高等植物の光合成
TOP 車山お知らせ 車山ブログ 歴史散歩 車山高原野鳥 たびネット信州
| 目次 |  |
| 1)光合成由来の生命エネルギー | |
| 2) 光合成器官 | |
| 3) 反応中心 | |
| 4)電子伝達系とプロトン駆動力の生成 | |
| 5)PSIの電子移動の精巧さと効率性 | |
| 6)光化学系 | |
| 7)PSII とPSIIの相互関係 |
地球上のすべての生物は太陽の光エネルギーに依存し、その生存に不可欠なエネルギーを獲得する唯一の方法が光合成photosynthesisである。しかも地球上のエネルギー源のほとんどは、原始から現在に繋がる光合成により生産された化石燃料である。
Photoは「光」、Syntheisは「合成」という意味であれば、その本質は、植物が太陽からの光エネルギーを吸収し、化学エネルギーに変えるプロセスを言う。植物という光合成生物は、太陽光エネルギーを利用して二酸化炭素CO2と水H2Oから有機物C6H12O6(糖など)と酸素O2を生成する。これこそが植物が成長するための基本的なエネルギー変換プロセスである。
具体的には、太陽光エネルギーにより二酸化酸素と水から、酸素を放出しながら炭水化物を合成する。炭水化物は、炭素C、水素H、酸素Oから構成される化合物で、主にエネルギー源として利用される。
6CO2 + 6H2O → C6H12O6 + 6O2
化学式 C6H12O6は、日本語で「グルコース」または「ブドウ糖」と呼ばれる。これは、単糖類(糖類の最小単位)の一種で、エネルギー源として重要な役割を果たす。単糖類として蓄えられたエネルギーは、植物の細胞活動に使われ、あらゆる生物のエネルギー源となる。
その炭水化物が分解されると、糖分が生成され、これがさらに微生物によって分解されると、乳酸や酢酸などの有機酸(酸性を示す有機化合物の総称)が生成される。例えば、果物が甘酸っぱくなるのは、糖分が乳酸や酢酸に変わるためである。また、牛乳が腐ると酸っぱくなるのも同じ理由である。
目次へ
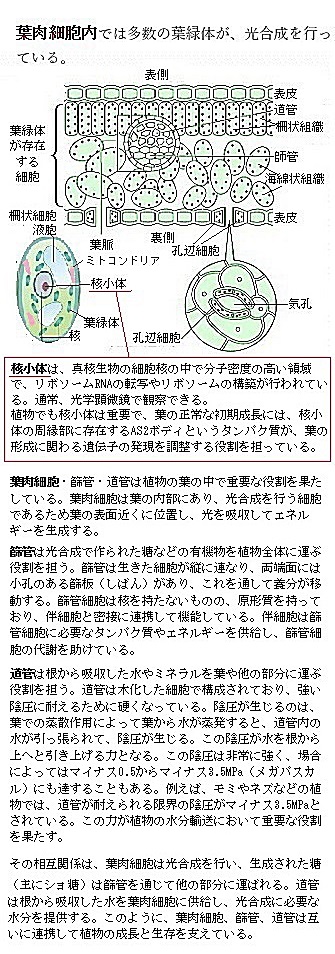 高等植物の最も活発な光合成器官は、葉の葉肉細胞である。その細胞内に多くの葉緑体を持ち、葉緑体自体は2枚の独立した脂質二重膜に包まれ、これらの膜は包膜envelopeと呼ばれる。
高等植物の最も活発な光合成器官は、葉の葉肉細胞である。その細胞内に多くの葉緑体を持ち、葉緑体自体は2枚の独立した脂質二重膜に包まれ、これらの膜は包膜envelopeと呼ばれる。この2枚の膜には、代謝に必要な様々な輸送システムを含む。その葉緑体の中には光を吸収する緑色の色素、葉緑素chlorophyllを多数含む。光合成は植物の葉緑体内で太陽光のエネルギーで水H2Oを酸化し、酸素O2を放出する。同時に二酸化炭素CO2に還元して、ショ糖などの大きな分子の炭水化合物を合成する。そのCO2の還元で終る複雑な一連の光合成反応系は、チラコイド膜上での反応系と、葉肉細胞の溶液中の炭酸固定反応系で行われる。
チラコイド膜Thylakoid membraneは、葉緑体やシアノバクテリアの中に存在する膜構造で、光合成の光化学反応が行われる場所である。チラコイドという名前は、ギリシャ語の「嚢thylakos」に由来する。そのチラコイド膜は、光合成色素やタンパク質複合体が埋め込まれたリン脂質二重層で構成されている。主なタンパク質複合体には、光化学系I・光化学系II・シトクロムb6f複合体・ATP合成酵素などがある。これらの複合体は、光エネルギーを利用して電子を移動させ、最終的にATPとNADPHを生成する。
葉緑体内では、2つの異なる機能単位である光化学系I(PSI)と光化学系II(PSII)があり、これらは植物・藻類・シアノバクテリアのチラコイド膜に存在し、光エネルギーを化学エネルギーに変換する役割を果たす。
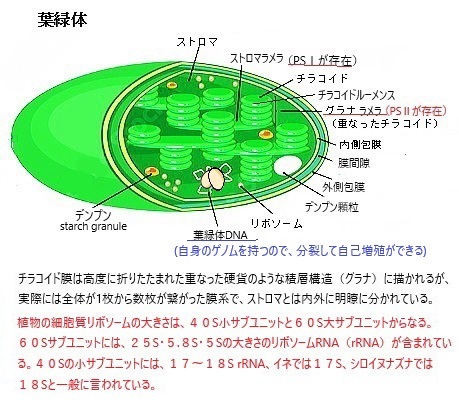
葉緑体の構造の中で最も目立つのが、チラコイドThylakoidと呼ばれる発達した内膜構造である。すべてのクロロフィルはこの膜上にあり、光合成の光反応はここで行われる。チラコイドは、円盤状の構造が積み重なって「グラナ」と呼ばれる構造を形成し、これが葉緑体の中に存在する。グラナの構造により光合成の効率を高める高い表面積対体積比を持つことになる。光合成の過程では、チラコイド膜で水の光分解・電子伝達系・ATP合成などが行われる。これにより、植物は光エネルギーを化学エネルギーに変換し、成長やエネルギーの貯蔵に利用する。
光化学系II(PSII)が、光合成の最初のステップで、光エネルギーを利用して水を分解し、酸素・プロトンH+・電子を生成する。炭素還元反応は、光合成の一部であり、特にカルビン・ベンソン回路で行われるが、この反応はストロマstromaと呼ばれる葉緑体内の、チラコイド外側の水溶液中の酵素で触媒される。チラコイドの大部分は、互いに密着した積層構造で、この膜が積み重なった構造をグラナラメラgrana lamellaと呼ぶ。積層せずにストロマに接してる膜構造をストロマラメラstroma lamellaと呼ぶ。
ストロマは、葉緑体の内部にある液状の部分で、ここで太陽光エネルギーにより二酸化酸素CO2と水H2Oから、酸素O2を放出しながら炭水化物を合成する。
6CO2 + 6H2O → C6H12O6 + 6O2
ストロマで起こるこれらの一連の生化学酸化還元反応は、総称してカルビン回路calvin cycleと呼ばれる。このカルビン回路で二酸化炭素がリブロース-1,5-ビスリン酸(RuBP)と結合し、最終的にグルコースなどの糖が生成される。
特にこの光合成の明反応では、光エネルギーを利用してATPとNADPHが生成される。この反応は葉緑体のチラコイド膜で行われる。光合成に必須な多様なタンパク質が、チラコイド膜に埋め込まれている。多くの場合、これらのタンパク質の一部は、チラコイド膜表面から水溶液中へ突き出している。これらの必須膜タンパク質は、疎水性アミノ酸を多く含むため、膜中の炭化水素部分に固まった疎水領域で安定して存続できる。チラコイド膜上での反応系を要約すると
チラコイド膜上での反応系は、光合成の重要な部分で、その光合成は、葉緑体のチラコイド膜とストロマで行われる一連の反応の2つに分けられる。
チラコイド膜上では、主に次の反応が行われる。光化学系II(PSII)では、光エネルギーを吸収し、水H2Oを分解して酸素O2を放出する。この過程で電子が生成される。生成された電子は、 電子伝達系の一連のタンパク質を通過しながらエネルギーを放出し、そのエネルギーを使ってプロトンH+をチラコイド内に移動させる。
光化学系I(PSⅠ)では、電子が再び光エネルギーを吸収し、NADP⁺をNADPHに還元する。ATP合成酵素は、プロトンH+の濃度勾配を利用してATPを合成する。 これらの反応により、光エネルギーが化学エネルギーに変換され、ATPとNADPHが生成される。これらはストロマでのカルビン・ベンソン回路で使用され、グルコースなどの有機物が合成される。
目次へ
 反応中心、アンテナ色素-タンパク質複合体、そして電子担体タンパク質が必須タンパク質である。その葉緑体の必須膜タンパク質は、各々固有の向きで埋め込まれている。一方はストロマ側に向き、他方はチラコイドルーメンス側に向くなどしている。
反応中心、アンテナ色素-タンパク質複合体、そして電子担体タンパク質が必須タンパク質である。その葉緑体の必須膜タンパク質は、各々固有の向きで埋め込まれている。一方はストロマ側に向き、他方はチラコイドルーメンス側に向くなどしている。ATP(アデノシン三リン酸)の生成は、光化学系II(PSII)で光エネルギーが吸収され、水H2Oが分解されて酸素O2とプロトンH⁺が生成される。電子が電子伝達系を通過する際に、プロトンがチラコイド内腔に移動し、プロトン勾配が形成される。このプロトン勾配を利用して、ATPシンターゼ(ATP合成酵素)がATPを合成する。
ATPシンターゼは、細胞内でエネルギー通貨と呼ばれるATPを生成する重要な酵素である。この酵素は、ミトコンドリアの内膜や葉緑体のチラコイド膜に存在し、プロトンH⁺の濃度勾配を利用してATPを合成する。
ATPシンターゼは、F(_0)モーターとF(_1)モーターという2つの部分から構成されている。F(_0)モーターは膜を貫通するプロトンH⁺の流れによって回転する。この回転が細胞膜の外側に突き出ているF(_1)モーターに伝わり、ATPの合成が行われる。
言い換えれば、プロトンH⁺がF(_0)部分を通過すると、そのエネルギーがF(_1)部分に伝わり、F(_1)部分が回転する。この回転運動が、ADPと無機リン酸(Pi)からATPを合成するためのエネルギーを提供する。 このプロセスが、細胞のエネルギー通貨であるATPを生成するための不可欠な前提となる。
葉緑体の主な共通点といえば外膜と内膜、独自のDNAを持っており、したがって、独自のリボソームや葉緑体DNAから転写されたRNAが存在している。その転写されたRNAは、植物の光合成やその他の重要な機能に関与するタンパク質を合成するための役割を担う。葉緑体には2種類のRNAポリメラーゼが存在する。一つは核にコードされている色素体RNAポリメラーゼnuclear-encoded plastid RNA polymerase(NEP)、もう一つは葉緑体自身にコードされているRNAポリメラーゼplastid-encoded plastid RNApolymerase (PEP)である。これらのRNAポリメラーゼは、葉緑体DNAがそれぞれの異なるプロモーター配列を認識し、特定の遺伝子の転写を開始する。
葉緑体DNAから転写されたRNAは、リボソームで翻訳されてタンパク質を合成する。これらのタンパク質は、光合成装置の構成要素やその他の葉緑体機能に必要な酵素なども含まれる。さらに、葉緑体DNAはリンの貯蔵物質としても機能しており、リン欠乏時にはDNAが分解されて、それに含まれるリンが再利用されることまで解明されている。
葉緑体とは、光合成をおこなう、半自律性の細胞小器官であり、色素体plastidの一種でもある。黄色のカロテノイドもあるが、クロロフィルが多量であるため一般的には緑色に見える。ただし褐藻の葉緑体はクロロフィルのほかにフコキサンチンを持っているため褐色に、紅藻はフィコビリン色素をもっているため紅色に見える。 種子植物など一般的には葉緑体は植物の葉に含まれるが、茎や枝、花弁や果実などの器官でも葉緑体が発達している場合も多い。トマトの果実やヒマワリの花弁の細胞などではカロテノイドを蓄積した有色体が見られる。ナスやキウイフルーツの果実も内側まで薄い緑色になっており、光合成の活性が見られる。
光化学系II(PSII)は、光合成の最初の段階であり、ここで光エネルギーが最初に取り込まれる。PSIIには光捕集複合体があり、ここで光エネルギーが吸収されると、クロロフィル分子が励起される。励起されたクロロフィルはエネルギーの高い電子を放出し、この電子が電子伝達系へと移動する。
PSIIで放出された電子は、水分子から供給される。水分子H2OはPSIIの酸化触媒複合体によって酸化され、酸素O2・プロトンH⁺・および電子を分離する。このプロセスで生成された酸素は大気中に放出され、電子はPSIIで放出された電子の補充として利用される。この一連の反応は「水の酸化反応」と呼ばれ、光合成全体において極めて重要である 。
目次へ
 |
| PSIは、680nm以上の遠赤色光をよく吸収し、PSIIは680nmの赤色光をよく吸収するが、遠赤色光では殆ど作用しない。この波長依存性の違いが、PSIに吸収された遠赤色光は、電子e-を受け取り還元できる強い還元物NADP+と弱い酸化物を作る。 NADP+ → NADPH (弱い酸化物) PSIIは水を分解できる非常に強い酸化能力と、PSIよりも弱い還元物e-を作る。 2H2O → 4e- + 4H+ + O2 この反応スキームの違いが、光合成の電子伝達の理解の基礎になり、その吸収極大波長から、PSIとPSIIの反応中心クロロフィルは、それぞれP700とP680と呼ばれる。 PSIIが作る還元物は、PSIでできた酸化物を再還元することになる。 光合成の過程では、PSIIとPSIが連携して働く。 PSIIは光エネルギーを利用して水H2Oを分解し、酸素、プロトン(H+)、および高エネルギーの電子e-を生成する。この電子がプラストキノン(PQ)を還元してプラストキノール(PQH2)を生成する。その後、PQH2はシトクロムb6f複合体を通じて電子をPSIに運ぶ。 PSIはこの電子を受け取り、さらに光エネルギーを利用して電子をフェレドキシン(Fd)に渡す。最終的に、フェレドキシン-NADP⁺還元酵素(FNR)がNADP+を還元してNADPHを生成する。このように、PSIIで生成された還元物は、PSIで生成された酸化物を再還元する役割を果たす。 |
光化学系IとNADPHの生成
電子伝達系を通過した電子は、ストロマラメラに存在する光化学系I(PSI)に渡される。PSIでも光エネルギーが吸収され、電子が再び高エネルギー状態に励起される。この高エネルギー電子は、フェレドキシンferredoxin (Fd) を介してNADP⁺還元酵素へと伝達され、最終的にNADPHが合成される。フェレドキシン は、内部に鉄-硫黄クラスター (Fe-Sクラスター) を含む鉄硫黄タンパク質の一つであり、電子伝達体として働く。光化学系I(PSI)では還元物質NADPHが生産されるが、このときの電子供与体がフェレドキシンであり、光化学系Iによって励起された電子がこの低い酸化還元電位を持つ電子伝達体に電子を渡し、その電子がフェレドキシン-NADP+レダクターゼFerredoxin-NADP+ reductase (FNR) の触媒により還元され、NADPHが生産される。
このNADPHは、カルビン・ベンソン回路(暗反応)で二酸化炭素を有機物に変換するために必要な還元力を提供する。このため、PSIはNADPH生成における主要な役割を果たし、PSIでの電子移動でも、非常に精巧に効果的に設計されており、これが光合成の全体的な効率を支えている。
PSIは、光合成の過程でも重要な役割を果たし、光エネルギーを化学エネルギーに変換するための一連の反応を行う。PSIの電子移動は、PSIが捕獲した光エネルギーを効率的に電子に変換し、最終的にNADP+をNADPHに還元する。この過程は、非常に高い量子収率を持ち、ほぼ100%の効率で行われている。この精巧な電子伝達系のPSIには、複数の電子伝達成分が含まれており、これらが連携して電子を効率的に移動させる。例えば、P700という特定のクロロフィル分子が光を吸収し、電子を放出する。この電子は、フェレドキシンなどの伝達成分を経て、最終的にNADP+に渡される。
この電子移動の過程では、量子力学的なトンネル効果が利用されており、これにより電子は非常に短い時間で移動する。このため、電子移動の速度が非常に速く、効率的で、そのPSIの電子移動の精巧さと効率性は、光合成の全体的な効率を高めるために不可欠な装置に.なっている。これにより、植物は光エネルギーを最大限に利用し、成長やエネルギー生産に必要な化学エネルギーを生成することができる。
光反応における電子移動の意義
光合成の光反応における電子移動は、単にエネルギーを移動させるだけでなく、エネルギーを蓄積し、利用可能な形に変換する役割を担っている。PSIIとPSI、そしてそれを結びつける電子伝達系は、光エネルギーをATPとNADPHという化学エネルギーに変換するための非常に精巧なシステムである。
このプロセスが正常に機能することで、植物は効率的にエネルギーを獲得し、成長や繁殖のために必要なエネルギーを供給することができる。光反応で生成されたATPとNADPHは、カルビン・ベンソン回路での二酸化炭素固定に利用され、最終的にグルコースなどの有機物が生成される。
目次へ
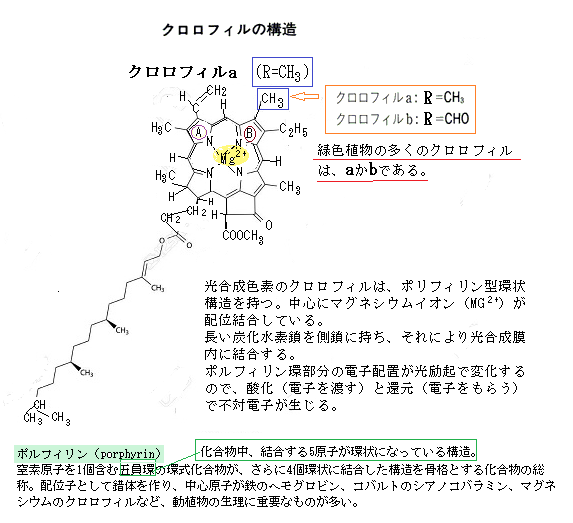 |
| クロロフィルはポルフィリン環と呼ばれる炭素と窒素からなる環状構造に、フィトール鎖と呼ばれる長い炭化水素の鎖がついた形を基本構造とする。 ポルフィリン環の中央にはマグネシウムMG2+が配位している。グネシウムイオンは、クロロフィル分子の中心に位置し、光エネルギーを吸収して化学エネルギーに変換する。 具体的には、クロロフィルは光を吸収し、そのエネルギーを使って水H2Oを分解し、酸素O2を放出する過程で、マグネシウムイオンは光エネルギーを効率的に吸収し、電子e-の移動を助けることで、光合成の反応を進行させる。 クロロフィルは、青い光と赤い光をよく吸収するため、残った緑色の光が散乱・反射されて植物の葉は緑色に見える。具体的には、クロロフィルは太陽光の主に青と赤の光を吸収し、そのエネルギーを利用して光合成を行う。この過程で、クロロフィル内の電子e-が励起され、エネルギーが高い状態になり、このエネルギーが、光化学系IとIIの反応中心に伝達され、最終的に化学エネルギーに変換される。 シアノバクテリアが出現した30億年前の地球の上空にオゾン層がなく、強烈な紫外線から逃れるために生物はある程度の海中の深さまで潜らなければならなかった。海中の生物に届く光は海の成分に大きく影響される。現在では波長の長い順に届きやすくなっているが、当時の海では様々な金属イオンや有機物が溶けており、そのような物質が緑色の光の透過を阻害したため、青と赤の光が光化学反応に使われるようになったのではないかと考えられている。 クロロフィルは、陸上植物・藻類・シアノバクテリアンなどが持ち、光合成色素の中でも、反応中心として働くクロロフィルaに対して、アンテナとして働くクロロフィルbやカロテノイドなどを「補助色素」と呼ぶこともあったが、現在では、クロロフィルaも反応中心として働くのはごく一部で、殆どはアンテナの役割を果たしていることが分かり、光合成色素分子の種類として「補助色素」といった分類はしていない。 |
太陽光を植物の葉緑体内のクロロフィルが吸収する。
(A: 光エネルギー ⇒ B: クロロフィル励起)
② 光化学系II(PSII)の活性化
励起されたクロロフィルから電子e-が放出される。
(B: クロロフィル励起 ⇒ C: 電子放出)
③ 水の酸化と酸素の生成
水分子H2Oが分解され、酸素O2、プロトンH⁺、電子e-が生成される。
(C: 電子放出 ⇒ D: 水の酸化)
④ 電子伝達系によるエネルギー移動
高エネルギーの電子e-がプラストキノンPQを通じてシトクロムb6f複合体へ移動する。
(D: 水の酸化 ⇒ E: 電子の移動)
⑤ プロトン駆動力の生成
電子e-の移動により、チラコイド膜内外でプロトン濃度勾配が形成される。
(E: 電子e-の移動 ⇒ F: プロトン駆動力)
⑥ ATPの合成
プロトン駆動力により、ATP合成酵素が働いてATPが生成される。
(F: プロトン駆動力 ⇒ G: ATP合成)
⑦ 光化学系I(PSI)の活性化
電子e-が光化学系Iに渡され、再び光エネルギーを吸収して高エネルギー状態に励起される。
(G: ATP合成 ⇒ H: 電子の再励起)
⑧ NADPHの生成
高エネルギー電子e-がフェレドキシンFdを通じてNADP⁺還元酵素に伝達され、NADPHが合成される。
⑨ NADPHの生成は、光化学系I(PSI)で光エネルギーが再び吸収され、電子がフェレドキシンに渡される。
フェレドキシン-NADP+オキシドレダクターゼという酵素が、NADP+を還元してNADPHを生成する過程では、
光化学反応ont-size : 80%;">(H: 電子の再励起 ⇒ I: NADPH生成)
⑩ 光合成の光反応、その未来への展望
多くの生物は、呼吸によってエネルギーを得ている。呼吸の過程では、有機物から電子e-が引き抜かれ、細胞内の電子回路を通じて移動する。この電子e-の流れがエネルギーを生成する原動力となる。 電子回路を通る電子は、水素イオンを移動させる働きがあり、この水素イオンの濃度差を利用して、ATP(アデノシン三リン酸)というエネルギー通貨が生成される。
光合成では、太陽光エネルギーを利用して水分子H2Oから電子e-が引き抜かれる。この電子e-は、光合成装置内のクロロフィルなどの色素分子を通じて移動し、エネルギーを伝達する。 この過程で、電子e-は最終的に二酸化炭素CO2と水H2Oと結合し、炭水化物C6H12O6を生成する。光合成の光反応は、電子e-移動を通じて光エネルギーを化学エネルギーに変換する非常に精巧なシステムであり、これが地球上の生命を支える基盤となっている。電子e-の移動のメカニズムを理解することは、光合成の全体像を把握する上で不可欠であり、また新しいエネルギー技術の開発に直結する可能性が極めて高い。
現代の研究は、この電子e-移動のプロセスをより深く理解し、さらには人工的な光合成システムの構築を目指している。
SPring-8では、光エネルギーを利用して水分子H2Oから酸素分子O2を生成する過程を詳細に解析している。フェムト秒X線自由電子レーザーを用いて、触媒が水分子H2Oを取り込み、酸素分子O2生成の準備が完了するまでの一連の動きを捉えることに成功している。 この成果は、光エネルギーを利用して水から電子e-と水素イオンhydrogen ionを取り出し、有用な化学物質を作り出す「人工光合成」の技術開発に重要な知見を提供している。
目次へ
 クロロフィルとカロテノイドなどの色素の大部分が、アンテナとして働くアンテナ複合体antenna complexは、光エネルギーを吸収し集めてその物理的エネルギーのまま反応中心複合体に渡す。アンテナ複合体は、反応中心での光化学反応を駆動するのに必要な光を吸収し、その励起エネルギーを反応中心へ渡す役目を果たす色素とタンパク質の複合体である。
クロロフィルとカロテノイドなどの色素の大部分が、アンテナとして働くアンテナ複合体antenna complexは、光エネルギーを吸収し集めてその物理的エネルギーのまま反応中心複合体に渡す。アンテナ複合体は、反応中心での光化学反応を駆動するのに必要な光を吸収し、その励起エネルギーを反応中心へ渡す役目を果たす色素とタンパク質の複合体である。緑色硫黄光合成細菌には、光エネルギー捕集のための特殊な膜外アンテナ系があり、その光捕集の中核部はタンパク質の関与しない色素のみからなっているが、それ以外は、アンテナ色素は必ずタンパク質と結合している。そのことで各色素の空間的な配置やエネルギー準位などが決められている。つまり反応中心への方向性のあるエネルギーの流れは、事前に決定されている。
この色素タンパク質複合体は、集光性タンパク質複合体と同義あれば、この複合体は、光を吸収し、そのエネルギーを反応中心reaction centerに伝達することで、光化学反応を駆動させる。 具体的には、クロロフィルやカロテノイドなどの色素分子が光を吸収し、そのエネルギーを効率的に反応中心に運ぶ役割を果たす。この間の移動は、純粋な物理現象で、それに関与する分子に化学変化は生じていない。
光化学系II(PSII)の反応中心クロロフィルはP680と呼ばれ、吸収される680nmの赤色光は、水を酸化できるほどの非常に強い酸化物とPSⅠより弱い還元物を生成する。PSIIの強い酸化物は水H2Oを酸化し酸素O2を分離し放出する。
水分子H2Oは分解され、酸素O2・プロトンH⁺・電子e-が生成される。
PSIIの反応中心には、特定の電子受容体と電子供与体が存在する。PSIIの一次電子供与体は、その特別な環境下にあるクロロフィルa分子の二量体で、P680と呼ばれる。
クロロフィルが二量体を形成することにより、反応中心により効果的に光エネルギーを吸収し、その一方、全体的に励起状態のエネルギーレベルを低下させ、より長波長でも吸収できるようにして、また、二量体を形成することにより、アンテナ系の色素から励起エネルギーを受け取るレベルを引き下げられれば、酸化還元電位を低下させ、隣接分子への電子供与もより容易になしえる。
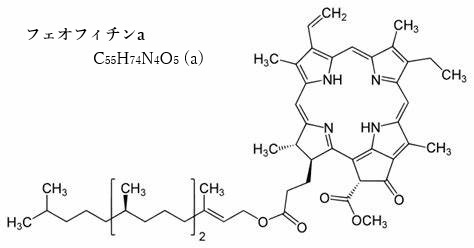
一次電子受容体として機能するのが、フェオフィチンa分子である。 この反応中心では、光エネルギーがクロロフィルP680によって吸収され、その結果、P680が励起される。励起されたP680は電子を放出し、フェオフィチンaに電子を渡す。この過程が「光合成の初期電荷分離反応」と呼ばれる。
クロロフィルaの中心に位置するMg原子が外れた分子はフェオフィチンa、クロロフィルbではフェオフィチンbになる。フェオフィチンpheophytinは、クロロフィル分子の中心にあるマグネシウムイオンMG2+が2H+に置き換わり、クロロフィルとは少し違った光学・化学的性質をっ持つようになる。
クロロフィルfとクロロフィルaは、どちらも光合成に関与する色素(葉緑素)であるが、クロロフィルaは、ほとんどすべての酸素発生型光合成生物(陸上植物・藻類・シアノバクテリア)に存在し、光エネルギーを化学エネルギーに変換する反応中心色素として機能する。一方、クロロフィルfは、2010年にオーストラリアのシャーク湾で発見された比較的新しい色素であり、クロロフィルaの2位のメチル基(R-CH3 と表される最も分子量の小さい炭化水素基)がフォルミル基(一般式はR-CHOで表される)に置換された構造を持ち、遠赤色光(波長700 nm以上)を吸収する能力がある。その構造式は C55H70O6N4Mgと考えらている。これは、クロロフィルaが吸収できない波長の光を利用できるため、特に光が少ない環境での光合成には有利に働く。波長が長くエネルギーの低い遠赤色光を吸収して、より高いエネルギーを必要とするクロロフィルaにエネルギーを渡すことが確認されている。つまり、クロロフィルfはクロロフィルaと補完的な役割を果たし、特定の環境条件下でも光合成をより効果的に行うために進化してきたと考えられている。
クロロフィルfは原核生物の一種シアノバクテリアから発見され、遠赤色光下で培養されたシアノバクテリアでは、白色光下で培養されたものと異なりクロロフィルf が新たに発現し、増えることが明らかになった。しかし、クロロフィルf は、多くの植物や藻類などが利用できない遠赤色光を吸収することができるが、具体的にどこに存在するのか、光化学反応の中でどのような機能を果たしているのかまでは解明されていない。
新たな研究によれば、クロロフィルf は光化学反応を直接駆動するのではなく、その機能をもつ別の色素にエネルギーを受け渡すことで、光化学反応を促進させることが明らかになった。また、クロロフィルfは、他のクロロフィルと比べて吸収スペクトルが長波長側にシフトしており、メタノール中での吸収ピークは706 nmと言う。 この新しい色素の発見は、光合成の研究において重要な進展が期待され、特に低光量環境での光合成効率を向上させる可能性が高くなった。
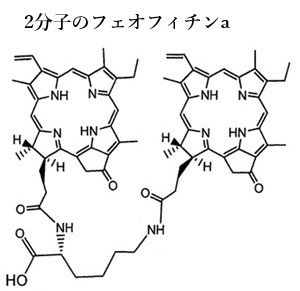 PSⅡ反応中心には2分子のフェオフィチンaが存在しており、このうちの1分子が一次電子受容体として機能する。フェオフィチンaは、クロロフィルが光エネルギーを吸収して励起された電子を受け取る役割を果たす。PSIIは、光合成の初期段階で重要な役割を果たし、水H2Oを酸素O2とプロトンH+に分解し、電子e-を供給することで、全体の光合成プロセスを支えている。
PSⅡ反応中心には2分子のフェオフィチンaが存在しており、このうちの1分子が一次電子受容体として機能する。フェオフィチンaは、クロロフィルが光エネルギーを吸収して励起された電子を受け取る役割を果たす。PSIIは、光合成の初期段階で重要な役割を果たし、水H2Oを酸素O2とプロトンH+に分解し、電子e-を供給することで、全体の光合成プロセスを支えている。フェオフィチンaは、一般的にはクロロフィルの分解産物として理解されているが、光化学反応中心内の二次もしくは三次電子受容体として存在するが、PSII反応中心においては重要な役割を果たし、一次電子受容体として機能する。
PSIIにおけるクロロフィルa分子の二量体dimerは、光合成の過程で非常に重要な役割を果たす。二量体とは、2つの同じ分子やサブユニット(単量体)が物理的または化学的な力によって結びついた分子または超分子のことを指す。PSIIの構造自体は、17個の膜貫通サブユニットと3個の膜表在性サブユニットからなる複合体で、クロロフィルaやβ-カロテン、フェオフィチンなどの色素分子が含まれている。
PSIIは、植物・藻類・シアノバクテリアのチラコイド膜に存在し、光エネルギーを化学エネルギーに変換する光化学反応を行う。PSIIの反応中心には、P680と呼ばれるクロロフィルaの二量体が存在する。この二量体は、光エネルギーを吸収して電子を励起し、最初の電荷分離を引き起こす。この過程で、P680と呼ばれる吸収される赤色光は、水を酸化(分解)できる非常に強い酸化物とPSⅠより弱い還元物を生成する。
 反応中心では、物理的光エネルギーの一部を利用して、電子がクロロフィルから電子伝達系を通して電子受容体に移動され、化学エネルギーに変えられ蓄えられる。それが、最終的にNADP+をNADPHに還元する。その一方、電子を失ったクロロフィルは強い酸化力を持つため、電子供与体から電子を受け取って再び還元される。
反応中心では、物理的光エネルギーの一部を利用して、電子がクロロフィルから電子伝達系を通して電子受容体に移動され、化学エネルギーに変えられ蓄えられる。それが、最終的にNADP+をNADPHに還元する。その一方、電子を失ったクロロフィルは強い酸化力を持つため、電子供与体から電子を受け取って再び還元される。電子供与体から電子を受け取る物質を電子受容体と呼ぶ。酸化還元反応において、電子供与体は酸化され電子を放出する。電子受容体はその電子を受け取って還元される。電子伝達系では、電子供与体から電子受容体に電子が移動し、その過程でエネルギーが生成される。このエネルギーは、細胞の様々な活動に利用される。実際の反応中心には、多くの色素から同時にエネルギーを受け取るため、より高度に活性化されている。
PSIは、光合成における重要な複合体の一つで、植物・藻類・シアノバクテリアに存在し、光エネルギーを利用して電子を移動させる役割を果たす。PSIIは、100以上の補因子を含む複雑な構造を持ち、光エネルギーを効率的に化学エネルギーに変換してATPとNADPH(還元物質)を生成し、これらを使って二酸化炭素から有機化合物を合成する。
酸素を発生する光合成では、水に由来する電子は電子伝達系を経てNADPHに入る。普通、この方向に電子は移動しにくいが、光合成生物は光エネルギーを使うことにより、これを可能にした。
20種近い高等植物・藻類・シアノバクテリアなどの葉緑体のチラコイド膜では、PSIとPSIIが連携して働き、NADP⁺が還元されてNADPHが生成される。具体的には、PSIIが光エネルギーを利用して水を分解し、酸素やプロトンH⁺、および電子を生成する。この電子は電子伝達鎖を通して移動し、最終的にPSIに到達する。PSIでは、さらに光エネルギーを吸収して電子を高エネルギー状態に励起し、その電子をNADP⁺に渡してNADPHを生成する。
このプロセスは、植物が光エネルギーを化学エネルギーに変換するための重要なステップとなる。NADPHは光合成の明反応(光依存反応)で生成される。具体的には、葉緑体のチラコイド膜で行われるPSIとPSIIの働きによって、NADP+が還元されてNADPHが生成さる。このNADPHは、光合成の次の段階である暗反応(カルビン・ベンソン回路)で、二酸化炭素を固定して糖を合成するためのエネルギー源(還元力)として利用される。
細菌でも植物細胞でも経過は大変良く似ている。光合成の炭素同化系では、光が当たると葉緑体の光化学反応系と電子伝達系の働きによりNADP+はNADPHに還元され、同化反応に利用されて、デンプンや糖類を蓄積する。光合成炭素同化系の酵素では、主に[NADP+・NADPH]が補酵素として利用される。これに対し、非同化器官の細胞では、また葉緑体でも暗所では、糖類などが分解される呼吸などに利用されるが、この時の酸化還元反応では主に[NAD+・NADH]が利用される。
NAD+/NADHはエネルギー代謝(リン酸基が転移される反応が多い)で登場するため、紛らわしいので注意が必要である。
細胞内で重要な役割を果たす補酵素で、NADは酸化型で、NADHは還元型である。NADは、細胞呼吸やエネルギー代謝において電子を受け取る役割を持ち、NADHはその電子を他の分子に渡すことでエネルギーを生成する。具体的には、解糖系やクエン酸回路で生成され、ミトコンドリア内でATPを生成するために使われる。
光合成のチラコイド反応は、その内膜上で進行する。その最終産物は、高エネルギー化合物ATPとNADPHで、それらは炭酸固定反応で糖の合成に使われる。その合成は葉緑体の中のチラコイドを囲む溶液領域ストロマで行われる。
PSIIでは、水分子H2Oが電子供与体として働き、電子e-を提供することで酸素O2が生成される。このプロセスは植物が光エネルギーを化学エネルギーに変換するための重要なステップとなる。反応中心と大部分のアンテナ複合体は、光合成膜には不可欠な要素である。真核光合成生物では、この膜は葉緑体中にあり、原核光合成生物の光合成は、その細胞膜か細胞膜から派生した膜で行われる。
原核生物は真核光合成生物のように核膜を持たず、細胞内のDNAは細胞質に剝き出しで存在する。従って細胞小器官organelle、例えば、葉緑体やミトコンドリアなどは存在しない。細胞小器官が高度に発達していることが、真核細胞を原核細胞から区別する。原核細胞は約35億年前に出現し、地球上の最初の生命体とされている。その後、約20億年前に真核細胞に進化した。真核細胞は核膜で囲まれた核を持ち、より複雑な構造体を保有した。
原核光合成生物には、酸素発生型と非発生型のものがあり、シアノバクテリア(藍藻)は酸素を発生させる光合成を行い、光合成細菌(紅色細菌、緑色細菌)は酸素を発生させない光合成を行う。シアノバクテリアは約27億年前に誕生し、光合成を行うことで酸素を生成し始めた。この過程により、地球の大気中の酸素濃度が増加し、現在の酸素豊富な地球環境が形成された。シアノバクテリアの光合成は、地球の「大酸化イベント」と呼ばれる現象を引き起こし、これが約24億年前に起こったとされている。このイベントにより、地球の大気中の酸素濃度が劇的に増加し、多くの生物が酸素を利用できるようになった。
生物の酸素呼吸の優位性
酸素呼吸(好気呼吸)は、生物にとって大きなしかも重要な優位性がある。酸素呼吸は、グルコース1分子から最大38分子のATPが生成される。これは、嫌気呼吸(酸素を使わない呼吸)に比べて非常に効率的で、また酸素呼吸は、長時間にわたって安定したエネルギーの持続と、酸素が豊富な環境であれば、効率的にエネルギーを生成できるため、広範な多様な環境への適応と、高等動物として多彩な活動や複雑な生理機能の維持が可能になる。
しかも、酸素呼吸の副産物は主に水と二酸化炭素であれば、これらは他の生物にとって比較的無害であり、容易に排出でき再利用もされる。また生物の排出物資源については、バイオマスが代表的な例であるが、再生可能な生物由来の有機性資源(石油・木材・生ごみ・家畜の排泄物)として、特にバイオマス発電にも利用される。
 宇宙創始以来、外部からのエネルギーの供給なしで物質が作られたことはない。H2OやCO2にしても自発的にグルコースは作りえない。植物の光合成能力を駆動するためには光エネルギーの供給が不可欠である。
宇宙創始以来、外部からのエネルギーの供給なしで物質が作られたことはない。H2OやCO2にしても自発的にグルコースは作りえない。植物の光合成能力を駆動するためには光エネルギーの供給が不可欠である。CO2 + H2O + 光源
として利用されるだけでなく、さまざまな生化学的反応に関与する。例えば、グルコースは解糖系やクエン酸回路を通じてATPを生成し、細胞のエネルギー供給を支えている。
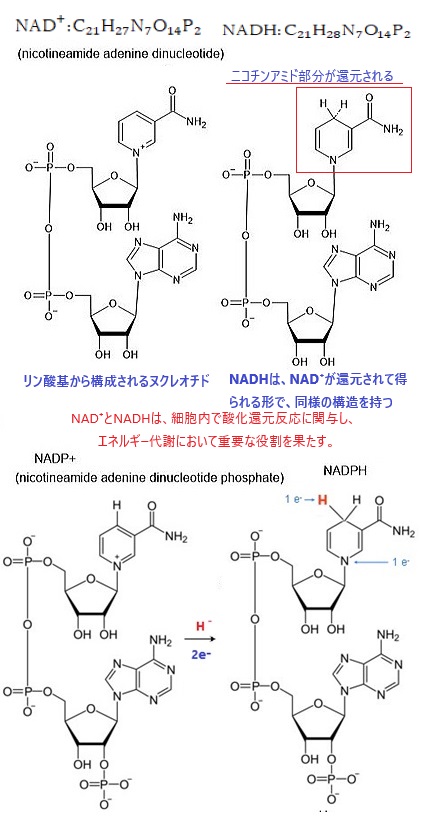
NADP(nicotinamide adenine dinucleotide phosphate)の化学式C21H21N7O17P3は、NADにリン酸基がついたもので、それ以外の部分の構造はNADと同一である。
一方、光合成経路は植物や一部の細菌が光エネルギーを利用して有機物を合成する過程で、光依存反応とカルビン・ベンソン回路(暗反応)の2つの主要な段階がある。グルコースの代謝過程である解糖系とクエン酸回路(TCAサイクル)では、異なる量のATPとNADH(およびNADPH)が生成される。
具体的には、解糖系では、1分子のグルコースが分解されて2分子のATPと2分子のNADHが生成され後、生成されたピルビン酸はミトコンドリアに入り、クエン酸回路(TCA回路)に入り、この重要な代謝経路で、糖質・脂質・タンパク質の酸化の最終段階に入り、エネルギーが生成される。
クエン酸回路では、さらに多くのNADHとFADH2が生成され、これらは電子伝達系に送られる。電子伝達系では、これらの高エネルギー電子キャリアが酸化され、そのエネルギーを利用してATPが大量に生成される。最終的に、1分子のグルコースから通常、合計で約30~32分子のATPが生成されるとされている。
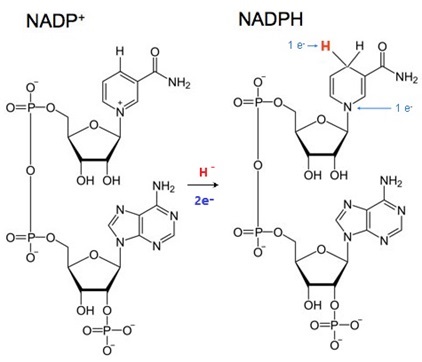
NADPHは主にペントースリン酸経路で生成される。リン酸は、補酵素NADPの重要な構成要素でるが、酵素に認識される際の構造的な特異性を生み出すために存在するものであって、リン酸基を他に供与したりするのに使われるものではない。NADPは、主に光合成や解糖系などの代謝経路で電子伝達体として機能する。
NADPは、酸化型(NADP+)と還元型(NADPH)の2つの状態を持ち、これらは生体内の酸化還元反応において重要な役割を果たす。例えば、光合成では、光化学系複合体Iによって生成された還元型フェレドキシンからNADP+に電子が伝達され、NADPHが生成される。NADPH(C21H29N7O17P3)は、生物の細胞内で重要な役割を果たす補酵素で、特に、還元反応に関与し、エネルギー代謝や抗酸化作用において重要となる。
エネルギー = (CH2O) + O2 となる。
CH2O(ホルムアルデヒド)は、グルコース分子(C6H12O6)の6分の1を表す。
ホルムアルデヒドはグルコースの合成過程で重要な役割を果たす。ホルムアルデヒドからグルコースを合成する過程は、主にホルモース反応formose reactionとして知られている。この反応は、ホルムアルデヒドがアルドール縮合を経て、C-C結合(新しい炭素-炭素結合を形成するため、有機合成において非常に有用)を形成し、糖類の骨格を構築し、最終的にグルコースなどの糖類を生成する一連の化学反応である。アルドール反応は、グリコールアルデヒドがさらにホルムアルデヒドと反応し、グリセルアルデヒドを生成する。生成されたアルデヒドがエピメリゼーションやカニッツァロ反応を経て、様々な糖類(例えばグルコース、フルクトース、マンノースなど)を生成する。この反応は、溶液中に水酸化物イオンOH⁻が多く含まれている塩基性条件下で進行する。グルコースだけでなく、フルクトースやマンノースなどの他のヘキソースHexose(六炭糖)も生成される。
(具体例として、アセトアルデヒド(CH3CHO)とベンズアルデヒド(C6H5CHO)のアルドール縮合反応を考えると、以下のような反応が起こる。
CH3CHO + C6H5CHO → C6H5CH(OH)CH2CHO( 3-フェニル-2-ヒドロキシプロパナール)
この反応は、新しい、C-C結合を形成するため、有機合成において非常に有用となる。
このC6H5CH(OH)CH2CHO化合物は、香料や医薬品の中間体として使用されることが多い。また、化学反応の研究や合成化学の分野でも重要な役割を果たしている。)
 また、グルコースは生体内でエネルギー源として利用されるだけでなく、さまざまな生化学的反応に関与する。例えば、グルコースは解糖系やクエン酸回路を通じてATPを生成し、細胞のエネルギー供給を支えている。
また、グルコースは生体内でエネルギー源として利用されるだけでなく、さまざまな生化学的反応に関与する。例えば、グルコースは解糖系やクエン酸回路を通じてATPを生成し、細胞のエネルギー供給を支えている。NADP(nicotinamide adenine dinucleotide phosphate)の化学式:C21H21N7O17P3は、NAD:C21H27N7O14P2にリン酸基(1価の官能基、 構造式は H2PO4 で3表される。)がついたもので、NADは、主に細胞呼吸に関与し、エネルギー生成に重要な役割を果す。NADは、解糖系やクエン酸回路で電子を運び、ATPの生成を助ける。
NADPは、主に光合成や脂質合成などの同化反応に関与する。NADPは、特に葉緑体での光合成において重要で、カルビン回路で二酸化炭素の固定に使われる。
一方、光合成経路は植物や一部の細菌が光エネルギーを利用して有機物を合成する過程で、光依存反応とカルビン・ベンソン回路(暗反応)の2つの主要な段階がある。グルコースの代謝過程である解糖系とクエン酸回路(TCAサイクル)では、異なる量のATPとNADH(およびNADPH)が生成される。
具体的には、解糖系では1分子のグルコースから2分子のATPと2分子のNADHが生成される。その後、クエン酸回路と電子伝達系を経て、さらに多くのATPが生成される。NADPHは主にペントースリン酸経路で生成される。リン酸は、補酵素NADPの重要な構成要素でるが、酵素に認識される際の構造的な特異性を生み出すために存在するものであって、リン酸基を他に供与したりするのに使われるものではない。NADPは、主に光合成や解糖系などの代謝経路で電子伝達体として機能する。
NADPは、酸化型(NADP+)と還元型(NADPH)の2つの状態を持ち、これらは生体内の酸化還元反応において重要な役割を果たす。例えば、光合成では、光化学系複合体Iによって生成された還元型フェレドキシンからNADP+に電子が伝達され、NADPHが生成される。NADPH(C21H29N7O17P3)は、生物の細胞内で重要な役割を果たす補酵素で、特に、還元反応に関与し、エネルギー代謝や抗酸化作用において重要となる。
目次へ
 PSIIは、光合成の過程で重要な役割を果たす複数のサブユニットからなるタンパク質複合体である。特に、反応中心にはD1とD2という2つのタンパク質で構成された主要な中核部分があり、加えていくつかのサブユニット、アンテナ複合体が含まれている。
PSIIは、光合成の過程で重要な役割を果たす複数のサブユニットからなるタンパク質複合体である。特に、反応中心にはD1とD2という2つのタンパク質で構成された主要な中核部分があり、加えていくつかのサブユニット、アンテナ複合体が含まれている。第一電子供与体クロロフィルと、その他のクロロフィルやカロテノイド、そして電子受容体であるフェオフィチンやプラストキノンは、すべて膜タンパク質D1とD2に結合している。
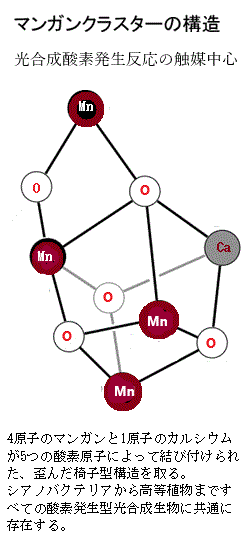
D1サブユニットは、 反応中心タンパク質で、マンガンクラスターを持ち、水H2Oの分解と酸素O2の生成に関与している。マンガンクラスターは、光合成における酸素発生反応の中心的な触媒で、PSIIの酸素発生複合体oxygen evolving complex(OEC)内にあって、水分子を酸化して酸素分子を生成する。
マンガンクラスターは、4つのマンガン原子と1つのカルシウム原子が5つの酸素原子によって結び付けられた歪んだ構造をしている。この構造は、光エネルギーを利用して水を酸化し、酸素を生成するために必要な高い酸化力を蓄える役割を果たす。
この反応は、光合成生物が太陽光を利用して水を分解し、酸素を放出する唯一のプロセスであり、光合成の酸素発生複合体は、この反応をする唯一無二の生化学反応能力を保持し、大気中のほぼすべてのO2の源泉となる。地球上の生命の進化において非常に重大な役割を果たしている。
水は以下の化学反応で酸化される。
2H2O ⇒ O2 + 4H+ + 4e-
この式は、2分子の水から4電子を引き抜き、1分子の酸素と4水素イオンを生成することを示す。
D2サブユニットは、 D1と非常に似ているが、マンガンクラスターを持たず、電子伝達の役割を担っている。D1サブユニットを経由してプラストキノンPlastoquinone(PQ)に伝達される。これらのサブユニットは、光エネルギーを化学エネルギーに変換する過程で重要な役割を果たす。PSIIでは、光エネルギーがクロロフィル分子(P680)によって吸収され、そのエネルギーが電子e-を高エネルギー状態にする。この高エネルギー電子e-は、まずフェオフィチンaに渡され、次に、フェオフィチンaから電子e-はプラストキノンに渡される。
プラストキノンは、光合成の光化学反応における電子伝達体として機能する。プラストキノンは、PSIIから電子を受け取り、シトクロムb6f複合体を介してプラストシアニンに電子を渡す。プラストシアニンは、光合成を行う植物や藻類に見られる銅含有タンパク質である。主に電子伝達を媒介する役割を果たし、PSIIのシトクロムb6f複合体からPSIのP700+へ電子e-を運ぶ。
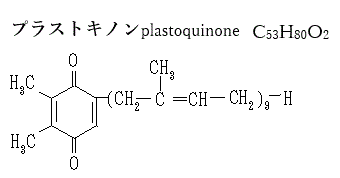 |
| プラストキノンは、光化学系IIからの電子を受け取り、チラコイド膜を通過してシトクロムb6f複合体に電子を渡す役割を果たす。この過程でプロトンH+も移動し、ATP合成に必要な電気化学的勾配を形成する。 さらに、プラストキノンは抗酸化剤としても機能し、活性酸素種を還元することで細胞膜を保護する。 プラストキノンは、電子を受け取るとプロトンH+も取り込み、プラストキノールPQH₂に変わる。このプロセスは、チラコイド膜を通じてプロトン勾配を形成し、ATP合成に必要なエネルギーを提供する。 この一連の電子伝達反応により、光エネルギーが化学エネルギーに変換され、最終的にATPとNADPHが生成される。これらは、カルビン回路での二酸化炭素固定に利用される。 |
放出された電子e-は、フェレドキシンなどの電子伝達体を通じてNADP+に渡され、NADPHが生成される。このプロセスは、光合成のリニア電子伝達反応の一部であり、植物がエネルギーを効率的に利用するために不可欠である。また、P700+の蓄積は、活性酸素種reactive oxygen species (ROS) の生成を抑制し、植物を酸化ストレスから守る役割も果たす。
光合成の過程で重要な役割を果たす光合成のリニア電子伝達反応linear electron transportは、PSIIとPSIを経て、水H2Oから引き抜かれた電子e-がNADP+を還元してNADPHを生成するプロセスである。この過程で酸素O2が生成され、ATPも合成される。 リニア電子伝達反応は、非循環的電子伝達とも呼ばれ、循環的電子伝達と対比される。循環的電子伝達では、電子e-が再びPSIに戻るため、NADPHは生成されず、ATPのみが生成される。このリニア電子伝達反応では、光合成の暗反応「カルビン・ベンソン回路」で使用されるNADPHとATPが生成される。
植物のミトコンドリアは、呼吸鎖電子伝達系を通じてATPを生成する。ATPは細胞のエネルギー通貨として機能し、さまざまな生理的プロセスを支える。ミトコンドリアは、クエン酸回路(TCA回路)を通じて代謝産物を生成し、これらは他の合成経路に供給する。 例えば、光合成が行われる葉緑体とミトコンドリアは相互作用し、光合成で生成されたNADPHがカルビン回路で消費されない場合、ミトコンドリアで酸化されることがある。
PSII系で光エネルギーを吸収し、水を分解して酸素O2やプロトンH+、および電子e-を生成する。電子伝達鎖は、ミトコンドリア内で行われる重要なプロセスで、NADHやFADH2(フラビンアデニンジヌクレオチド)から電子e-を受け取り、これを一連の酵素複合体を通じて酸素に渡すことでATPを生成する。
FADH2は、エネルギーキャリアとして重要な役割を果たす分子で、FAD(フラビンアデニンジヌクレオチド)が還元されてできる形態で、ミトコンドリア内での酸化的リン酸化の過程で使われる。その詳細は、FADH2はクエン酸回路や脂肪酸のβ酸化で生成され、その後、FADH2が酸化されることで、電子が電子伝達系に供給され、最終的にATP(アデノシン三リン酸)を生成する。FADH2は1分子あたり2分子のATPを生成することができる。
生成された電子は一連の電子キャリアを通じて移動し、その過程でプロトンH+がチラコイド膜を越えて移動する。
PSI系で電子がさらに光エネルギーを吸収し、NADP+をNADPHに還元する。また、プロトンH+の濃度勾配を利用してATPが合成される。
目次へ