様なグループであり、様々な種が異なる生態や生殖戦略を持つ。
例えば、病原性を持つ「赤痢アメーバ」や、自由生活をする非病原性のものもある。
その「自由生活アメーバ」は、病気を引き起こさず、環境中で自由に生活する。
例えば、アメーバ・プロテウスやディフフジア・コロラタなどの「自由生活アメーバ」は土壌や水中、
そして空気中など、さまざまな環境で生息している。
「赤痢アメーバ」とは異なり、直接的に人間や動物に害を及ぼすことはない。
「自由生活アメーバ」は、多くの場合、変形性を持ち、形を変えることで、
細かい隙間を通り抜けて、エサを捕まえる。
アメーバ運動と呼ばれる独特の方法で、偽足pseudopodiaと呼ばれる突起物を伸ばして
這うように移動して、主に細菌や小さな有機物を捕食する。
偽足を使って餌を包み込み、細胞内に取り込む。
「自由生活アメーバ」は、湿った土壌や水たまり、湖や川などの淡水環境、時には塩水環境にも適応する。
多様な環境下で生息し、一部の種では空気中や樹木の樹皮などでも見られる。
これらのアメーバは、生態系内で重要な役割を果たしている。
例えば、土壌中の細菌を捕食することで、栄養素の循環を促進したり、
水中ではプランクトンの一部として他の生物の餌となることもある。
無性生殖が一般的で、主に分裂によって増殖するが、一部の種では、
非常に稀であるが有性生殖も報告されている。
アメーバの「異食性」とは、他の生物や有機物を捕食する習性のことを指す。
アメーバは、光合成を行って栄養を得るのではなく、
他の微生物や有機物を捕らえて食べることで栄養を摂取する。
アメーバの異食性が、さまざまな環境で生き残るための重要な要素となっている。
例えば、湖や川の底にある有機物・細菌・藻類などを食べることで、栄養を確保する。
彼らは偽足(ぬか足とも呼ばれる)を使って獲物を包み込み、食胞と呼ばれる小胞に取り込む。
偽足により、捕食や適応環境の探索が可能となる。
食胞内で酵素による消化が行われ、栄養が吸収される。
水中環境を好むが、湿った土壌や動物の腸内など多様な場所で生息する。
乾燥や栄養不足などの不利な条件下では、アメーバは被胞(シスト)を形成し 被胞内で活動を停止し、
環境が改善するまで休眠状態を保つ。
アメーバは主に無性生殖、特に二分裂によって増殖する。
一つのアメーバが成長し、細胞核が分裂し、続いて細胞質が分裂することで2つの新しいアメーバが生まれる。
ただし、特定の条件下では有性生殖に関連する現象が観察されることもある。
例えば、栄養不足や環境ストレス時には、遺伝物質の交換が行われることがある。
これは一般的に真の有性生殖とは区別される。
ゾウリムシは単細胞の原生動物で、池・湖・川などの淡水環境に広く分布し、環境の変化に対する適応力が高い。
ゾウリムシは水中だけでなく、湿った環境でも生存できる。
昆虫や小動物が泥や植物を運ぶ際に、ゾウリムシがその体表に付着して新たな場所に移動することがある。
風によってゾウリムシの一部が移動し、雨の溜水に落ちることもある。
雨の溜水は一時的な生息地であるが、新たな環境に入っても迅速に増殖し、生息地を広げることができる。
環境が乾燥しても、耐久性のある胞子やシストcystとして生存し、次の湿潤時期に再び活動を開始する。
シストは厚い保護膜で覆われており、外界からのダメージから、守られる。
その内部で、ゾウリムシは代謝を抑え休眠状態に入る。
腐敗した植物質の中にも生息する。繊毛【シリア】を使って移動し、
細胞口(孔口)を通して小さな食物粒子を取り込む。
取り込まれた食物は食胞と呼ばれる細胞小器官内で消化される。
通常は無性生殖によって二分裂し、新しい個体を形成するが、
環境が厳しくなると有性生殖を行い、遺伝的多様性を高める。
緑藻green algaeは光合成を行い、植物の祖先とされる。藻類の中でも非常に多様性があり、
海洋から淡水環境まで広く分布し、土壌などにも生息する。
緑色植物のうち、陸上植物を除いたものの総称して緑色藻とも言う。
緑藻は、多様な種類が存在し、緑藻の中には、単細胞のものから、
群体・多細胞・多核嚢状(多核藻)のものまで多様な種が含まれ、それぞれ独自の生態を持っている。
多核体Coenocyteは、大型の細胞に多数の核が含まれた状態のことで、原生生物において多数の例がある。
一般に細胞は1つの核を持ち、細胞が成長すると核が分裂し、同時に細胞質も分裂するが、
ある種の生物では細胞が成長すると、核は分裂するが細胞質は分裂せず、結果として複数の核を持つ細胞が生じる。
これを続けていけば、巨大な細胞膜の細胞質の中で多数の核が存在する状態を生じる。これが多核体である。
多核体の緑藻は「多核藻」とも呼ばれる。細胞小器官などは、通常の細胞と共通であり、
1つの細胞の振る舞いとして観察される。
多核藻
藻類の大きな多核細胞は、単一の細胞に複数の核が含まれている合胞体構造で、この生体はコエノサイトcoenocyteとして知られている。コエノサイトは、藻類の大きな多核細胞で、細胞内に複数の核が集合し、細胞の成長や形態形成に関与する。この過程で、核が細胞内を動き回り、特定の位置に集まることで、細胞の成長点や鞭毛などの.新しい構造の形成が促進される。この現象は、特に光合成によるエネルギー供給や遺伝子発現に関与しており、多核細胞の特性を活かした形態形成が行われている。例えば、フシナシミドロVaucheriaなどの藻類では、青色光の照射に応答して核が集合し、正負の光屈性、光成長反応、光照射域での成長点の誘導を示すことが知られている。
緑藻多核体の「正負の光屈性」の運動は、光の強さや方向に影響される。例えば、緑藻は光源に向かって移動し、成長する。これは光合成を最適化するための反応と言える。他方、「負の光屈性」反応は、一部の細胞小器官が光源から遠ざかる方向に移動する。 この動きは、光受容体によって光を感知し、それに応じて細胞の成分や運動の方向を最適化する。光受容体が活性化されると、特定のホルモンやシグナル伝達経路が作動し、細胞の動きを変える指示が出される。
緑藻は浅所に適応しているが、ある種の緑藻は浅所で見られず、沿岸部の深所に分布している。沿岸部深所の光は一般に緑色であり、そのような環境に好んで生える緑藻は、浅所の白色光ではなく外洋の海中の青色光も効率よく利用する。緑藻の合胞体構造は、外洋の海中にも存在する。緑藻の多核体には非常に多様な藻類があり、淡水と海水の両方で見られるが、外洋にも適応して生息している種があり、その中には合胞体構造を持つものも含まれている。その合胞体構造は、細胞壁が融合して巨大な多核細胞を形成する特異な構造であるため、栄養の交換や環境への適応において有利な形態とされている。
緑藻の合胞体構造におけるこのステート遷移state transitionでは、通常の細胞の集合体とは異なる形態や機能を示す。合胞体は単細胞の状態から多細胞の状態に形態を変化させることがある。これにより、細胞内の相互作用や機能が変化する。
合胞体細胞内自体では特定の機能を分化させるようになる。例えば、細胞内の一部では光合成を行い、その一方、その細胞内で栄養分の輸送や代謝を担うなど統制された分業が達成されている。
: 緑藻の合胞体では、環境条件に応じて異なる生活環を持つことがある。例えば、乾燥や低温などのストレス条件に対する適応として、合胞体の形成が促進されることがある。このステート遷移では、緑藻が生存と繁殖を維持するために重要な役割を果たす。
ステート遷移には、青色光や赤外光などの特定波長の光を受けた際に、2つの光化学系(光化学系Iと光化学系II)の励起バランスを補正する仕組みもある。緑藻もステート遷移を行うことが明らかになったことで、このステート遷移が従来知られていた陸上植物などのものとは異なり、青緑色光を利用する利点である。青緑色光は海中を満たしている。植物の祖先は海中で誕生し、やがて陸上へ進出した。ステート遷移の起源が、植物がまだ海中に生息していた時代にまで遡ること、そして当初から海中で光合成調節メカニズムが機能していたことを示唆している。多細胞植物が新しい形を作るには、分裂組織を作って細胞数を増やすしかないが、多核細胞は核を寄せて形を整え対応する。
またフシナシミドロは、隔壁がない糸状体、つまり、フシナシミドロの細胞は隔壁がないため、糸状体が1つの連続した細胞として機能する。その糸状体は時々分枝し、まばらに広がる。その表層部には多数の核が存在し、それぞれの核は細胞内に分散し、核の数に相当する緑色の葉緑体が存在し、光合成を行う。
多核体のカウレルパCaulerpaは、カウラーパ科(緑藻類)の海藻の属で、それらは、多くの核を持つ単一の細胞のみで構成されているため、世界最大の単一細胞の1つになっている。これは、フシナシミドロ属やカウレルパ属に属する藻類など、特定の種類の藻類で発生する。これらの藻類は、細胞分裂を別々の細胞ですることなく、大きくて複雑な機能を働かせる生物として成長することができる。これはほとんどの多細胞生物とはかなりかけ離れて居る。
多核細胞multinucleate cellsは、複数の核を持つ、つまり複数の核が1つの共通の細胞質を共有する真核細胞で、多核細胞の有糸分裂は、すべての核が同時に分裂する協調的で同期的な方法で起こるか、個々の核が時間と空間それぞれで独立して分裂する非同期的現象も見られる。
また多核藻multinucleated algaeの生殖は、他のタイプの藻類とは異なる。多核体の緑藻には複数の核があるため、その生殖方法には無性生殖と有性生殖の両方が行われている。
カウレルパCaulerpaも、多核体の藻類であれば、海洋環境に広く分布し、無性生殖と有性生殖の両方を行う。カウレルパの無性生殖は、主に破片化や葉状体の分裂によって無性生殖を行う。この破片化は、葉状体が破断し、それぞれが新しい個体として成長する。葉状体の分裂は、葉状体内の細胞が分裂し、新しい個体を形成される手段である。
カウレルパの有性生殖は、配偶子の形成によって行われる。雌雄の配偶子が形成され、受精することで新しい個体が生まれる。この過程は、他の多くの藻類と同様に、温度や光の条件に依存する。
多核体のフシナシミドロVaucheriaも、無性生殖と有性生殖の両方を行う。フシナシミドロの無性生殖は、胞子形成sporangium formationによって行われる。無性胞子aplanosporesや遊走子zoosporesと呼ばれる胞子が形成され、これが新しい個体に成長する。これらの胞子は、特定の環境条件、光・温度・栄養状態などが整うと発芽し、新しい藻類体を形成する。
カウレルパやフシナシミドロなどの合胞体の中の葉緑体chloroplastの数は、通常、核の数とは直接対応していないことが多い。合胞体の多くの核を持つ細胞質内で、クロロプラストは分散している。核の数に比例してクロロプラストの数が増減するわけではなく、それぞれ独立して調整されている。
有性生殖は、配偶子形成gametangium formationを通じて行われる。フシナシミドロは、異形配偶子oogamousを形成する。雌性配偶子(卵細胞)と雄性配偶子(精子)がそれぞれの配偶子嚢内で形成され、受精によって新しい個体が生まれる。受精卵zygoteは、環境が適した時に発芽し、新しいフシナシミドロ個体となる。
これらの生殖方法により、多核体の緑藻は効果的に繁殖し、環境の適応範囲を広げる能力を高める。特に、有性生殖によって生成された多様な遺伝子組み合わせは、環境変動に対する強い耐性能力が付加される可能性が高い。
 緑藻の生態と光合成のプロセスは非常に興味深い。
緑藻の生態と光合成のプロセスは非常に興味深い。光合成は、緑藻がクロロフィルを利用して光エネルギーを吸収して、二酸化炭素と水から酸素と有機物(糖)を生成する化学反応である。
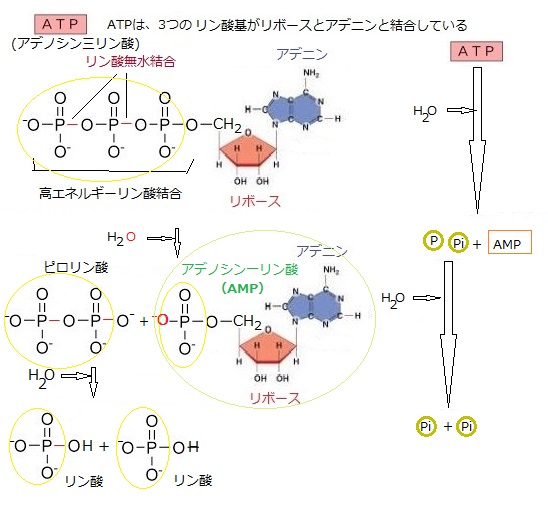
また捕らえた光エネルギーはATP(アデノシン三リン酸)の生成に利用される。
ATPは細胞のエネルギー通貨として機能する。
二酸化炭素を固定して生成したATPとNADPHを利用して、二酸化炭素が有機物(主に糖)に固定される。
このプロセスはカルビン・ベンソン回路と呼ばれる。
緑藻の光合成は、地球の酸素供給の一部を担っており、
水域のエコシステムにおいても重要な役割を果たしている。
クロレラ Chlorellaは、淡水に生息し、円形の単細胞藻類で、栄養価が高く、健康食品としても利用される。
クラミドモナスChlamydomonasは、クロレラと同じく単細胞で、淡水に生息している。
鞭毛を持ち、泳ぐように運動する。
珪藻は「フラストゥルム」と呼ばれるシリカ(SiO2)を含む細胞壁を持つ単細胞の藻類。
海洋や淡水に広く分布している。
このフラストゥルムは二つの殻が合わせて形成されており、ペトリ皿のような構造を持っている。
多様な形態を持ち、種によって形や模様が異なる。その美しい形状は顕微鏡観察の楽しみの一つと言う。
珪藻は光合成を行い、二酸化炭素と水から酸素と有機物を生成し、その酸素を放出により、
水中の生態系において重要な役割を果たす。
また珪藻は他の多くの水中生物にとって重要な栄養源となる。
プランクトンとして存在し、魚や他の水生生物の食物連鎖の一部を構成する。
粘菌は多核体のアメーバ状の細胞を形成し、分裂や融合を繰り返す。湿った環境で腐植物を分解する。
細胞性粘菌は、バクテリアや酵母菌を餌とする。まわりにその餌があるとき、独立したアメーバ細胞として
餌を盛んに食べて、分裂して数を増やす。
このとき、アメーバ細胞は自由に動き回ることができ、動物細胞のように振舞う。
単細胞生物として生活するが、餌がなくなると一部の細胞がある化学物質を出すと、
その細胞に向かっておよそ10万個の細胞が集まって1つの集合体を作る。
さまざまな研究から、このときの集合体は単なる細胞の集まりではなく、
ヒトのような多細胞生物が持つ機構のほとんどを持つので、多細胞生物ということができる。
集合体は条件の良い場所をもとめて、ナメクジ構造を形成して移動する。
卵菌Oomycetesは、水中や湿った土壌で見られる。菌糸を形成し、植物の病原菌として知られている。)
⑤ 真正細菌界Bacteria : 細菌を含む一群で、細胞膜はあるが、単細胞の原核生物。
原核生物は核膜を持たない。遺伝情報は環状のDNA分子として存在し、殆どの細菌やシアノバクテリアを含む。
ミトコンドリアやゴルジ体などの膜に囲まれた細胞小器官は無いが、リボソームは存在する。
(真正細菌界は非常に多様で、さまざまな種類が存在し、それぞれ独自の特徴を持っている。
彼らは環境や産業、生態系において重要な役割を果たしている。
例えば、藍藻は地球の酸素供給に貢献し、
偏性好気性菌や嫌気性菌は土壌の栄養循環に寄与している。
真正細菌は驚くべき適応力を持ち、多様な環境で生き抜く能力がある。
藍藻は、光合成を行い、酸素を生成する。淡水や海洋、土壌に広く分布し、植物の祖先とされている。
ブドウ球菌・連鎖球菌・放線菌などは、多くは抗生物質を生成する能力を持つ。
大腸菌・サルモネラ菌・ネイセリア菌など外膜は、リポ多糖を含み、免疫系からの防御機能を強化する。
結核菌は、酸素がない環境では生存できない。
破傷風菌などは、酸素を嫌い、酸素の存在下では生存できない。
大腸菌は酸素があってもなくても生存できる菌。)
⑥ 古細菌界 Archaea : 真正細菌とは異なる原核生物の一群。
 |
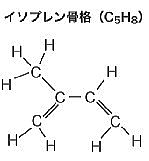 |
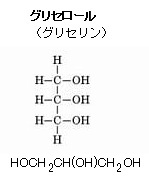
| グリセロールの役割は、リン脂質やトリアシルグリセロールなどの脂質分子の骨格として機能し、細胞膜の構造を形成する。 リン脂質は細胞膜の二重層を構成し、膜の流動性と透過性を調整する。 グリセロールはヒドロキシル基(化学式は-OH)の存在により、水分を引き寄せる性質が生じ、細胞膜の保湿を維持する。これにより、細胞膜の機能を正常に保つための柔軟性が加わる。 グリセロールは、脂質代謝の過程で分解され、グルコースやATPの合成に利用されるため、細胞のエネルギー供給に重要な役割を果たす。 一部のグリセロール誘導体は、細胞間のシグナル伝達に関与し、細胞の応答や適応に寄与する。これらの役割を通して、グリセロールは生体膜の構造と機能の維持に欠かせない要素になる。 化学式は-OHのヒドロキシル基は、水素原子と酸素原子から成る化学基で、この基は多くの有機化合物や無機化合物に見られる。例えば、アルコール(エタノールなど)はヒドロキシル基を含む代表的な有機化合物である。 ヒドロキシル基は、分子に付加されることでその物性や化学的性質を大きく変えることができる。例えば、ヒドロキシル基が付加されることで、化合物の溶解性が増すこともある。 古細菌の脂質単層は、特殊なイソプレノイド鎖脂質から成り立っており、これらはグリセロールの両端に結合している。具体的には、脂質単層の基本単位は、グリセロールの中央に結合したビホスファタチジル基(化学式:C10H21N5O7P2)と、両端に結合したイソプレンisopreneは、構造式CH2=C(CH3)CH=CH2のモンマーであり、イソプレノイドの分子式はイソプレンの倍数であるから(C5H8)nで表される。 二重結合を2つ持つ炭化水素化合物のイソプレノイド鎖は、直鎖ではなく、分岐した構造を持つため、膜の流動性と安定性が増す。 イソプレノイド鎖はグリセロールにエーテル結合で結合しており、この結合はエステル結合よりも化学的に安定する。 |
極限環境に適応するための特異な特性を持ち、地球上の生態系において重要な役割を果たしている。
その研究は、生命の起源や地球外生命の可能性についての洞察を提供する可能性がある。
細菌と混同されがちであるが、遺伝的および生化学的に異なる存在である。
真正細菌界や真核生物とならんで、ひとつの古細菌界domainを形作る古細菌は、
進化系統樹としては、「核膜が無いDNAが細胞質内にむき出しの原核生物の共通祖先から、真正細菌と原始古細菌に進化し、その後、原始古細菌から核膜を持たない単細胞の原核生物のままの古細菌界と核膜を持つ真核生物に分かれ進化した」)
メタン生成古細菌Methanogensは、無酸素環境(嫌気環境)でメタンを生成する。
沼地や動物の腸内、下水処理施設などに生息している。
そのメタン生成はエネルギー代謝の一環として行われ、これは炭素循環において重要な役割を果たす。
極限好塩古細菌Halophilesは、高塩濃度環境に適応し、塩湖や塩田などに見られる。
高濃度の塩に耐えるための特殊な代謝経路を持ち、細胞内外の塩濃度を調整する能力がある。
好熱酸性古細菌Thermoacidophilesは、高温かつ酸性の環境に適応し、温泉や火山地帯、深海熱水噴出口などに生息する。
高温と酸性度に耐えるためのタンパク質や細胞膜の特殊な構造を持っている。
極限好圧古細菌Piezophilesは、高圧環境に適応し、深海の海底などに生息している。
高圧に耐えるための細胞膜や酵素の特殊な構造を持つ。
超好熱古細菌Hyperthermophilesは、極端な高温環境(80℃以上)に適応し、
深海熱水噴出口や火山地帯の熱水泉などに見られる。
高温に耐えるための耐熱性タンパク質や酵素を持っている。
古細菌(アーキア)は、光合成・メタン生成・硫黄代謝など、さまざまな代謝経路を持つ。
古細菌の細胞膜は、その特殊な化学構造と機能によって特徴付けられる。
その細胞膜は、一般的な細菌や真核生物の細胞膜とは異なる。
古細菌の膜脂質はエステル結合ではなく、エーテル結合によって
グリセロール(グリセリン)と脂肪酸が結びついている。
グリセロールの両端に長いイソプレノイド鎖(イソプレン単位C5H8が繰り返し結合したもの)
が結合しているため、膜全体が1枚の脂質分子で構成されることになる。
古細菌の脂肪酸は直鎖ではなく、分岐鎖を持つことが多い。
通常の脂肪酸の代わりに、イソプレノイド鎖を持つことがある。
これにより、長い炭化水素鎖が形成され、膜脂質として機能し膜の安定性が増す。
一部の古細菌では、脂質二重層ではなく、脂質単層を形成することがある。
これは極限環境での生存に有利である。
その特殊な細胞膜構造は、エーテル結合【R-O-R’】を持つ脂質二重層から成り、極限環境に対する耐性を持っている。
その遺伝的多様性は、古細菌が真核生物と共通する遺伝子も持ち、進化の過程で独自の枝を形成していることを示す。
ジメチルエーテル:化学式 CH3OCH3
ジエチルエーテル:化学式 C2H5OC2H5
エーテルは有機化学で溶媒として広く使われる。ジエチルエーテルは、抽出や反応の媒体として一般的、
一部のエーテルは麻酔薬としても使われている。)
門 脊椎動物、節足動物など100種類ほど
綱 哺乳綱、両生網、爬虫網 生物→動物から、さらに動物の中を分類
目 霊長目(サル目)、有鱗目(爬虫類の一種であり、トカゲ、ヘビ、カメレオンなどが含まれる)など
科 ヒト科 イネ科、ウマ科、ウシ科など
人間と一緒に暮らすウマは現在、約600種類にのぼる。現代のウマには、たくさんの「血統」が存在すると思っている人もいるかもしれないが、ケンタッキーダービーに出走したサラブレッド種のオス20頭のDNAを調べたなら、その大半が、1700年に生まれた1頭の種馬まで遡るY染色体を持っていることがわかった。その種馬はダーレーアラビアンという名前で、現存するオスのサラブレッドの95パーセントはこの個体とつながっている。
過去最大規模のゲノム時系列の研究によって、人間による極端な品種改良の結果、家畜のウマの遺伝的多様性が極めて低くなっていることが明らかになった。しかも過去1,000年の間に、すべての家畜ウマのY染色体の多様性が低下していた。
こうように、遺伝子の多様性が低いと、遺伝的な病気になりやすい。ウマにはすでに、夜盲症や筋疾患などの障害が現れている。更新世チバニアン期の70万年以上前のウマの脚の骨から、これまでで最も古いゲノム(全遺伝情報)が解読された。既に、更新世後期の7万~8万年前という古代の化石のゲノムでも解読が可能になっていた。その更新世のウマのゲノムの解読を通して、現生のウマ・シマウマ・ロバを含むウマ属の系統の祖先が今から新生代新第三紀鮮新世の400万~450万年前に出現していたことを突き止められた。
さらに、古代のウマのゲノムからは、現生の家畜ウマと、絶滅の危機に瀕している野生のプシバルスキーウマPrzewalski's Wild Horse (モウコノウマ)との進化上の関係も明らかになった。プシバルスキーウマはモンゴルのステップ地帯原産で、現存する最後の野生ウマだ。中央アジアの草原にかつて広く分布した野生ウマで、一部の家畜ウマの祖先型でもある。19世紀以降生息地の破壊と狩猟によって急激に姿を消し、現在ではモンゴルのタンシ・シャラ・ヌル山脈にわずかの群れが生き残るのみとなった。通常、10頭以下の小集団で生活するといわれる。日中は砂漠で過ごし、日が暮れると草地に現れて、草を食べ水を飲む。近年では絶滅寸前と思われ、1~2頭を見たという目撃例がわずかにあるにすぎない。1926年以来モンゴル人民共和国政府によって保護されてきたが、成果が見られない。
プシバルスキーウマ(蒙古野馬;学名 Equus ferus przewalskii)の染色体数は66本で、家畜ウマの祖先の共通の系統から分岐した種であることがわかった。現生の家畜馬のウマも蒙古野馬の別の一亜種である。その染色体数は64本。2つのグループが分岐したのは更新世後期の約5万年前のことだった。
プシバルスキーウマは、一度は野生下では絶滅したと見られ、わずか数十頭の飼育個体から再野生化が試みられた。これほど少ない個体数では、遺伝的多様性が小さすぎて種の保存は困難であるが、プシバルスキーウマはアラブやアイスランドホースといった家畜ウマの品種よりも遺伝的多様性が大きいことが今回の研究で明らかになった。
プシバルスキーウマは、種の保存活動を行うに足る十分な遺伝的多様性を備えていることが分かった。プシバルスキーウマは、ノウマとしては唯一の野生馬と考えられていた。それが近年の研究では、1968年頃に野生下では一度絶滅したと見られている。加えて現生のプシバルスキーウマはその家畜馬が古い時代に再野生化した子孫であるとの説が唱えられていたが、2021年の再調査では、プシバルスキーウマは家畜化されたことはないという結論に達した。現在は飼育されていたプシバルスキーウマの子孫の一部が、辛うじて再び野生化している。
旧石器時代約5,500年前の北カザフスタンの新石器時代のボタイ遺跡の発掘調査により、最初期の家畜馬の痕跡が確認された。その遺跡の馬はプシバルスキーウマの系統であることが分かった。したがってプシバルスキーウマから分岐した現生の家畜馬の系統ではない。
北カザフスタンの新石器時代のボタイ遺跡の発掘調査によれば、カザフスタンは馬の飼育が始まった地域でもある。住居や多数の石器、象牙製品が発見された。およそ4,000年前、青銅器時代には、カザフスタンの地域にはアンドロン文化とベガズィ・ダンディバイ文化と呼ばれる部族が住んでいた。彼らは農耕や牧畜を行い、戦車を巧みに操る戦士でもあった。現在でも、古代の人々が部族の神殿や聖域として利用した岩の上には、戦車の絵が描かれているのを見ることができる。太陽に焼かれた黒い岩肌には、踊りの場面や太陽の頭を持つ神々の姿、強大なラクダや雄牛など、古代の神々を象徴する彫刻が刻まれている。彼らこそが、現在も続いている銅の採掘を始めた人々でもあった。今日のジェズカズガンやサヤクの銅採掘場がその例である。この時代の人々は優れた戦士であり、羊飼いや農夫でもあり、同時に熟練した金属加工技術者でもった。青銅を用いて、斧・ナイフ・短剣、さらには様々な装飾品を製作していた。
ウマは一頭前後のオスが中心となり、全ての雌を率いるハーレムを構成する。雌には序列があり、年長のメスの序列が一般的には高い、という小規模の群れで暮らす。雄は群れを率いて、他の雌が独身の雄に奪われないように群れを守っている。若い雄がリーダー雄を倒して、ハーレムを奪う。ウマは草原の草生草や牧草などを含む粗飼料を選択しないで採食する典型的なグレイザーgrazerである。
1966年にハンガリーの昆虫学者によって目撃されたのを最後に野生下でウマの目撃情報が確認されなくなり、恐らく1968年頃、野生下では一度絶滅したと見られている。かつて発見された多くの個体が欧米諸国の動物園に送られており、その子孫が生き残っていたことから、飼育下での計画的な繁殖が始められ、再野生化が試みられた。現在は、世界各地の動物園で1,000頭以上が飼育されている。モンゴルのフスタイ=ヌルー保護区で再野生化が行われ100頭以上に回復している。また、新疆ウイグル自治区の自然保護区などでは、再野生化の目的で飼育個体の一部が保護区に移されている。
属 ヒト属、イネ属、ウマ属、ロバ属、シマウマ属、ノロバ属、イヌ属など
種 ホモ・サピエンス 200万種類以上のものがある。
生物の 科 属 種 の分類は、主に以下の基準に基づいて行われる。
形態学的特徴
科は、複数の属をまとめたもので、共通する形態や構造的特徴に基づいて分類されます。例えば、犬科の動物(犬・キツネ・オオカミなど)は共通の特徴を持つ。
属は、同じ科に属する複数の種を含むグループで、より具体的な形態的特徴や遺伝的特徴に基づいて分類される。
種は、もっとも具体的な分類単位で、交配して繁殖可能な個体群を指す。
遺伝的特徴
分子生物学的手法(DNAシークエンシングなど)を用いて、遺伝子レベルでの相同性や差異を解析し、分類を行う。
DNAシークエンシング(DNA sequencing)は、生物のDNAの塩基配列(A、T、C、G)の正確な順序を決定する技術で、この手法は、生物の遺伝情報を理解し、遺伝子の機能や進化の歴史を解明するのには不可欠である。
生理学的特徴
生物の生理的なプロセス(例: 酵素の働き、代謝経路など)に基づいて分類を行う。
生態的特徴
生物がどのような環境に住み、どのような生態的役割を果たしているかに基づいて分類される。
これらの基準を総合的に利用することで、生物は科学的かつ体系的に分類される。
生物は地球上で最も多様で複雑な存在であり、その理解には多くの科学的なアプローチが必要となる。
進化は、生命がどのようにして現在の形になったのか、そしてどのようにして新しい種が生まれるのかを探る科学である。
生態系では、 生物がどのようにして互いに、そして環境と相互作用するのかを理解する。これには食物連鎖、環境の変化への適応、そして生物多様性の保全が含まれる。
遺伝学では、遺伝子の構造と機能、生物がどのように遺伝情報を次世代に伝えるのかを研究する。
生理学では、生物の体内で起こる化学反応やプロセスを探る分野で、どのようにしてエネルギーを生成し、使用するのか、どのようにして成長し、修復するのかを理解する。
解剖学では、生物の構造、特にその内部構造を詳細に調べる。これには、解剖によって組織や器官を研究することも含む。
これらの基準を総合的に利用することで、生物は科学的かつ体系的に分類される。
目次へ
生物学における「ドメインdomain」は、生命の分類体系の中で最も広範な分類カテゴリーの一つ。
ドメインは、全ての生命を大きく三つの主要なグループ、バクテリア・アーキア・真核生物に分けるために使用される。
バクテリアBacteria(真細菌) : 核膜を持たない単細胞の原核生物で、多くの異なる環境で生息している。
アーキアArchaea(古細菌) : 核膜を持たない単細胞の原核生物であるが、バクテリアとは遺伝的・生化学的に大きく異なる。一般に極限環境(高温、高塩分など)で生息する微生物群を含む。具体的には以下のような生物がいる。
メタン生成菌Methanogenは、酸素が存在しない環境でメタンを生成するアーキア。主に湿地や下水処理施設、そして動物の消化器官内に生息している。動物の消化器官内に生息するメタン生成菌の代表的なものとしては、メタノバクテリウム属(Methanobacterium)やメタノコックス属(Methanococcus)などが挙げられる。これらの菌は、動物の腸内で有機物を分解し、メタンガスを生成する役割を果たす。
好熱菌 : 高温環境で生存できるアーキア。例えば、熱水噴出孔や温泉などで見られる。「ピロリドス」と特定される好熱菌は存在しないようであるが、一般的に好熱菌は高温環境で生息し、特殊な酵素や代謝経路を持っている。これらの細菌は、熱水噴出孔・温泉・工業的な高温プロセスなど、極限的な温度条件で成長する。
代表的な好熱菌の一つサーマス・アクアティカスThermus aquaticusはその高い好熱性から産生するタンパク質の耐久性も高く、多くの酵素類が実用化されている。DNA を鋳型としてDNA を合成する DNA 依存性 DNA ポリメラーゼと、RNA を鋳型として DNA を合成する RNA 依存性 DNA ポリメラーゼの、2つのタイプがあるが、前者はDNA複製やDNA修復において中核的な役割を担う酵素である。この技術は、医学・法科学・遺伝子研究など多岐にわたる分野で利用されている。
好塩菌 : 高濃度の塩分環境で生息するアーキア。塩湖や塩田などに生息し、ハロバクテリアHalobacteriumなどがある。ハロバクテリアのプロテインproteinや酵素は、塩耐性を持つため、塩分の多い環境でも安定して機能する。この特性を利用して、食品加工や医薬品の製造に応用されることがある。またハロバクテリアは有毒な廃棄物の分解に利用される。特に、高塩濃度の廃水処理において、その耐塩性が役つ。ハロバクテリアは光合成を行う能力を持つ種もあり、バイオエネルギーの生産に利用される可能性がある。例えば、バクテリオロドプシンという光反応タンパク質は、光を利用してエネルギーを生成することができる。
好酸菌 : 強い酸性環境で生存できるアーキア。鉱山の排水や酸性温泉などに見られる。
アーキアは、通常の環境にも存在するが、その特殊な生態や極限環境への適応が特に興味深い。アーキアの研究は、生命の限界や生物の多様性を理解する上で非常に重要である。
アーキア(古細菌)とバクテリア(真細菌)はどちらも原核生物であり、核膜を持たない細胞を持っているが、遺伝的および生化学的にはいくつかの重要な違い。
遺伝的な違い
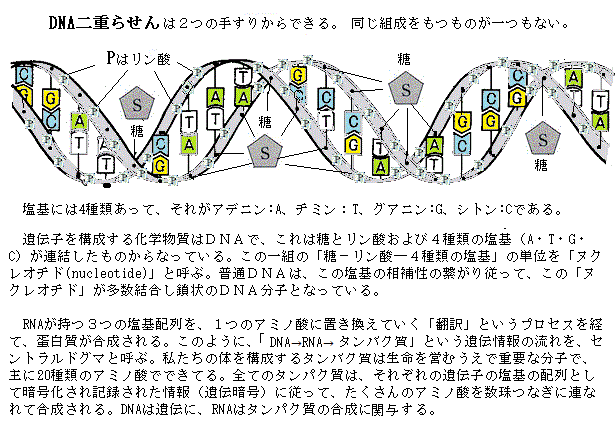 |
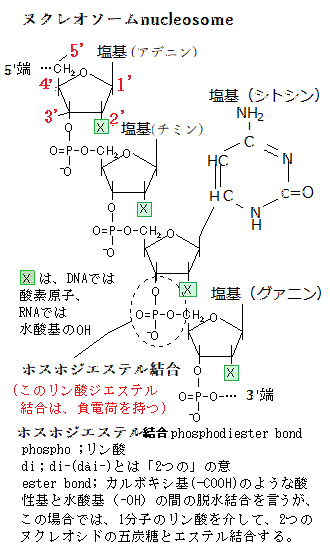
| DNA(デオキシリボ核酸)とRNA(リボ核酸)は、どちらも遺伝情報の管理と伝達に重要な役割を果たすが、いくつかの違いがある。 DNA構造は二重らせん構造を持っている。これは二本のヌクレオチド鎖がらせん状に巻きついた構造である。 DNA役割は遺伝情報を長期的に保存し、その情報を次世代に伝える役割がある。細胞の核に存在し、全ての遺伝子を含んでいる。 DNAの塩基はアデニン(A)、チミン(T)、シトシン(C)、グアニン(G)で構成されている。 RNA構造:は通常、一本のヌクレオチド鎖から構成されている。二重らせん構造ではないが、一部のRNA分子は二次構造をとることがある。rRNA(リボソームRNA)の二次構造の特定の役割は、リボソームの主要な構成要素の一つとなり、リボソームの構造形成に重要な役割を果たし、タンパク質合成の場として機能する。mRNA(messengerメッセンジャーリボ核酸)の翻訳過程で重要な役割を果たす。rRNA二次構造は多数のヘアピンループhairpin loopやステムループ構造を持ち、これがリボソームの複雑な三次構造の基盤を形成する。ヘアピンループ構造は、rRNAや他のRNA分子でよく見られる二次構造の一つで、一本のRNA鎖が折り返す茎の部分(ステムstem)は、自己相補的な配列を形成することにより、ヘアピンのような形状になる部分を示す。折り返し部分loopは非相補的な塩基配列であり、その前後の茎の部分(ステムstem)は相補的な塩基対が形成されるため、その部分は二重らせん構造を取る。 ステムループ構造stem loopでは、ステム(茎部分)は相補的な塩基対が形成される領域であり、ループ部分は非相補的な塩基配列が折り返して形成される領域であり、この構造はRNAの折りたたみや機能において重要な役割を果たす。 リボソームはタンパク質合成の場であり、rRNAとリボソームタンパク質が組み合わさって構成されている。リボソームには大サブユニットと小サブユニットの2つの部分があり、それぞれにrRNAが含まれている。大サブユニット(50Sサブユニット)はタンパク質を形成するアミノ酸を結合する反応を行う。大サブユニットには、5S rRNAと23S rRNAが含まれている。小サブユニット(30Sサブユニット)は、mRNAの読み取りと、適切なtRNA(転移RNA):が結合する場を提供する。小サブユニットには、16S rRNAが含まれている。 rRNAはリボソームの構造を安定化し、リボソームの機能を助ける役割を果たしている。例えば、rRNAはmRNAとtRNA(transfer転移RNA)の正しい位置合わせを行うことで、正確なタンパク質合成をサポートする。tRNA:の役割は、タンパク質合成の過程で、アミノ酸をリボソームに運搬し、mRNAのコドンcodon(mRNA上の3つの連続したヌクレオチド)に対応するアンチコドンanticodonを提供する。これにより、正しいアミノ酸がポリペプチド鎖(タンパク質)に結合される。アンチコドンはtRNA分子の一部で、mRNAのコドンと相補的な3つのヌクレオチドから構成されている。タンパク質合成の過程では、tRNAは特定のアミノ酸を運んできて、リボソームでmRNAのコドンと結合する。この結合は、アンチコドンとmRNAのコドンの相補的な関係に基づいて行われる。例えば、mRNAのコドンがAUGの場合、tRNAのアンチコドンはUACとなり、これにより、tRNAがメチオニンというアミノ酸を運んで来る。mRNAのコドンがUUUの場合、tRNAのアンチコドンはAAAとなり、フェニルアラニンというアミノ酸を運んで来る。このプロセスは、タンパク質の正確な合成に重要であり、遺伝情報が正確に翻訳されるために必要となる。 コドンcodonは、mRNA上の3つの連続したヌクレオチドが一組になって、特定のアミノ酸を指定するためのコードを持っている。mRNAはタンパク質合成の設計図として機能し、リボソームで読み取られ、tRNAが対応するアミノ酸を運んでくる役割を果たす。 mRNAのコドンの例としては • AUG:メチオニン(開始コドンとしても知られる) • UUU:フェニルアラニン • GGC:グリシン 開始コドンはタンパク質合成の開始を示し、終止コドン(UAA、UAG、UGA)はタンパク質合成の終了を示す。 tRNAは、タンパク質合成において重要な役割を果たすRNA分子で、その独特な二次構造がクローバーリーフ形状に似ていることから、クローバーリーフ構造と呼ばれ、特定の三次構造(L字型)を形成して機能する。これにより、アミノ酸とアンチコドンの正確な配置が可能になる。RNAの二次構造は、その機能を実現するために不可欠で、二次構造の形成によって、RNA分子は適切な立体配置を取ることができ、特定の化学反応や相互作用を効率的に行うことができる。この構造は以下のように4つの主要な腕(ステムループ)と1つのアミノ酸結合部位から成り立っている。 Dアーム(D loop)の部分にはジヒドウリジン(Dihydrouridine)という特殊な塩基が含まれている。Dアームは構造の安定化と折りたたみに関与する。 アンチコドンアーム(Anticodon arm)のアームには、アンチコドンがあり、mRNAのコドンと相補的に結合する。これにより正しいアミノ酸がリボソームへ運ばれる。 TΨCアーム(T armまたはT loop)の部分には、シュードウリジンPseudouridineという特殊な塩基が含まれている。TΨCアームはリボソームとの結合に重要な役割を果たす。 アミノアシル結合部位(Acceptor stem)のアームの3'末端にはCCA配列があり、ここにアミノ酸が結合する。アミノアシルtRNAシンテターゼという酵素が、この部分にアミノ酸を結合させる。 可変ループVariable loopの一部のtRNAには、TΨCアームとアンチコドンアームの間に変動する長さのループが存在する。この可変ループはtRNAの種類によって異なる。 このクローバーリーフ構造は、tRNAが適切に機能し、mRNAの情報に基づいて正確にアミノ酸をリボソームに運ぶために必要となる。 |
アーキアとバクテリアは両方とも原核生物ですが、遺伝的には大きな違いがある。特にrRNAの配列においてその違いが顕著で、rRNAはリボソームの一部を構成するRNAで、タンパク質合成の場として重要な役割を果たす。
The DNA molecule is shaped like a double helix. (DNA分子は二重らせんの形状をしている)
塩基が形成するヘリックスhelixとは、DNAやRNA分子の構造において、塩基が特定のパターンで結合し、らせんhelix(ヘリックス)構造を形成することを指す。
DNAの二重らせん構造 : DNAは二重らせん構造を持っており、2本のヌクレオチド鎖が互いに巻きついている。塩基はそれぞれのヌクレオチドに結合しており、A(アデニン)とT(チミン)、C(シトシン)とG(グアニン)が特異的にペアを形成して、内側で水素結合を作り出す。これにより、二重らせん構造が安定する。
RNAの二次構造 : RNAは通常一本鎖であるが、内部で折りたたまれ、ヘリックスやループといった二次構造を作る。例えば、tRNA(転移RNA)には、このような二次構造が見られる。RNAの塩基も特異的にペアを作り、A(アデニン)とU(ウラシル)、C(シトシン)とG(グアニン)が結合する。
このヘリックス構造は、分子の安定性を高めるだけでなく、DNAやRNAの機能においても重要な役割を果たす。具体的には、遺伝情報の保持や伝達、タンパク質合成など、生命活動の基本的なプロセスに関与している。
具体的に言うと、アーキアとバクテリアのrRNA配列の違いには特徴がある。
配列の異なるセグメントがある。rRNAには16S rRNA(小サブユニット)や23S rRNA(大サブユニット)といった異なるセグメントがあるが、これらのセグメントの塩基配列がアーキアとバクテリアで異なる。この違いは、特定の位置における塩基の置換(例えば、アデニンがグアニンに置き換わるなど)や挿入・欠失によるものである。
(DNAのセグメントとは、DNAの特定の部分や区分のことを指す。DNAは長い分子鎖で構成されており、その中には特定の機能や役割を持つ部分がある。例えば、遺伝子【タンパク質をコードする部分】やプロモーター領域【遺伝子の発現を制御する部分】などがDNAのセグメントとして挙げられる。
リボソームの小サブユニットと大サブユニットで説明すると、リボソームはタンパク質合成の場であり、2つの主要な部分から構成されている。
小サブユニット(30Sサブユニット) : 原核生物では30Sサブユニットとして知られている。 主に16S rRNAを含む。 mRNAの結合部位として機能し、mRNAの翻訳開始を助ける。
大サブユニット(50Sサブユニット) : 原核生物では50Sサブユニットとして知られている。 主に23S rRNAと5S rRNAを含む。 タンパク質合成中にアミノ酸を結合させる役割を果たす。 リボソームがタンパク質を合成する際、小サブユニットがまずmRNAに結合し、その後大サブユニットが結合して機能的なリボソームが形成される。このリボソーム全体が、mRNAの情報を基にアミノ酸を連結してポリペプチド鎖【タンパク質】を生成する。
実際には「30Sサブユニット」という名称は「30個のサブユニットがある」という意味ではない。ここでの「30S」は、リボソームの質量やサイズに関係する単位である「スヴェドベリ単位(S単位)」を指す。
スヴェドベリ単位(S単位)とは、分子や粒子の沈降速度を測定するための単位で、遠心分離機によって測定される。数値が大きいほど、粒子の沈降速度が速く、一般的にサイズや質量が大きいことを示す。
リボソームのサブユニット 原核生物のリボソームは、以下のように構成されている。
小サブユニット(30Sサブユニット) : 16S rRNAと21種類のタンパク質を含む。 主にmRNAの結合部位として機能する。
大サブユニット(50Sサブユニット) : 23S rRNA、5S rRNAと34種類のタンパク質を含む。 アミノ酸を結合させる役割を果たす。
これらのサブユニットが組み合わさって、完全なリボソーム【70Sリボソーム】)を形成する。70Sという数値も同様にスヴェドベリ単位を示しており、リボソーム全体の沈降速度に基づいている。
つまり、「30Sサブユニット」という名称は、そのサブユニットの特性を示す科学的な指標であり、実際の数とは無関係である。 )
 二次構造の違い : rRNAの二次構造、すなわち塩基が形成するヘリックスやループの形状が異なる。この構造の違いは、リボソームの全体的な構造や機能に影響を与える。
二次構造の違い : rRNAの二次構造、すなわち塩基が形成するヘリックスやループの形状が異なる。この構造の違いは、リボソームの全体的な構造や機能に影響を与える。 保存された領域と変異領域 : 一部のrRNA配列は全ての生物種で保存されており(保存された領域)、他の部分は変異が見られる(変異領域)。アーキアとバクテリアでは、この変異領域が異なるため、遺伝的に明確に区別される。
分子時計の役割 : rRNAの変異速度は一定であることが多いため、進化的な距離を測る「分子時計」としても使用される。このため、アーキアとバクテリアのrRNA配列の差異は、両者が異なる進化の道を歩んできたことを示していることになる。
これらの違いにより、科学者はアーキアとバクテリアを遺伝的に明確に区別することができる。特に16S rRNA配列の比較は、微生物の分類や同定において非常に有用である。
アーキアのrRNA配列はバクテリアのものとは大きく異なり、これが遺伝的に両者を明確に区別する要因となっている。 具体的には、以下の点が挙げられる。
進化的関係 : rRNA配列の違いは、アーキアとバクテリアが異なる進化の系統に属していることを示している。これは、両者が共通の祖先から分岐した後に、それぞれ独自の進化を遂げたことを意味する。
機能的特性 : rRNA配列の違いは、リボソームの構造や機能に影響を与える可能性がある。この違いにより、アーキアとバクテリアは異なる環境や条件下で生存する能力を持つことができる。
分類学的区分 : rRNA配列の比較に基づいて、アーキアとバクテリアはそれぞれ独自の分類学的カテゴリーに置かれている。これにより、遺伝学的に明確に区別されることが科学的に確認されている。
このように、rRNA配列の違いは、アーキアとバクテリアを遺伝的に区別する重要な基準となっている。
遺伝子構造と発現
アーキアの遺伝子構造や発現機構は、真核生物に似ている。例えば、アーキアはヒストンのようなタンパク質を使ってDNAをパッケージングしするが、バクテリアにはこれがない。
生化学的な違い
有機基functional groupは、有機分子内で特定の化学反応を引き起こす特性を持つ原子団や結合のグループを指す。その有機基が、分子の物理的および化学的性質に大きな影響を与える。
例として、ヒドロキシル基(-OH)、カルボキシル基(-COOH)、アミノ基(-NH2)などがある。有機基は、分子の一部であり、他の原子や分子と結合することで異なる化学特性を生み出す。
炭化水素基hydrocarbon groupは、炭素と水素からなる有機基の一種で、分子内の炭素骨格に含まれる。炭化水素基は、炭素-炭素および炭素-水素結合から成り、アルカン(-CH3、-CH2-)、アルケン(-CH=CH-)、アルキン(-C≡C-)などの種類がある。 これらの基は、炭素と水素のみを含むため、基本的には疎水性(親油性)を示す。炭化水素基は、エネルギー源としての用途が最も一般的で、ガソリン・ディーゼル・ジェット燃料など自動車や航空機などの燃料として使用されている。また、ポリエチレンやポリプロピレンとして、包装材・日用品・玩具・医療機器など、様々な製品に使用されるプラスチックの原料として使用されている。このように有機基は多様な化学特性を持ち、分子の機能に影響を与えるのに対し、炭化水素基は特に炭素と水素からなる基であるため、その性質や応用が異なる。
アーキアの細胞膜は、エーテル結合Ether Bondを持つリン脂質で構成されており、極限環境への適応に寄与している。バクテリアの細胞膜はエステル結合Ester Bondのリン脂質でできている。
エーテルは、2つの炭素原子が1つの酸素原子を介して結合している構造を持つ。一般式は R-O-R'(R と R' は炭化水素基)。例えば、ジメチルエーテル (CH₃-O-CH₃) やジエチルエーテル (C₂H₅-O-C₂H₅) など。エーテル結合は比較的安定で、極性が低く、水に溶けにくい性質がある。また、エーテルは揮発性が高く、多くの有機溶媒として使用される。
エステルは、酸とアルコールが縮合反応して形成される構造を持つ。一般式は R-COO-R'(R と R' は炭化水素基)。例えば、酢酸エチル (CH₃-COO-C₂H₅;エチルの化学式C₂H₅-の官能基を持つ) や脂肪酸エステル(R-COO-R':R は脂肪酸の炭化水素鎖・R' はアルコール由来の炭化水素鎖、脂肪酸とアルコールに依存し、基本的な構造はエステル結合)など。エステル結合は分極しており、香りや風味のもととなる揮発性物質や生体分子、例えば脂肪酸エステルに多く見られる。水と反応して酸とアルコールに分解する加水分解反応を起こす性質もある。
これらの化学結合は、有機化学および生化学の中で重要な役割を果たしており、それぞれの特徴を理解することで、化合物の性質や反応性をより深く理解することで実用化の分野が広がる。
細胞壁の組成は、バクテリアの細胞壁はペプチドグリカン(ムレイン)という物質で構成されているが、多くのアーキアはペプチドグリカンを持たないか、異なる成分で細胞壁が作られている。
(ムレインは、多くの細菌の細胞壁を構成する主要な成分で、これによって細菌は外部環境からの圧力に耐え、形状と剛性を保つことができる。
ムレインの構造は糖鎖N-アセチルグルコサミン(NAG)とN-アセチルムラミン酸(NAM)の糖分子が交互に連結して長い鎖を形成する。
NAMの分子に結合した短いアミノ酸鎖、これらのペプチド鎖がクロスリンク(交差結合)して糖鎖を網の目のように連結し、強固な三次元構造を形成する。)
代謝経路では、アーキアは独自の代謝経路を持つことが、メタン生成などの特殊な代謝プロセスを行う。バクテリアではこのような代謝は見られない。
これらの違いにより、アーキアとバクテリアは生物学的に大きく区別され、異なる進化の道を歩んできたと考えられている。
真核生物Eukaryaは、真核細胞でできているが、原核生物は原核細胞でできている。真核細胞はDNAが核という膜で包まれているが、原核細胞はDNAが細胞内でむき出しになっている。真核細胞は細胞小器官という特化した器官をもつが、原核細胞は細胞小器官をもたない。核膜を持つ細胞を持つ生物で、動物・植物・菌類、そして原生生物Protistなど多様な生命を含む。
原生生物は、真核生物の中で動物・植物・菌類に分類されない単細胞の微生物を主に指す。これらは多様な形態と生態を持つグループで、非常にさまざまな種類がある。
原生生物の定義では、以下の特徴を持つ真核生物のグループを示す。原生生物は単細胞であるが、一部は多細胞であるか、群体colonyを形成する。自養性(光合成を行う)や他養性(他の生物を捕食したり有機物を分解する)など、多様な生活様式がある。原生生物は多様なグループに分類される。
アメーバAmoeba proteusは、偽足(擬似的な足)を使って移動し、捕食を行う。繊毛虫Ciliophoraのゾウリムシは、細かい繊毛を使って移動し、餌を取り入れる。鞭毛虫Euglenozoaのユーグレナは、鞭毛を使って移動し、一部は光合成を行う。緑藻ChlorophytaのクロレラChlorellaは、光合成を行う単細胞植物で、淡水や海水に生息する。アピコンプレクサApicomplexaのマラリア原虫Plasmodiumは、宿主の細胞内に寄生し、複雑な生活環を持つ。ディアトムDiatomsは、シリカ(ケイ素)でできた細胞壁を持ち、主に海洋および淡水環境で光合成を行う。
原生生物は、生態系において重要な役割を果たし、食物連鎖の基礎を支えたり、病原体として人間や他の動物に影響を与えたりする。
これらのドメインは、遺伝子の解析などの現代の科学技術を用いて、生物の系統関係(進化の歴史)をより正確に反映するために定義されている。
原核生物Prokaryotesと原生生物Protistsは、異なる生物分類群に属しており、それぞれの特徴や生態も大きく異なる。
核膜の有無 : 原核生物は核膜を持たないが、原生生物は持つ。
細胞小器官 : 原核生物は膜結合型細胞小器官を持たないが、原生生物は持つ。
細胞の多様性 : 原核生物は主に単細胞生物であるが、原生生物は単細胞から多細胞まで多様である。
細胞壁 : 原核生物はムレインを含む細胞壁を持つが、原生生物の細胞壁の成分は様々。
上記ように、原核生物と原生生物はそれぞれの構造や機能において大きく異なる。
原核生物の細胞構造 : 原核生物は核膜を持たない単細胞生物で、遺伝物質DNAは細胞質内に存在するが環状のDNA分子であることが多い。原核生物は基本的に膜結合型の細胞小器官を持っていない。代わりに、リボソームなどの非膜結合型の構造が見られる。ミトコンドリアや葉緑体は、エネルギーの生成や光合成を行う重要な細胞小器官であるが、これらは真核生物に特有のものである。興味深いことに、ミトコンドリアと葉緑体は、進化的にはかつての原核生物が、他の細胞と共生する形で取り込み真核生物として進化した(共生説)。
原核生物のリボソームは、細胞質内を自由に浮遊する形で存在している。これを「遊離リボソーム」と呼ぶ。これらのリボソームは、主に細胞内で使用されるタンパク質の合成に関与する。具体的には、mRNA(メッセンジャーRNA)からアミノ酸を結びつけてタンパク質を生成するプロセス、つまり「翻訳の場」として機能する。
mRNAワクチンは、ウイルスの表面にある「スパイクたんぱく質」と呼ばれるたんぱく質の遺伝情報を含んだ「mRNA」をヒトの体内に投与する。 この「mRNA」はヒトの細胞の中で設計図のように働いて次々とスパイクたんぱく質が作られる。すると免疫の働きでこのスパイクたんぱく質に対する抗体が多く作られるようになり、実際にウイルスが入ってきてもすぐに攻撃できるようになる。 ただ、mRNAは不安定で、ワクチンとして投与した場合も、すぐに分解されてしまうため体内には残らない。また、細胞の中に入ってもヒトの遺伝子がある「核」という部分には入り込むことがないため安全性が高い。
遊離リボソームの他にも、原核生物にはさまざまな遊離分子が役割を担っている。
• tRNA(転移RNA) : アミノ酸をリボソームへ運び、タンパク質合成の際に重要な役割を果たす。
• mRNA(メッセンジャーRNA) : DNAからの遺伝情報をリボソームに伝えるメッセンジャーの役割を持つ。
• 酵素 : 様々な生化学反応を促進するために自由に浮遊する。
これらの分子が協力して、原核細胞の生命活動を支えている。
種類 : 原核生物には細菌(バクテリア)と古細菌(アーキア)が含まれる。
細胞壁 : 多くの原核生物はムレイン(ペプチドグリカン)を含む細胞壁を持つ。これは細胞の形状と剛性を維持するのに役立つ。
分裂方式 : 二分裂によって増殖する。
遺伝情報の交換 : 一部の原核生物は形質転換、接合、トランスダクションなどの方法で遺伝情報を交換する。
原生生物の細胞構造 : 原生生物は主に単細胞生物であるが、一部は多細胞生物も含まれる。真核生物であり、核膜を持ち、ミトコンドリアや葉緑体などの膜結合型細胞小器官を持つ。
種類 : 原生生物には、原生動物・藻類・粘菌が含まれる。これらは非常に多様な生物群である。
細胞壁 : 一部の原生生物、特に藻類は細胞壁を持つが、成分はセルロースやシリカなど異なる。
分裂方式: 二分裂、出芽、胞子形成など様々な方法で増殖します。 生態系の役割: 一部の原生生物は光合成を行い、一次生産者として働くものもいます。他のものは分解者や捕食者として生態系に貢献します。
目次へ