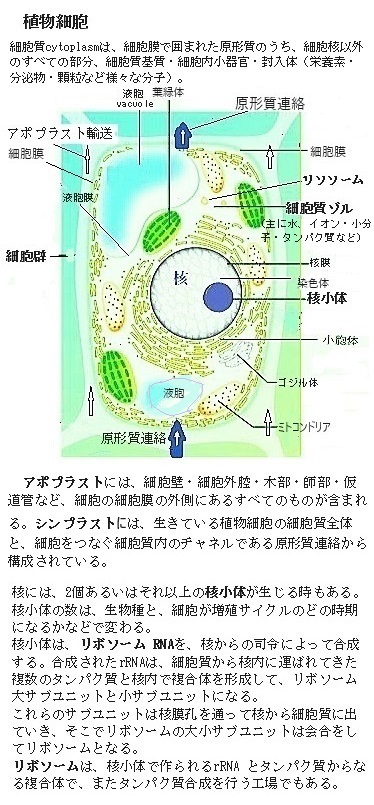
植物細胞の体内循環
TOP 車山お知らせ 車山ブログ 歴史散歩 車山高原野鳥 たびネット信州
|
 諏訪湖の 北岸にあたる 下諏訪の湖岸を歩くと 富士山の全景が!! |
「原形質膜plasma membrane」と「細胞膜cell membrane」は、同じものを指している。この膜は、細胞の内部環境を保護し、物質の出入りを制御する役割を担う生体膜で、具体的には、脂質二重層とその中に埋め込まれたタンパク質から構成されており、そのタンパク質は膜タンパク質と呼ばれている。細胞膜は、イオンや有機化合物に対する選択的透過性によって、細胞内や細胞小器官への物質の出入りを制御し、さらに、細胞膜は細胞接着やシグナル伝達などのプロセスに関与し、細胞外構造の接着表面としても機能している。また選択的透過性を持っている。
 細胞膜を、電子顕微鏡で見ると、約10㎚(ナノメートル)の極薄い膜で、タンパク質のかたまりを含んだ2層のリン脂質の膜(脂質二重層)からできている。
細胞膜を、電子顕微鏡で見ると、約10㎚(ナノメートル)の極薄い膜で、タンパク質のかたまりを含んだ2層のリン脂質の膜(脂質二重層)からできている。(酸素原子の大きさは、約0.1㎚。葉緑体の大きさは植物によって異なるが、一般的には約10㎚程度。ただ、葉緑体は光合成や他の重要な機能を担当する細胞内小器官であるため、そのサイズは植物の種や細胞の成熟度によって変化する。)
その選択的透過性とは、細胞膜が特定の物質だけを通す性質を指す。具体的には、細胞膜のリン脂質部分を通過できるのは小さな分子である酸素や二酸化炭素、または疎水性の分子である。一方で、アミノ酸や糖、また小さくともイオンはリン脂質部分を透過できないため、膜を貫通した「輸送タンパク質」を介して通過する。
(陽子数つまり原子番号が大きいイオンの方が回りの電子を“グッ”と引きつけ、イオン半径は小さくなる。O2-は8個,F-は9個,Na+は11個,Mg2+は12個,Al3+は13個の陽子を含んでおり、陽子数が多いほど周りの電子を“グッ”と引き付けイオン半径はより小さくなる。 よって、イオンの大きさは、O2- > F- > Na+ > Mg2+ > Al3+となる。)
「輸送タンパク質」は、生体膜を貫通し、物質の輸送をするタンパク質の総称で、その細胞膜には様々なタンパク質が埋め込まれており、物質の出入りを制御している。
チャネルchannelとしては、イオンチャネルやアクアポリンなどがある。イオンチャネルは膜を貫通する穴を持ち、選択的にイオンや低分子化合物を通すことできるため、エネルギーを必要としない。
アクアポリンaquaporinは、細胞膜に存在する細孔poreを持ったタンパク質で、水分子のみを透過させる「水チャネル」として知られている。アクアポリンは、人間の多くの細胞や一部のバクテリア、植物などの有機生命体にとって、水分子を輸送する不可欠なタンパク質である。
能動輸送体active transportersはエネルギーを必要とする輸送体で、膜の向きとは関係なく化学エネルギーや電気化学ポテンシャル差を利用する。例えば、ATPアーゼを持つイオンポンプやABC輸送体がある。
ATPアーゼは、アデノシン三リン酸(ATP)のリン酸結合を加水分解する酵素群の総称で、ATPは生体内のエネルギー通貨と呼ばれ、エネルギーを要する生物活動に関連したタンパク質の多くが、この酵素の活性に依存している。具体的な役割としては、ATPの合成・筋収縮・細胞内物質輸送・細胞外物質輸送・イオン濃度勾配の作成・解毒作用・発光・発電・あらゆる生体膜の融合・膜タンパク質の品質管理など生体内のさまざまな生命活動に貢献し、その領域は極めて広い。
植物細胞膜における小胞輸送に関与するATPアーゼの一例として、液胞型ATPアーゼvacuolar ATPase(V-ATPアーゼ)がある。これは、ATPのエネルギーを利用してプロトン(H+イオン)を、膜を越えて輸送する酵素を言う。このプロセスにより、細胞内の小胞や小器官organelleの内部を酸性化する。この酸性化は、リソソームやエンドソームなどの小胞が正常に機能するために重要である。
細胞・生物は数多くの複雑な生命反応を効率的に遂行するために、その機能・役割にとって最適な環境やタンパク質・脂質成分などを保持する器官形成や細胞内の小器官organelle形成という巧妙な手段を用いる。 細胞膜に囲まれた空間では、さまざまな生理的プロセスが行われるが、そのpHは細胞内の機構・機能にとって重要な環境因子の一つであり、厳密に調節されている。例えば細胞質では中性付近に調節されている。一方、分泌経路やエンドサイトーシス経路に位置するゴルジ装置・液胞・分泌顆粒・エクソソーム (小胞)などの細胞内の小器官organelleはその内腔側がpH 4.5~6.5程度の弱酸性に保たれる。大まかにいってタンパク質の流れに沿ってより酸性側に傾くpH勾配を形成している。それがゆえに、これらは総称して酸性オルガネラと呼ばれる。
(エンドサイトーシスendocytosisとエキソサイトーシスexocytosisは、細胞が物質を取り込んだり放出したりする重要なプロセスである。この二つのプロセスにより、細胞膜を直接通過できない大きな分子や微生物、老廃物などが細胞内外に運ばれる。)
V-ATPアーゼは、ATPで駆動される回転モーターを持ち、2つの主要な部分から構成されている。上部のモーターはATPを分解してエネルギーを生成し、そのエネルギーで下部のモーターが回転し、プロトン(H+イオン)を膜を越えて輸送する。
この酵素は、細胞内のpHバランスを維持し、タンパク質の品質管理や神経伝達、免疫反応など、さまざまな生理機能をサポートしている。他にも、植物細胞の小胞輸送に関与するATPアーゼとして、P4型ATPアーゼがあり、これは細胞内や細胞表面の膜から小胞を生成する際に重要な役割を果たす。
Na+/K±ATPアーゼ(ナトリウム-カリウムポンプ)は、細胞膜内にあるタンパク質で、細胞内からナトリウムイオンNa+を排出し、代わりにカリウムイオンK+を取り込む役割を果たす酵素である。その機能より細胞の浸透圧の調節や活動電位の発生などで、筋肉の収縮や神経細胞の活動電位の発生など、多くの生理的プロセスに関与する。
植物細胞膜の活動電位は、動物細胞と同様に、イオンチャネルの働きによって発生する。その基本的なメカニズムは、植物細胞が刺激を受けると、細胞膜の電位依存性イオンチャネルが開き、 脱分極: ナトリウムイオンNa⁺やカルシウムイオンCa²⁺が細胞内に流入し、膜電位が正の方向に変化する。カリウムイオンK⁺が細胞外に流出すると、膜電位が元の負の状態に戻る。一時的に膜電位が静止電位よりもさらに負の状態になることがある。 このようにして、活動電位が発生し、細胞内外の情報伝達が行われると、オジギソウやハエジゴクのように、活動電位を利用して素早い運動を行うことができる。
一方、気孔は植物の葉や茎に存在する小さな開口部で、ガス交換や蒸散を調節する。気孔の開閉は、主に孔辺細胞の膨張と収縮によって制御される。
Na+/K±ATPアーゼ自体は動物細胞に特有のものであり、植物の気孔の開閉には、植物細胞にも類似の独自のイオンポンプがあり、それにより取り込むカリウムイオンK+の濃淡が浸透圧作用として働き、孔辺細胞の膨張と収縮に関与し、気孔の開閉を調節している。
イオン輸送性ATPアーゼは、ATPの加水分解エネルギーを使って、通常では生体膜を透過できないイオンの輸送を行うタンパク質群である。F型・A型・V型・P型の4つのサブタイプがあり、イオン濃度勾配を利用してATP合成や逆反応のATP加水分解を行う。
ABC ATPアーゼは、ATP Binding Cassette (ATP結合カセット)と呼ばれるATP結合部位を持つタンパク質で、物質の取り込みや排出に関与する。
AAA ATPアーゼは、細胞内小器官への輸送や膜融合、またDNA複製や転写調節などに関与するタンパク質である。これらのATPアーゼは、生体内のエネルギーの関与するさまざまな反応に寄与しており、生命活動に不可欠なタンパク質である。
ABC輸送体(ATP-binding cassette transporter )は、ATP結合カセット輸送体ATP-binding cassette transportersの略称でABCトランスポーターまたはABCタンパク質とも呼ばれる細胞膜を通して物質を輸送するためのタンパク質で、ATPのエネルギーを利用して様々な基質を運ぶ。ABC輸送体は、細胞外から栄養分を取り込み、細胞内から不要物を排出する。また外来の薬物の輸送に関与する。
受動輸送体passive transportersは、電気化学ポテンシャル差をエネルギー源として物質を輸送するキャリアーである。原核生物では陽子proton、真核生物ではナトリウムイオンNa+が使われることが多い。
 |
| 葉が緑色に見えるのは葉の中にクロロフィル(葉緑素)という緑色の色素があるからで、葉緑素は、 細胞の中にふくまれる葉緑体の中のグラナというものに含まれている。 グラナは、電子顕微鏡で見ると直径が0.4~0.6μ(1ミクロンは1,000分の1mm)、円板状をしておりリ 脂質とたんぱく質の円板が互い違いに層状に積み重なった構造で、隣り合ったグラナどうしは、 薄い膜を介して結び付いている。 |
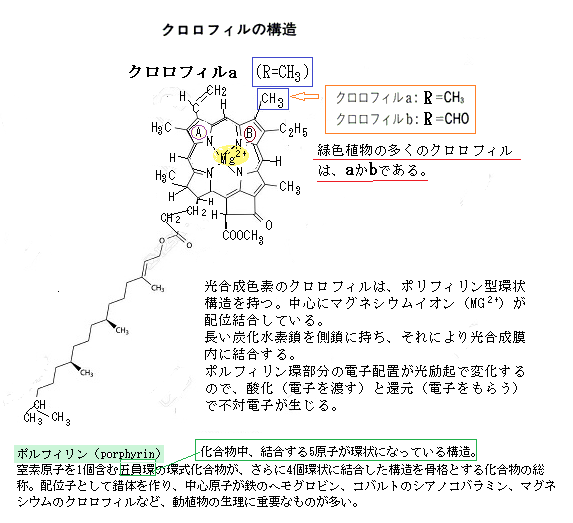
葉緑体にもMgが結合し、光合成に欠かせない元素になる。クロロフィル(葉緑素)は光合成の明反応で光エネルギーを吸収する役割を持つ化学物質で、葉の葉緑体に存在している。マグネシウムMgは特に葉緑素の構造において重要で、光合成において光エネルギーを吸収する色素として、その中心にマグネシウムイオンMg²⁺が配位している。マグネシウムはクロロフィルのテトラピロール環の中心に配位し、光エネルギーを効率的に吸収して化学エネルギーへ変換する。マグネシウムは、二酸化炭素と水を原料にして有機物の炭水化物を合成する際に欠かせない元素である。
マグネシウムは細胞内で水和した2価の陽イオンMg2+として存在し、様々な酵素反応の補因子としての役割がある他、核酸やタンパク質の構造維持に不可欠で、特にATPが関与する酵素反応にはマグネシウムが必要となる。Mgは植物においては、葉緑素のテトラピロール環の中心に配位する。
マグネシウムは、植物の生命維持に必要な必須栄養素の一つだが、根から吸収されたマグネシウムは、マグネシウム輸送体タンパク質である「CST2」により、細胞内の巨大な細胞内小器官「液胞」に輸送され貯蔵される。この貯蔵システムが気孔開口に重要であることが明らかになった。この機構により、植物はマグネシウムを恒常的に維持し、正常な成長と発育が支えられる。この発見は、植物のマグネシウム利用を理解する上で重要な知見を提供し、マグネシウム含量を高めた高栄養価の農作物の開発など、農業分野での開発応用が期待されている。
目次へ
樹木を含め全ての植物は太陽の光エネルギーを利用し光合成によって、空気中の二酸化炭素と根から吸収した水から有機物を合成する。有機物は炭素原子と水素原子を含む分子の化合物で、可燃性のものが多い。有機物は燃えやすいものが多く、メタン CH4は最も単純な有機物ガスで、メタンガスを燃やすと二酸化炭素CO2や水H2Oが発生する。ガソリンも同様で、その燃焼反応は
C8H18 + 12O2 → 8CO2 + 9H2O となる。
紙も燃えれば二酸化炭素と水を発生させるから有機物に分類される。逆に、無機物は炭素を含まない物質で、例えばガラスはシリカSiO2(二酸化珪素)が主成分の珪砂(けいしゃ)と呼ばれる砂がガラスの主原料ある。鉄や銅なども無機物に分類される。 無機物とは炭素を含まない物質で、燃やすと一般的に灰燼するが二酸化炭素は発生しない。この違いが重要で、有機物と無機物を見分けることを容易にする。
この動物と異なる植物の最大の特性で、その機能によって、人類に食糧や木材を初めとする繊維、薬などなどの有機素材や、エネルギーなどが供給され、さらに地球環境の保持にも貢献している。何よりも酸素を補給してくれる。
しかし、光合成によって作られる主な産物である糖が、そのままで植物のさまざまな組織に行き渡り多様な成分になるわけではない。例えば、樹木の幹では葉で光合成によって作られた糖が幹の形成層に移動し、ここで新しい細胞が作られ、幹の成分が形成される。そのため、幹に多い細胞壁の成分であるセルロースやリグニンを糖から作り変えるためにはエネルギーが必要となる。そのエネルギーは糖を酸素呼吸によって酸化して生物のエネルギー通貨であるATPを生産し、これによって作り変えが進行する。
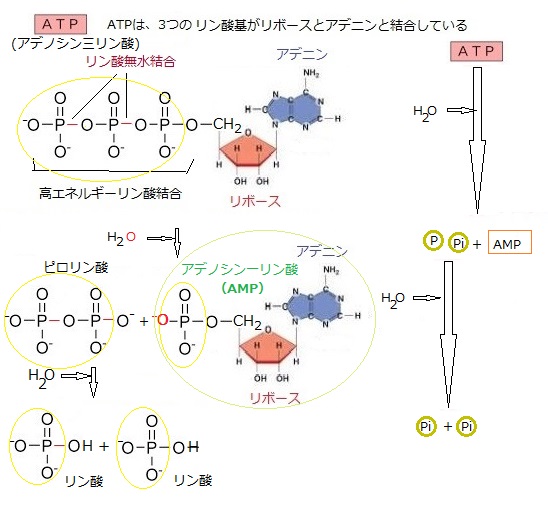 |
| ATP(アデノシン三リン酸)の分子構造のアデニンはプリン塩基の一種で、他の分子との相互作用において重要な役割を果たす。 例えば、酵素や受容体との結合において、アデニンの構造が特定の認識サイトに適合することで、ATPが正確に機能することができる。 リボースは五炭糖(化学式はC₅H₁₀O₅)でリボ核酸(RNA)の構成成分でもあり食品添加物や運動後の栄養補助食品としても利用されている。 具体的には、アデニンがリボースと結合してアデノシンを形成し、そのアデノシンに3つのリン酸基が結合している(アデノシン三リン酸)。この構造により、ATPはエネルギーを貯蔵し、必要に応じて放出することができる。 ATPの3つのリン酸基同士の結合は「高エネルギーリン酸結合」と呼ばれ、多量のエネルギーが蓄えられている。この結合が切断されると、エネルギーが放出され、さまざまな生体反応に利用される。 またADPにリン酸基が再結合することで、ATPが再生成される。このプロセスは、呼吸や光合成などの代謝反応によって行われる。 ATPは生体内でエネルギー通貨として機能し、様々な生化学反応において重要な役割を果たしながら、物質の能動輸送なども担う。 |
動物が食物を酸素呼吸によって酸化して生きていく上に必要なATPを獲得しているように、植物は光合成産物である糖を酸素呼吸によって酸化して、植物体の維持に必要なエネルギーATPを生産する。
樹木は、葉以外の組織でも昼夜分かたず、酸素呼吸によるATP生産が、樹木の成長と維持には欠かせない。樹木や作物がこれらのために酸素呼吸によって光合成産物である糖を消費する割合は、植物の種類によって異なるが、少なくとも50%位にはなる。
観葉植物を光が余り強くない室内に長い間おいておくと次第に衰えていくのは、呼吸による糖の消費が、弱光下での光合成による糖の生産能力を上回り、やがて酸素呼吸に必要な糖が枯渇し、成長や植物体の維持に必要なATPを充分に補給できなくなるためである。
植物は、酸素を海綿状組織などに細胞間隙を介して拡散させ、内部の細胞の表面に届ける。細胞を取り囲む原形質膜は比較的容易に酸素を透過するため、酸素は溶存酸素の形で細胞質中のミトコンドリアの表面に到達し、次いでミトコンドリア膜を介して更に内部に取り込まれ、呼吸の過程で消費される。細胞間隙が少ない領域では、細胞表面から取り込まれた溶存酸素が濃度平衡の仕組みで細胞から細胞へと輸送され、酸素を消費する組織に補給されるものと考えられている。
(濃度平衡定数concentration equilibrium constant【K】は、化学反応の平衡状態の維持により、物質の濃度によらず一定の値を保つ。
具体的、可逆反応(A ⇆ B)において、平衡定数 (K) は次の関係式で表される。
平衡定数は、可逆反応の平衡状態を表す指標で、具体的には、反応式の係数によって次のように表される。
K = [C]c [D]d / [A]a[B]b
ここで、(A), (B), (C), (D)は反応物や生成物を表し、それぞれの濃度を([A]),([B]), ([C]), ([D])とし、
その指数(a), (b), (c), (d)を反応式の係数とすれば、
平衡定数は、温度が一定の場合、物質の量によらず一定の値を保つ。通常は物質のモル濃度(mol/L)で表される。
ここで、[A] と [B] はそれぞれ反応物、[C]と[D]はその生成物としてモル濃度で示す。温度が一定であれば、濃度平衡定数 (K) は変わらないが、
温度を変えると(K) は絶対温度 (T) の関数となる。)
一般的には太い木の真ん中などには呼吸活性の高い組織で構成されている。樹木は葉以外の組織でも昼間に酸素呼吸を行い、成長や維持に欠かせないATPを生成している。樹木や作物が酸素呼吸によって光合成産物である糖を消費する割合は、植物の種類によって異なるが、少なくとも50%と高い数値を示す。
植物は、気孔を通じて酸素を取り込む。気孔は葉の表面に存在し、酸素は葉の内部に拡散する。植物の体内細胞、例えば太い木の真ん中の細胞でも呼吸を行っている。これらの細胞は、細胞質の中のミトコンドリアで酸素を利用する。酸素は細胞質中のミトコンドリアの表面に到達し、呼吸の過程で使用される。
植物は酸素を取り込むための、循環系やヘモグロビンのような担い手を保有していない。代わりに、酸素は細胞間隙を通って拡散し、内部深くの細胞の表面にまで到達する。細胞を取り囲む原形質膜は酸素を透過しやすいため、酸素は溶存酸素の形で細胞質中のミトコンドリアの表面に到達し、呼吸の過程で利用される。細胞間隙が少ない領域では、細胞表面から取り込まれた溶存酸素が濃度平衡の仕組みで細胞から細胞へと輸送され、酸素を消費する部分に供給されることが考えられる。
なお、太い木の真ん中などは呼吸活性の高い組織で構成されていないことが一般的である。樹木の幹は、生命活動を維持するために呼吸を行っている。この呼吸は、葉緑体による光合成とは異なり、根から吸収した水と養分を使って行われている。
幹の表面にある気孔は、樹木の呼吸に重要な役割を果たしている。幹の細胞呼吸をサポートする気孔は、葉と同様で酸素を吸収し、二酸化炭素を排出する。特に若い枝や新しい成長部分での細胞呼吸はより活発である。 気孔の開閉は、孔辺細胞と呼ばれる特殊な細胞によって制御され、これによりガス交換が調整される。樹木の幹の細胞呼吸は、葉緑体がない幹部分における呼吸であるが、植物の健康と成長にとって非常に重要な機構と言える。
幹の細胞呼吸内で行われる呼吸プロセスでも、細胞は酸素を消費し、糖や他の有機物を分解してエネルギーを生成する。このエネルギーは、成長や代謝に利用される。
水分輸送は、幹が根から吸収した水と養分を葉や他の部分に運ぶ重要な役割も果たす。この水分輸送は、幹内の導管を通じて行われる。
目次へ
植物の成長は、土壌中の水や溶存ミネラルをいかに取り込むかにかかっている。植物の根系には、水や溶質を取り込むために必要な組織が備わっている。その水分やミネラルの輸送に関わる植物の組織には、大きく分けてアポプラストapoplastとシンプラストsymplastという2つの区画がある。
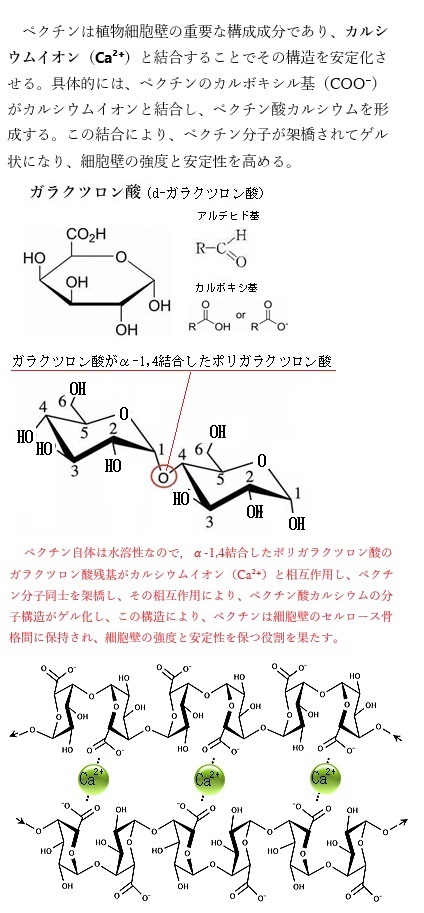
アポプラストには、細胞壁・細胞外腔・木部・師部・仮道管など、細胞の細胞膜の外側にあるすべてのものが含まれる。多細胞陸上植物内の細胞は細胞壁同士で接着されている。植物の細胞壁同士を接着する役割を果たしているのは、ペクチンという多糖類で、細胞壁間に存在し「セメント」として機能し、隣接する細胞の細胞壁をしっかりと結びつけている。
 ペクチンの構造式は、主にガラクツロン酸galacturonic acidがα-1,4結合したポリガラクツロン酸polygalacturonic
acidから成り立っている。
ペクチンの構造式は、主にガラクツロン酸galacturonic acidがα-1,4結合したポリガラクツロン酸polygalacturonic
acidから成り立っている。ペクチンは、植物の細胞壁に存在する多糖類で、特に、カルシウムイオンCa²⁺との架橋によって巨大なゲル状の分子構造を形成する。このペクチン酸カルシウムの分子構造は、植物細胞壁の重要な構成要素となり細胞壁の強度を高める。 このようにして、ペクチンはセルロース繊維の間隙を埋め、細胞壁の物理的な強度を維持する役割を果たす。また、カルシウムは細胞膜の安定化にも寄与し、植物の成長や発達において重要な役割を担っている。
ペクチン酸カルシウムの分子構造 カルシウムCaは原子番号 20、原子量 40.08 のアルカリ土類金属元素である。地殻中 の存在率が 3.39%、酸素・ケイ素・アルミニウム・鉄に次いで 5 番目に多い元素とされている。 カルシウムは動植物にとって欠かせない元素で、哺乳動物ではカルシウムが骨を構成する主成分のほか、神経や細胞内の情報伝達などにも重要な役割を果たしている。
植物では カルシウムが主に細胞壁の構成成分で、細胞と細胞を強固に結びつけることで植物体を構成する。また、細胞膜の安定化、細胞内染色体の構造維持及び細胞内の新陳代謝で発生した有機酸の中和無害化にも欠かせない成分である。
植物の中にカルシウムが一番多く存在しているのは細胞壁である。細胞壁の構造はセルロースから構成する微繊維の束が、多糖類や糖タンパク質からなるマトリ ックと呼ばれる基質に埋め込まれている形である。その基質を構成する多糖類は葉や若枝、 果実など若い組織の細胞では主にペクチンで、茎などの固い木質部ではヘミセルロースが多い。
セルロースと一緒に細胞壁を構成するペクチンpectinとは、細胞壁を熱水やキレート溶液で抽出して得られる酸性多糖の総称で、ペクチン質またはペクチン性多糖とも呼ばれる。
細胞壁においてペクチンはセルロースの微繊維を固めるほか、細胞と細胞の接着にも大きな役割を果たしていると考えられている。ペクチンのカルボキシル基(COO-)がカルシウムイオンCa2+と結合してペクチン酸カルシウムとなって初めて、液体中のコロイド粒子が粘度を増加させ、流動性を失って固体状になる。
すなわち、細胞壁をコンクリート構造物と例えば、セルロースが鉄筋、ペクチン酸カルシウムがセメントと言える。
(キレート溶液は、金属イオンと強く結合する化合物・キレート剤chelate agentを含む溶液で、複数の結合部位を持ち、金属イオンと結合することで非常に安定したキレート化合物を形成する。代表的なキレート剤として、エチレンジアミン四酢酸EDTAがよく使用されている。
キレート溶液は、金属イオンの定量分析で、溶液中の金属イオン濃度を測定する。)
ペクチンは、カルシウムイオンを介して架橋を形成することで、細胞壁の力学的強度を調節し、植物の成長や形態形成に重要な役割を果たしている。これにより、細胞壁は植物個体全体に一連の編み目状に連結して、植物個体の水路がすべて完成される。これがアポプラストの構造である。
(ペクチンは植物細胞壁多糖の主成分の一つで、双子葉植物の細胞壁では多糖の約 30 % を占め、しかも、入手しやすい多糖でもあり、さまざまな用途に用いられているが、意外にも、植物内でのペクチンの構造・機能など、基礎的な知見が驚くほど乏しい。未だにペクチンの構造に不明な部分が多く、生合成機構や生理機能などの全貌がつかめていない。)
細胞壁は、セルロースを主体とする高分子多糖類から構成されているので、多くのヒドロキシ基(OH基)を持ち、水分子と水素結合を形成するため、親水性が高い。親水性の多糖類は、水を保持する力が強く、保水性を向上させる効果がある。例えば、デンプンは水に溶けやすく、食品の増粘剤として利用されることが多い。細胞壁も、いろいろな酵素やたんぱく質を分泌する。
一方、シンプラストは、生きている植物細胞の細胞質全体と、互いに細胞をつなぐ細胞質内のチャネルである原形質連絡から構成されている。原形質protoplasmは、細胞膜に囲まれた細胞の内容物全体、具体的には、核と細胞質、そして一般に細胞膜を含む。特に、原形質流動は、植物細胞内で見られる現象で、細胞内の物質や小胞体などの細胞小器官organelleが流れるように動くことを言う。この流動性により、細胞内の物質輸送や細胞の成長が可能になる。
植物の細胞外腔は、植物の細胞と細胞の間に存在する空間のことで、この空間は、細胞壁や細胞膜の外側に位置し、細胞間隙とも呼ばれる。細胞外腔には、主に空気や水が含まれており、植物のガス交換や溶存ミネラルなどの水分の移動に重要な役割を果たす。
目次へ
植物の組織の中を分子が移動する経路はいくつかも考えられるが、アポプラスト経路・シンプラスト経路・膜横断経路などが主な径路と言える。
多細胞陸上植物内の細胞は、細胞壁を充填する酸性多糖のペクチンにより細胞壁同士で接着されているから、アポプラスト経路内の水や溶解したミネラルは、細胞膜を通過せずに細胞壁外の空間に沿って移動する。シンプラスト経路では、水と溶質が細胞質に沿って移動する。この経路では、細胞から隣の細胞に移動する際に、物質は細胞膜を越える必要があり、それが原形質連絡を介しての移動となる。
細胞壁の内壁に密着して細胞膜があり、その細胞膜内が呼吸や物質代謝の場で、隣同士の細胞膜は特殊な場合を除き直接接触していない。細胞小器官の膜を含め細胞膜は、脂質二重層の構造で、その中にいろいろなタンパク質がはさまっている。前記の細胞膜連絡の特殊な場合とは、隣同士の細胞質が原形質連絡Plasmadesmata(PD)を介し細胞壁を貫通して細胞質同士がつながっていることで、細胞壁の場合と同様に、細胞質は原形質連絡を介してすべて連絡している。これがシンプラストで、個体内の物質の移動の観点から見ると、細胞壁間の移動は原形質連絡を介した細胞質の移動となる。
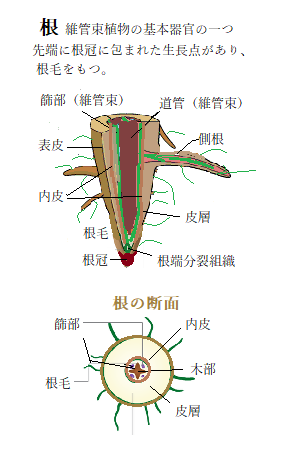 |
| 植物は土壌に根を伸ばし、土壌に含まれるさまざまな物質の中から植物の生育に必要な物質(栄養)を吸収している。ただ、土壌中には栄養素以外にも植物の生育に有害な物質や病原菌などが含まれている。これらの植物体内への侵入を防ぐため植物の根にある内皮と呼ばれる細胞層が障壁となって、その侵入を遮断する。 カスパリー線は、根の内皮細胞の間に形成される細胞壁の構造で、1865年にドイツの植物学者ロバート・カスパリーによって発見された。 根の内皮は、栄養の輸送経路である維管束を取り囲むようにして同心円状に存在し、その内皮を形成する内皮細胞には、カスパリー線とスベリン層とよばれる細胞壁を作る。カスパリー線は内皮細胞と内皮細胞の間に形成され、その隙間を埋めることによりアポプラスト輸送の防御障壁として機能する。スベリン層は、内皮細胞全体を覆うように形成され、細胞外から内皮細胞内に有害物質が入ること事態を防ぐ。 近年、研究グループは低カルシウム条件で生育が阻害されたシロイヌナズナの変異型株を単離し、この変異株や他のカスパリー線変異株を解析比較することにより、側根発生部位における障壁の存在と、その障壁がスベリン層により形成されていることが解明された。 この発見の端緒となった変異株は、低カルシウム条件下で生育阻害に加え、地上部カルシウム濃度の低下、カスパリー線の形成異常、スベリンの過剰蓄積が起きていた。この中の一つの現象であるスベリンの過剰蓄積が植物の物質輸送に与える影響を調べると、その観察の過程で、側根発生部位ではアポプラスト輸送が行われていないことと、その輸送の障壁としてスベリン層が機能していることが示された。 スベリン層の形成を観察すると、側根が発生するに従って、側根発生部位で蓄積することが確認できた。その結果と、側根の発生過程から、スベリンは側根と内皮細胞の間に生じた隙間を埋めることにより、アポプラスト障壁を形成していることを明らかになった。 これらの構造は、植物が土壌から栄養を吸収する過程で、極めて重要な役割を果たしている。 |
膜横断経路は、物質が細胞膜を越えて移動する経路で、細胞から細胞へ移動する際に細胞膜を通過する必要がある。
光合成で炭素1モルを同化すると、数百モルの水が植物体を通過すると言う。このうちの何割かの水は、シンプラスト経由で移動するが、水の輸送が植物細胞にとって最も多量の膜輸送であることがわかる。この膜輸送を担う実体として、細胞膜にある水を特異的に通す水チャネルのタンパク質、アクアポリンaquaporinである。
このタンパク質は、水分子を選択的に通過させることで、細胞内外の水分バランスを調整する。アクアポリンは、植物・動物・微生物などさまざまな生物に存在し、特に腎臓や赤血球などで重要な役割を果たしている。例えば、腎臓では尿の濃縮や再吸収に関与し、体内の水分を効率的に管理する。
生体膜においては、脂質二重層を介した水の動きは拡散として、また水チャネルを介した輸送は体積流として解釈されることが多いが、実際には拡散も体積流も同時に起きている。現実的には全体としての水の流速、すなわち単位時間あたりに流れる水の量、つまり「流量(J)」は、単位時間あたりに流れる水の体積で表す。一般的には、立方メートル毎秒(m³/s)で表される。
例えば、水が1秒間に1m進む場合、その流量は1立方メートル毎秒1(m³/s)になる。
それは、 J (流量)=G(水の透過性)×V(駆動力) という式で考えていくことができる。
単位膜面積当たりの水透過率(Lp)は、植物種や細胞によってさまざまな値が発現される。 植物が正常に生育している場合、根では能動的なイオン吸収などによって浸透圧を高めて土壌よりも低い水ポテンシャルを保っている。道管では根より低い水ポテンシャルになっており、葉では同化産物や無機イオンの蓄積および蒸散によって、さらに大きな負の水ポテンシャルが生じている。このような水ポテンシャル勾配が有効に形成されているので、根での水吸収と地上部への水輸送が滞りなく行われている。
土壌の水ポテンシャルが低下し始めて、土壌と根の間で水ポテンシャル差が小さくなった場合、アクアポリン(タンパク質)を増加させて水透過性を増大させることが、水吸収を確保するために有効である。しかしストレスがより強力であれば、水は逆に根細胞から土壌に向かって動き、植物は脱水される。
この場合、アクアポリンの数を減らすか、あるいは細胞間連絡を閉鎖するかして一時的にでも根の水透過を下げて脱水を回避するのが有効である。その後、ストレス環境下に適応するためには、無機イオンの蓄積や浸透圧調節物質の合成などによって植物体内の水ポテンシャルを下げることによって植物にとって望ましい方向の駆動力を回復させる。その上で、アクアポリンの数を増やすなどして水透過性を上げて吸水と成長を持続させる。
ただ実際の水ストレス‐植物相互作用のシステムはもっと複雑で、葉の水ポテンシャルが低下すると、それを感受して気孔を閉じ始めると、蒸散が減り木部や根の水ポテンシャルも変化し、植物ホルモンABAやカルシウムイオンなども関与してくる。水チャネルだけで植物の水分生理が完結するのではなく、組織や細胞が互いに影響し合いながら植物体全体で働く適応機構の一部として水チャネルは機能し、制御されている。
根に入った水が移動する経路としては、シンプラスト(細胞間連絡によってつながっている細胞質の連続構造)、アポプラスト(細胞間隙による水流)の2つが古くから提唱されてきた。現在では、どちらの経路も水輸送においては機能していて、環境条件や生理条件で2つの経路を通る割合が変わるものと理解されている。また複数の細胞を横切って動く細胞横断経路transcellular pathも考えられているが、通常、実験的にはシンプラスト経由の水の移動と区別して測定することができないので、2つを合わせてcell-to-cell pathと呼ぶこともある。
シンプラストはもともと細胞質の連続体と考えられていたが、現在では液胞横断経路transvacuolar pathとも呼ばれる、細胞質から液胞膜を横切って水が流れる経路も広い意味でのシンプラストに含めて考え、こちらの方が寧ろシンプラストにおける主要な流路である可能性が高いと評価されている。これは「単位面積当たりの水透過率Lp」が、細胞膜に対して液胞膜の方が数倍から数十倍高いことが一般的であるとわかってきたためで、液胞が水の移動の際の抵抗にならないのなら、通常細胞体積の90%以上を占める液胞を横切って水は動くと考えられるからである。
仮に液胞膜のLpが細胞膜のLpよりも小さいとすると、細胞外の浸透圧が急に増加したとき、細胞膜を通って細胞質の水が急速に失われ、細胞質の体積が急変する。液胞膜が細胞膜より高いLpを持つことは、そのようなショックを防ぐのに役立っている。
主としてアポプラスト経由で根組織内を移動してきた水も、成熟した組織ではカスパリー線が存在するために一度はシンプラストに入ると考えられている。カスパリー線は、植物の根の内皮細胞の細胞壁に存在する脂質からなる帯状の構造で、これにより水や溶存物質の受動的な流動を制限し、中心柱への流入を管理する。カスパリー線はリグニンやスベリンsuberinから成り、根の内皮細胞の細胞壁に形成されている。これにより、植物は不要な物質が体内に入るのを防ぎ、必要な栄養素を効率的に吸収する。したがって最低一度は細胞膜にある水チャネルを介した膜輸送が行われることになる。
近年、栄養吸収に異常があるシロイヌナズナの変異型株の解析により、根の成長過程において、主根から分岐してくる側根の発生部位における障壁が、スベリンと呼ばれる脂質層により形成されていることを明らかになった。本成果により、植物の根がその成長に応じて物質輸送の障壁を柔軟に形成し、栄養吸収を効率良く行う仕組みの一端を明らかにされた。スベリンと呼ばれる脂質の一種が、物質輸送における障壁として機能し、カスパリー線と異なる二つの細胞壁構造が強調して、アポプラスト障壁を形成していた。
植物は土壌に根を伸ばして栄養や水を吸収するが、不必要な物質は回避する。そのためには、植物体内と体外を分ける障壁が必要であるはずだが、この障壁が植物の成長過程において維持される機構が解明されていなかった。今回、異常があるシロイヌナズナの変異型株を解析することにより、根の成長過程において、主根から分岐する側根の発生部位における障壁が、スベリン層により形成されていることが明らかにされた。この時に働く細胞膜型水チャネルの性質や制御は、次に道管に移行する際には、シンプラストの水は再び細胞膜水チャネル経由で出て行く必要がある。この時の水チャネルは、シンプラストに水が入る時に働くものと別のアイソフォームisoformがあるかもしれないことが想定された。
アイソフォームisoformは、同一の遺伝子から生成される異なるタイプのタンパク質を指す。これらは選択的スプライシングalternative splicingや異なるプロモーターの利用などによって生じる。この場合のSplicingには、「解いて組み継ぎする、 より継ぎする、 継ぎ合わせる」の意味で使われる。アイソフォームは、同じ遺伝子から異なる機能や特性を持つタンパク質を生み出すことで、生物の多様性と適応能力を高める。
選択的スプライシングは、真核生物の遺伝子発現の過程で、mRNA前駆体のコード領域に含まれる遺伝子と、異なる遺伝子が成熟mRNAにコード化される仕組みであるが、このプロセスにより、1つの遺伝子から複数の異なるタンパク質が作られる可能が高まり、タンパク質の多様性が増して、生物が環境に適応しやすくなる。
木部に入った水の一部は途中で篩管に移行するが、道管の中で気泡が発生すると、水の流れが遮断される。これは低温条件下で起こりやすいが、一度これが発生すると、木部の水透過性が大きく低下し、枯死にいたる。これを避けながら何本もの道管間を移行して地上部に向かい、葉肉組織内の細胞の空間である細胞間隙から蒸気とし放出する。その細胞間隙の出口が気孔である。気孔が充分開き、かつ細胞間隙内‐大気間の水蒸気圧差が充分あれば、大量の水がここから蒸散される。
気孔の開閉が植物体内の水分状態や光環境などによって制御されていることはよく知られている。気孔開閉の実体は孔辺細胞の膨圧変化による。しかしこの膨圧運動の時に同時に働いているはずの孔辺細胞の水チャネルについては、まだ研究が進んでいない。
葉に到達した水の一部は葉肉細胞などにも水チャネル経由で取り込まれる。地上部では、根で働く水チャネルとは別のアイソフォームが発現、機能していることが想定される。
目次へ
 |
| 動物や植物などの真核生物の細胞を構成する細胞小器官の一つである小胞体は、一重の生体膜に囲まれた板状あるいは網状の膜系で、核膜の外膜とつながっるへん平な袋状の膜が幾重にも折り重なった構造をした細胞小器官である。 主に、細胞内において物質の合成と輸送を担う。 粗面小胞体と滑面小胞体と呼ばれる二つの種類の細胞小器官に分類される。 粗面小胞体は、その名の通り、膜構造の表面に多数の釘や鋲が打たれたような形状をしている。そうした粗面にみられるゴツゴツした部分の一つ一つが、リボソームribosomeと呼ばれる粒子状の複雑な構造体である。 リボソームで合成されたタンパク質は粗面小胞体内腔に取り込まれ、そこで適切に折りたたまれ修飾される。その後、ゴルジ装置へと輸送され、さらに修飾や梱包が行われる。最終的に、これらのタンパク質は細胞膜や他の小器官へと運ばれたり、細胞外へ分泌されたりする。 滑面小胞体の表面にはリボソームがなく、滑らかな外観をしている。主に中性脂肪・コレステロール・ステロイドホルモンといった細胞の生命活動に関わるその他の様々な物質の合成が行われている。また、カルシウムイオンの貯蔵庫として機能し、カルシウムの放出と取り込みを調節する。 小胞体は、細胞の正常な機能を維持するために不可欠な役割を担う。 |
トランスポーターは、特定の物質を細胞膜を通して移動させるタンパク質で、エネルギーを使わずに濃度勾配で物質を移動させる「受動輸送」と、エネルギーを使って濃度勾配に逆らって物質を移動させる「能動輸送」の2種類がある。
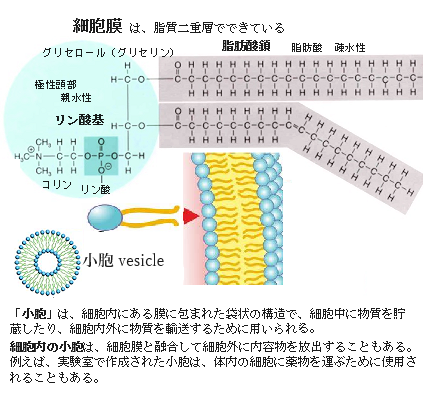
細胞膜を含め、全ての生体膜の基本構造は、グリセロリン脂質であるホスファチジルコリンやホスファチジルエタノールアミンなどの脂肪酸(炭化水素鎖にカルボキシ基R-COOHを有する1価のカルボン酸)どうしが向かい合って形成される脂質二重層である。
ホスファチジルコリンPhosphatidylcholine (PC)は、細胞膜の主要な構成成分で、その基本構造のグリセロリン脂質の親水部では、リン酸のヒドロキシル基(構造式が 炭化水素Rと結合する「-OH」基で表される1価の官能基)とコリンのヒドロキシル基が反応して形成される(コリンとリン酸のエステル結合と言う)。また疎水部の脂肪酸鎖がグリセロリン骨格にエステル結合した構造をしている。その脂肪酸の組み合わせが多数あることからホスファチジルコリンに属するリン脂質は複数存在している。それが「リン脂質」の総称で呼ばれる。 ヒドロキシル基(−OH)相互の脱水結合は、一般的に「脱水縮合」と呼ばれる反応で、2つのヒドロキシル基が結合し、水分子H₂Oが脱離して新しい結合が形成さる。この構造により、ホスファチジルコリンは両親媒性、つまり親水性と疎水性の両方の機能を発揮し、細胞膜の形成や機構で重要な役割を果たす。
物質が細胞膜を介して反対側に移動するためには膜を横切らなければならないが、その際に疎水性の高い脂肪酸部分が脂質バリアーとして働く。このため、脂溶性の高い物質は膜を透過しやすく、逆に水溶性の高い物質は膜を透過できない。これによって細胞外の物質が無制限に細胞内に侵入するのを防ぎ、また細胞内の必要な物質が細胞外へと散逸するのを防いでいる。
このような細胞膜のバリアー機能は、細胞が生きていく上で不可欠であるが、単なるバリアーとして働くだけでは、能動的な恒常性は維持できない。細胞膜は物質移動のバリアーであるとともに、細胞にとって必要な物質の取り込み、不要な物質の排出の窓口という、いわば相反する機能を同時に活用している。そのために発達してきたのが細胞膜に存在する各種のトランスポーターである。トランスポーターは膜貫通型のタンパク質であり、酵素やレセプター(受容体)など他のタンパク質とともに生体膜を構成するが、外部からのシグナル(情報伝達物質やホルモンなど)を受け取る機能も発揮する。全てではないが多くのタンパク質は、脂質層の中を側方拡散することができる。つまり脂質分子は二重膜中を自由に水平移動できる。同一層内での移動は極めて速く、大型の細菌の細胞膜の端から端まで約2μmを1秒で移動できる。
 |
| オーキシンは植物ホルモンの一種で、植物の成長を調整する役割を持っている。除草剤として使用されるオーキシン化合物は、特に双子葉植物(広葉雑草)に対して効果的で、異常成長を引き起こして枯死させる作用がある。 イネ科植物(単子葉植物)にはほとんど影響を与えないため、水田などで雑草を除去するために使用される。 |
その長距離移動は維管束系を介するが、維管束系から植物ホルモンの隣接細胞内への移行は、トランスポーターやパーミアーゼなどの膜貫通タンパク質が関与している。これらのタンパク質は、ホルモンを細胞膜を通して輸送する役割を担っている。
例えば、アブシジン酸(ABA)という植物ホルモンは、ABCトランスポーターと呼ばれる輸送因子によって細胞内から細胞外へと運ばれる。また、オーキシンのような植物ホルモンも、特定のトランスポーターによって細胞内外を移動する。
オーキシンは、茎の先端(頂芽)で合成され、茎を下に向かって運ばれ、細胞の伸長、根の形成、葉の向き制御などに関与する。その細胞の分裂・分化・伸長を介して、根や葉などの器官発生、光・重力・水分の方向に応じた伸長、果実の成熟など、植物の発生と成長のほぼ全てに関与する重要な因子と言える。特にクライマクテリック型果実、つまりその成熟時に呼吸量が著しく増大する現象が見られるリンゴ・モモ・アボカド・パパイヤ・ナシ・バナナ・アンズ・メロン・マンゴー・トマトなど、この型の果物の成熟過程において植物ホルモンであるエチレンの濃度が通常の1,000倍以上も劇的に増加する。
主なクライマクテリック型果実には、オーキシンの増加とエチレンの生成との関連が見られ、一般的に、クライマクテリック型の果実では、成熟が進むとエチレンの生成量が増加し、果皮の色や果肉の食感、風味などの変化が促進され、また収穫後の貯蔵中に追熟を行うことができる特性がある。また、受粉によって、めしべで生成されるオーキシンとジベレリンが果実形成を促進すると考えられている。
高濃度のオーキシンが存在すると、細胞分裂が促進され、シュート(茎と葉)の形成が生じる。また除草剤や挿し木の発根促進、果実の品質管理など農作業にも広く利用されている。
目次へ
農薬のような脂溶性低分子物質は、細胞膜の脂質二重層lipid bilayerを通過しやすい特性を持っている。脂質二重層は、細胞膜の基本構造を成す重要な要素、これは、親水性(親水基)と疎水性(疎水基)の両方を持つリン脂質分子が二重に並んで形成される膜である。
親水性部分のリン脂質分子の頭部は水に親しみやすい親水性を持ち、細胞膜の外側と内側に向いている。疎水性部分のリン脂質分子の尾部は水を嫌う疎水性を持ち、膜の内側に向かって並んでいる。
この二重層構造により、細胞膜は細胞内外の環境を隔て、特定の物質のみを通過させる選択的透過性を持つ。具体的には、分子が小さく、脂溶性が高いため水に溶けにくく、電荷を持たない(疎水性と非極性を有する)物質ほど、細胞膜を通過しやすく、これらの物質は濃度勾配に従って細胞膜を直接通過し、細胞内に吸収される。
この特性により、脂溶性低分子物質は植物の細胞膜を容易に通過し、効果的に作用することができる。多くの農薬はこの特性を利用して、植物の内部に浸透し、害虫や病原菌を駆逐制御する。
水溶性の低分子物質や無機イオン類は、細胞膜にある膜貫通型のタンパク質であるトランスポーターと細胞膜を貫通して形成されたチャンネルにより、細胞質と細胞壁の間の移動を行う。
 |
| アクアポリンは、細胞膜に存在するタンパク質で、水分子を選択的に通過させる「水チャネル」として機能する。これにより、細胞内外の水の移動を効率的に行う。
アクアポリンは、植物・動物・微生物など多くの生物に存在し、特に腎臓や赤血球などで重要な役割を果たしている。例えば、腎臓では尿の濃縮や再吸収に関与し、体内の水分バランスを保つための主要なタンパク質の一つとして存在する。 |
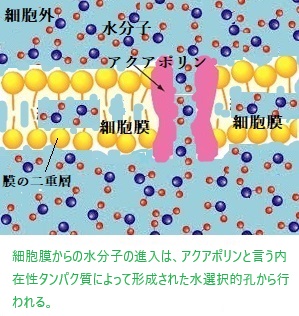
アクアポリンAquaporinと言う細胞膜に局在する膜貫通型タンパク質が「チャネル」を通る水の高速輸送を担う。「チャネル」とは、細胞膜に存在する小さなイオンを通す孔のことで、これらのチャネルでは、リン脂質二重層膜がイオンを通すための水で満たされた通路を形成している。水チャネルとも呼ばれる。アクアポリンは水分子を選択的に透過させ、イオンや他の物質は通さない特性があり、植物や動物の細胞において、水分子の輸送に不可欠な役割を果たす。物質的な孔は、いくつかのサブユニットの集合体によって構成され、親水性アミノ酸残基が並んでいる。この孔の狭い領域は「選択性フィルタ」と呼ばれ、どのイオンが通過できるかを決定する。
細胞膜は細胞と外界を隔て、内部の一定の環境に保つために必須の構造で、核や小胞体、ミトコンドリアなどの細胞小器官も膜を持ち、膜を挟んだ特異的な輸送により内部を適切な環境に保っている。脂質は細胞膜の全重量の半分に過ぎず、残りの半分は膜に突き刺さった膜タンパクで構成されている。
膜タンパクは、膜を横切る分子の輸送やシグナル伝達、ATP合成などの膜を足場とする化学反応を触媒する。水溶性の低分子物質やイオンは、膜貫通型タンパク質(トランスポーター)やチャネルを通じて細胞膜を通る。例えば、水はアクアポリンという細胞膜に局在するタンパクがチャンネルを通る。このように、細胞膜の構造と膜タンパクの協力によって、物質の選択的な輸送が実現されている。
チャンネルの太さは細胞質側の必要度によって変化し、水の濃度、つまり浸透圧で物理的に移動する。外から与えた肥料塩は、アポプラストからそれぞれ特異なトランスポーターで細胞質へ取り込まれる。トランスポーターの多くはATPをエネルギーとして利用する。
この膜貫通経路では、溶解したミネラルや水が細胞から細胞へ移動する際に、細胞壁を越えてある細胞から次の細胞へ流入する。これらの経路は相互に排他的なものではなく、溶質によっては程度の差こそあれ、複数の経路の利用が重なる。
植物の細胞外腔(さいぼうがいこう)は、植物の細胞と細胞の間に存在する空間のことを言う。この空間は、細胞壁や細胞膜の外側に位置し、細胞間隙とも呼ばれる。細胞外腔には、主に空気や水が含まれており、植物のガス交換や水分の移動に重要な役割を果たす。
これ以外に、維管束系による長距離移動があるが、もう1つの可能性のある経路が液胞経路で、この経路はほとんどが水分子の移動に限定される。植物細胞では、浸透圧が水の移動を促進する。液胞は植物細胞内で大きな役割を果たし、糖や無機イオンの集積によって浸透圧差が生じる。この浸透圧差により、水が液胞内に移動する。さらに、液胞の輸送は、2つのプロトンポンプ(ATPaseとPPase)によって促進され、溶質の取り込みに活力を与えている。また、液胞には特殊な輸送タンパク質であるアクアポリンが存在し、水やグルコース、スクロースなどの溶質の輸送に関与している。
液胞膜には水チャネルが多く存在し、水の移動をさらに促進する。液胞が膨らむことで細胞の膨圧が生じ、これが細胞の形状維持や成長に寄与する。
「アクアポリン」(aqua=水、porin=孔の意味、水チャネル)は生体膜にある主要なタンパク質のひとつで、水の透過孔(水チャネル)を形成し、植物体内の水の流れに重要な役割を果たしている。アクアポリンは細菌から動植物まで普遍的に存在しており、人間の体内では腎臓などで重要な役割を担っている。
 植物では、動物以上に多様なアクアポリン分子種が備わっている。近年、二酸化炭素などの低分子化合物を透過させるアクアポリン分子種も発見され、低分子化合物の輸送体としての生理機能が注目されている。 いくつかのアクアポリン分子種は原形質膜において細胞外から細胞内への二酸化炭素の取り込み、別種のアクアポリンでは細胞内で葉緑体への二酸化炭素炭素の取り込みに関与して、葉緑体内のルビスコrubiscoに二酸化炭素をスムーズに供給することで光合成機能を向上させている。
植物では、動物以上に多様なアクアポリン分子種が備わっている。近年、二酸化炭素などの低分子化合物を透過させるアクアポリン分子種も発見され、低分子化合物の輸送体としての生理機能が注目されている。 いくつかのアクアポリン分子種は原形質膜において細胞外から細胞内への二酸化炭素の取り込み、別種のアクアポリンでは細胞内で葉緑体への二酸化炭素炭素の取り込みに関与して、葉緑体内のルビスコrubiscoに二酸化炭素をスムーズに供給することで光合成機能を向上させている。アクアポリンの水分子輸送速度は非常に高い。各種イオンチャネル、イオントランスポータなどの輸送体の中でも最速といわれ、条件が良ければ1秒間に10億以上の水分子を輸送する。その一方では、水分子H2Oは透過させても、水素イオンH+は通さない特殊な選択機能を有す分子構造を持ち、生体内の体質を保全している。
分子種によってはグリセロール・尿素・アンモニア・二酸化炭素・酸素・過酸化水素・ホウ酸・ケイ酸・亜ヒ酸・アルミニウムなどの低分子は透過する。植物のアクアポリンは、シロイネナズナでは35種、イネでは33種の分子種が存在し、大きなタンパク質ファミリーを形成している。しかも、輸送基質特性はアクアポリン分子種ごとに異なり、水以外の輸送基質それぞれに選択される分子機構については未だ分析中と言う。
アクアポリンの活性は、遺伝子やチャネルタンパク質の発現量によって調節されるだけでなく、外界や生体内の環境変化に応じて素早く活性を変化させている。 開花には花弁の細胞が吸水して伸長することが必須で、この時にアクアポリンが関与している。
開花とチャネルタンパク質のリン酸化によるチャネルの開閉には密接な関係がある。 チャネルタンパク質のリン酸化は、植物の細胞膜に存在するチャネルの開閉を制御する重要なメカニズムの一つで、リン酸化とは、タンパク質にリン酸基が付加することで、その構造や機能が変化する現象を言う。これにより、チャネルが開いたり閉じたりすることが可能になる。
開花においては、特定のチャネルタンパク質がリン酸化されることで、細胞内外の水やイオンの移動が調節され、花弁の細胞が膨張して開花が促進される。水チャネルタンパク質アクアポリンは、リン酸化によって活性化され、開花に必要な水の移動を助けている。このように、チャネルタンパク質のリン酸化は、植物の生理機能において非常に重要な役割を果たしており、開花のタイミングや効率にも大きな影響を与えている。
アクアポリンの活性調節は、低酸素とか浸透圧ストレス(塩ストレスや乾燥によって起る)などの環境下での根の水透過制御に直接関与していることも示されている。
アクアポリンの活性調節では、植物が冠水されたり、土壌中の酸素濃度が低下したりする低酸素ストレスの環境下や、乾燥や塩分濃度の高い環境などで細胞から水分が失われる浸透圧ストレスの環境下では、アクアポリンが根における水の透過制御に直接関与していることも示されている。
低酸素状態では、植物細胞内のpHが低下し酸性環境に傾く。酵素は特定のpH範囲で活発に働くため、pHの変化は代謝プロセスに影響を与える。また酸性環境は細胞膜の構造や機能に影響を与え、イオンや分子の輸送が妨げられる可能性もある。また植物の成長を抑制することがある。
例えば、根の成長が遅くなり、栄養吸収が困難になる。植物はこの酸性環境に適応しようとする一連の反応を示す。例えば、レタスなどの植物は酸性環境下で根毛を発達させ、水や栄養の吸収をより効率化することで生育阻害を回避する。
浸透圧調節の主な方法としては、細胞内に溶質、例えばカリウムイオンK+や糖類を蓄積することで、細胞の膨圧を維持しようとしする。植物の細胞のpHはだいたい8くらい、その細胞のpHが低下すると、細胞膜にはATPによって駆動される水素イオンH+のポンプ(ATP-プロトンポン)があり、このポンプによって細胞から水素イオンH+がひとつ放出されると、同じ電荷をもったカリウムイオンK+がカリウムイオンチャネルを通って細胞内に取り込まれる。カリウムイオンチャネルはタンパク質から出来た細胞膜などに存在するため、特異的にカリウムイオンを通す。しかし、カルシウムイオンCa2+やマグネシウムイオンMg2+などの二価のアルカリ土類金属はほとんど通過しない。この浸透圧ストレスに対する応答は、乾燥ストレスに対する応答と基本的に共通しており、植物は環境の変化に応じて様々な生理的変化を起こす。
目次へ