更新世後期の時代 『第四紀の大量絶滅時代』
|
 2027年4月9日,高島城の桜庭園 |
ケナガマンモスMammuthus primigenius(primigenius=原初の)は新生代の約500万年前(鮮新世)から完新世の約4,000年前まで生息していた。ケナガマンモスMammuthus primigeniusは、更新世(氷河期)の終わり頃のヤンガードリアス期((約12,900~11,700年前))の急激な寒冷化による環境変化にも適応して生き延びていた。
ケナガマンモスは、マンモス属の中でも特に寒冷地に順応した種で、厚い毛皮や脂肪層を持ち、寒冷な環境に適応し変異し進化してきた。一般的に「マンモス」と呼ばれることが多い。マンモス属には他にも異なる種が存在し、それぞれの生息地に適応して特徴が異なる。ケナガマンモスは7種類ほど確認されているマンモスの仲間のひとつでアジアやヨーロッパ、北アメリカと広い範囲に生息していた。マンモスの中では体の大きさは平均的で、現在、アフリカに生息しているアフリカゾウよりもやや小型で、その特徴的な大きくカーブした長い牙は武器ではなく草の上に積もった雪を払うために用いられたと考えられており、主食はマツやヤナギ、カバなどの樹木の表皮や葉や枝だったとされている。日本国内においても北海道の襟裳岬や夕張市などでマンモスのものと思われる臼歯が見つかっている事からツンドラ化した北海道にも生息していたことになる。
それらの化石から、北海道の地理的条件や氷河期の海面低下によって、マンモスがシベリアからサハリンを経由して北海道に渡ってきたと考えられている。この巨大な哺乳類は、氷河期の厳しい寒冷地に適応しており、厚い毛皮や脂肪層で寒さをしのいでいた。日本列島では、当時の気候や地形が彼らの生息に適していた地域が限られていたため、主に北海道に集中していた。寒冷地に適応し、厚い毛皮や脂肪層を持っていた。草食性で、草や低木を主食としていた。
寒冷の気候にも適応していたケナガマンモスの絶滅の原因については、急激な気候の変化によって食料としていた植物が激減してしまったとする説や、現代のアフリカゾウのように人の手による過剰な狩りを原因とする説、人間が持ち込んだ家畜から移った伝染病が原因であるとする説など様々なものが想定されている。寒さの厳しい地域に生息していた事が幸いし、化石だけでなく肉や毛の残った個体が多く見つかっている事もケナガマンモスの特徴となっている。最後のケナガマンモスの生存記録は、北極海の東シベリア海とチュクチ海との間にあるロシア領のウランゲリ島で確認されている。
ウランゲリ島でケナガマンモスが長く生存できた理由としては、孤立した環境が挙げられる。この島は海面上昇によって大陸から切り離され、外部からの捕食者や人間の狩猟圧が少なかったため、比較的安全な生息地であった。それが絶滅の原因でにもなった。ウランゲリ島のマンモスは地理的に隔離され、近親交配によって遺伝的多様性を弱められ、そのため極端な突然の気象現象に適応することができず絶滅を招いたようだ。この島では、約4,000年前までケナガマンモスが生き延びていた。最後のマンモスが死んだ時、ギザの大ピラミッドは既にエジプトに建設されていた。
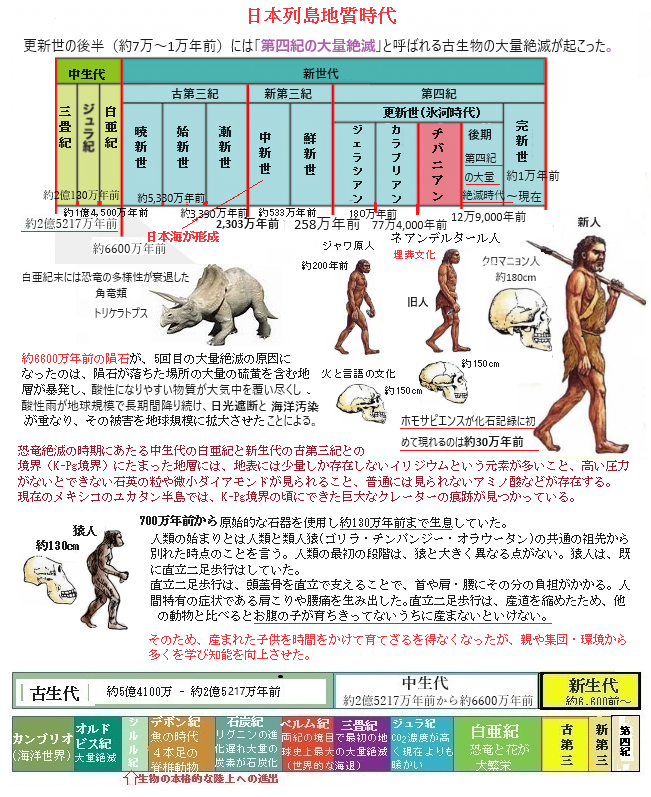 ケナガマンモスは、主にユーラシア大陸の広範囲、シベリアからヨーロッパや北アメリカ大陸に生息していた。日本列島では、かつては北海道の一部にも生息していた。北海道では、ケナガマンモスの化石がいくつか発見されている。例えば、北広島市周辺では更新世後期(第四紀の大量絶滅時代)の約4万5000年前のケナガマンモスの化石が確認されており、ナウマンゾウと共存していた可能性があるとされている。また、夕張市やえりも町、根室海峡の海底でもケナガマンモスの臼歯化石が発見されており、これらの化石の年代は約4万5,000年から2万年前の間とされている。
ケナガマンモスは、主にユーラシア大陸の広範囲、シベリアからヨーロッパや北アメリカ大陸に生息していた。日本列島では、かつては北海道の一部にも生息していた。北海道では、ケナガマンモスの化石がいくつか発見されている。例えば、北広島市周辺では更新世後期(第四紀の大量絶滅時代)の約4万5000年前のケナガマンモスの化石が確認されており、ナウマンゾウと共存していた可能性があるとされている。また、夕張市やえりも町、根室海峡の海底でもケナガマンモスの臼歯化石が発見されており、これらの化石の年代は約4万5,000年から2万年前の間とされている。ナウマンゾウは更新世(氷河時代)チバニアン期の約40万年前から更新世後期の2万年前まで、主に東アジアを中心に生息していた。ナウマンゾウは比較的温暖な気候に、マンモスは寒冷な気候に適応した環境適応の違いが、体の特徴の違いにも表れている。マンモスの体毛は非常に長く、最長で1mに達するものもあり、厳しい寒冷地での生活に適応し変異.した。
ナウマンゾウの体毛はマンモスほど長くはない。これは、ナウマンゾウがマンモスほど寒冷な環境に適応する必要がなかったことを示す。しかし、現在生きているアジアゾウよりは長い体毛を持っていた。これは、ナウマンゾウが生きていた最終氷期という、現代よりも寒冷な時代に適応した結果だと言える。
マンモスとナウマンゾウの牙には、顕著な違いがあり、マンモスの牙は極端に湾曲し、長いもので5mを超えるものもある。この特徴的な形状は、マンモスの生活様式と密接に関連していた。長く湾曲した牙は、雪を掻き分けて地面の植物を探したり、木の皮を剥いだりするのに適していた。また他のマンモスとの争いや捕食者からの防御にも優れている。ナウマンゾウの牙はそれほど長くなく、湾曲も緩やかであった。これは、ナウマンゾウがマンモスほど厳しい環境で生活する必要がなかったことを示唆している。ナウマンゾウの牙は、現代のアジアゾウに近い形状だったと考えられており、主に木の枝を折ったり、樹皮を剥いだりするのに使われていたと推測されている。
ケナガマンモス以外のマンモスは、アフリカ・アジア・ヨーロッパ、そして北アメリカなど広範囲に生息していた。例えば、コロンビアマンモスは北アメリカの草原や森林地帯に適応していた大型種で、比較的毛が少ないのが特徴であった。また、クレタ島に生息していたクレタマンモスMammuthus creticusは、非常に小型で、肩高が約1m程度と推定されており、島嶼環境に適応して小型化した「島嶼矮化」の例とされている。 クレタマンモスは約350万年前までギリシャのクレタ島に生息していたとされている。
最古のマンモスは約600万年前のアフリカにルーツがあり、その後ユーラシアや北アメリカに分布を広げ、その過程でそれぞれが進化した。絶滅の要因としては、気候変動や人類の狩猟、伝染病などが挙げられている。
コロンビアマンモス Mammuthus columbi の名前にある「コロンビア」は、南米コロンビアでもサウスカロライナ州の州都コロンビアでもなく、探検家クリストファー・コロンブスに由来する。このマンモスは北アメリカから中米にかけて生息していた種である。 祖先にあたる、更新世の前期カラブリアンの約180万年前~中期チバニアンの約20万年前にかけて、主にユーラシア大陸北部に生息していたマンモスの一種トロゴンテリーゾウMammuthus trogontheriiには劣るが、体高4m・推定体重10tと、平均的なアフリカゾウを凌ぐ巨体であった。牙も発達しており、平均的な長さは3.5m、最大で4.9mにも達した。
トロゴンテリーゾウは、寒冷な気候に適応し、広大な草原やステップ地帯を中心に生息していた。実は、日本でも滋賀県、新潟県の十日町市や長岡市、津南町など、また千葉県でも化石が発見されており、ムカシマンモスとして知られる個体もこの種に含まれると考えられている。これらの地域は化石の集中産出地として知られている。 これらの化石は、トロゴンテリーゾウの生息域や移動経路を解明する上で重要な資料とされている。 このゾウは、肩高が約4.5m、体重が20t近くにも達する非常に大型の哺乳類で、ゾウ科の中でも最大級の種の一つであった。寒冷な環境に適応するため、長い体毛を持っていたと考えられているが、後のケナガマンモスほどではなかったようだ。
フランスのコタンタン半島西方沖合い、英国海峡に浮かぶ島々であるチャンネル諸島Channel Islandsから発見されたピグミーマンモスは、島嶼性矮小化した種とされており、最大でも体高2m・推定体重1.3t程にしかならなかった。 他のマンモスと同じく主に草を食べていたようだが、アメリカに進出しただけあってサボテンも食べていたことが糞化石から明らかとなっている。
北米大陸にはマンモスを代表する種であるケナガマンモスも棲息していたが、こちらはカナダなどより北方の地域に棲息し、南方に棲息したコロンビアマンモスとはすみ分けていたとされている。とは言え、一部の地域では重複して生息しており、その結果両種の間の交雑個体もいたらしく、かつて独立種とされていたジェファーソンマンモスがこれに相当する。ケナガマンモスほど毛深くはなかったが、本種も体毛を生やしていたと判明している。 多くの大型哺乳類と同様に更新世末期に絶滅したが、理由としては気候変動の他、ネイティブアメリカンの祖先とされるパレオ・インディアンの狩猟も影響したと考えられている。その証拠にコロンビアマンモスの化石には、石器による傷跡や装飾品に加工されたものも確認されている。 但し、こうした狩猟の痕跡の発見例は非常に少なく、それよりも末期の化石には病変の痕跡の方が多数確認されており、人類を含めユーラシア大陸から進出した哺乳類由来の伝染病が大きく影響していたとする説が有力になっている。
コロンビアマンモスの化石で見つかる、いくつかの病理学的な痕跡は、骨であれば、関節炎や骨の変形が確認されている。これらは、マンモスが高齢になるにつれて発生した可能性があり、特に体重が重いことが関節に負担をかけたと考えられている。 また、牙の摩耗や損傷も見られることがあり、これは食事や環境との相互作用によるものと推測されている。さらに、化石の中には、感染症や栄養不足の痕跡が見て取れるが、それららの詳細については、現在も研究が進行中であると言う。 こうした病理学的な痕跡は、マンモスの生活環境や健康状態を理解する上で重要な手がかりとなる。さらに詳しい調査結果の発表が待たれる!
目次へ
 スリランカゾウの子供 スリランカゾウの子供 |
 タイゾウは3,000頭以上飼育され観光業に従事する。 |
| ナウマンゾウやケナガマンモスはアジアゾウと近縁関係にある。これらはすべてゾウ科Elephantidaeに属する動物で、共通の祖先を持っている。 ナウマンゾウは、アジアゾウに近縁なパレオロクソドン属Palaeoloxodonに属しており、特にアジアゾウと遺伝的に近いとされている。 ケナガマンモスはマンモス属Mammuthusに属し、アジアゾウと非常に近い遺伝的関係を持っている。 実際、アジアゾウとケナガマンモスの間の遺伝的差異は、アジアゾウとアフリカゾウの間の差異よりも小さいとされている。 これらの動物は、それぞれ異なる環境に適応して進化したが、遺伝的なつながりが深いことが研究によって明らかになっている。 |
ナウマンゾウは、寒冷な気候に適応するために皮下脂肪が発達し、全身が体毛で覆われていたと考えられている。肩高は約2.5~3mで、現生のインドゾウ(アジアゾウ属)よりもやや小型であった。ナウマンゾウの最大の特徴が口元の牙であるが、長さが2.4m以上、直径も20cm以上まで成長する。主に針葉樹と広葉樹の混成林を生息地としていた。
ナウマンゾウは、北海道から九州まで、日本の様々な地域で多くの化石が出土している。ナウマンゾウの化石と共に、旧石器時代の人類が使用したと考えられる石器が発見されている。特に長野県の野尻湖周辺では、ナウマンゾウの化石とともに槍の穂先として使用された可能性のある石器や、解体作業に使われたとみられるスクレイパー(皮はぎ用の道具)などと共伴している。 これらの発見状態から、旧石器時代の人々がナウマンゾウを狩猟し、その肉や皮を利用するのはもとより、化石の散らばり方や道具の配置から、計画的な集団狩猟や解体作業が行われていたことが知られる。
(人類が集団で狩猟を行うようになったのは、チバニアン期の約20万年前からとされている。この時期にホモ・サピエンスが進化し、言語や集団的協調の能力を高めたことで、集団での狩猟が世界全体に広く伝播した。
ホモ・サピエンスが嚆矢であるかどうかについては、定かではないが、ホモ属の中でも、ホモ・エレクトスやネアンデルタール人も集団的狩猟を行っていた証拠が見つかっている。
集団での狩猟が本格化したのはホモ・サピエンスの時代とされている。 その進化の過程で、人類はマンモスやトナカイなどの大型動物を狩猟するために、戦略的な追い込みや罠を使用する技術を向上させ続けた。 )
ナウマンゾウは、更新世のチバニアン期(寒冷期)の約34万年前にユーラシア大陸から日本列島に渡ってきたと考えられている。当時、海面が下がり、大陸と日本列島が陸続きになる「陸橋」が形成された。この陸橋を通じて、中国北部からナウマンゾウは日本列島に移動して来た。特に長野県の野尻湖周辺では大量の化石が発掘されており、ナウマンゾウの研究が進められている。また、静岡県浜名湖や北海道幕別町忠類などでも化石が発見されている。それぞれの地域で異なる研究が進められている。これにより、各地の化石の時代的背景や環境、化石が出土した地層研究など、さらなる研究によってどの場所が最古の化石を保有しているかが明らかになるかもしれない。こうした知見が深まれば、過去の地球環境や生態系の変遷を理解するのにも役立つようになる。
チバニアン期の約34万年前の日本列島は、寒冷な気候が特徴で、氷期と間氷期が繰り返されていた。この時期、中国大陸との間に陸橋が形成され、動物たちが移動するルートが出来た。 ナウマンゾウ以外にも、以下の動物が渡ってきた。オオツノジカは、寒冷地に適応した種で、 マンモスもナウマンゾウと同様に寒冷地に生息していた大型のゾウや大型肉食動物のクマやオオカミなどの捕食者も移動してきた可能性がある。
中国大陸から日本列島に渡ってきたクマには、ツキノワグマUrsus thibetanusとヒグマUrsus arctosの両方が含まれていた。ツキノワグマはアジア全域に広く分布しており、日本では本州や四国に生息している。一方、ヒグマはより寒冷な地域に適応しており、北海道に生息している。 陸橋が形成された時期に、これらのクマが日本列島に移動してきたが、それぞれの生息地や環境への適応が異なるため、分布地域も異なっている。小型哺乳類のネズミやウサギなども陸橋を利用して移動したと考えられている。 これらの動物たちは、日本列島の生態系に大きな影響を与え、後の人類の狩猟対象にもなった。陸橋の形成は、動物だけでなく植物の分布にも影響を与えたとされている。
陸橋の形成によって移動して来た植物には、いくつかの興味深い種が含まれている。例えば、イネ科植物は陸橋を通じて移動した種子が新しい地域に定着し、旧石器時代の農耕の基盤となり有用な生業の一つになった。 ヒユ科アカザ属の一年草アカザなどの雑草は、陸橋を利用して広がり、ホウレンソウの仲間で味が似ているため、古代より食用として栽培もされ若芽や若葉を摘んで食べている。照葉樹林の構成するクヌギやコナラなどの樹木が移動し、森林の構成に変化をもたらしたばかりか、その果実のドングリは旧石器時代より大切な食用として工夫調理されてきた。 これらの植物は、動物の移動とともに生態系の変化を促進し、地域の多様性を高める役割を果たしてきた。
日本列島に人類が初めて登場したのは、更新世後期の約3万8000年前から3万年前頃とされている。この時期、現生人類(ホモ・サピエンス)が海を渡り、日本列島に到達したと考えられている。当時も氷期で海面が現在よりも低く、陸続きや近接していた地域が多かったため、渡来が比較的容易だった。当時の人々は、狩猟採集を主な生活手段としており、石器を用いて生活していた痕跡が各地の遺跡から発見されている。長野県の野尻湖周辺では豊富に出土している。
沖縄本島の中部、那覇市の北東約10 kmの宜野湾市の普天満宮本殿の近くにある洞穴から、約3万2000年前に人類が火をたいた炉跡が見つかった。2024年4月12日、沖縄国際大学で新里貴之准教授が発掘調査に関する会見を開いた。
諸外国でも古代人はドングリを食料として利用していた。
古代ヨーロッパでは、ドングリを粉にしてパンや粥のような食品を作ることが一般的であった。ドングリの渋みを取り除くために、水にさらしたり、灰を使ってアク抜きを行ったりしていた。
北アメリカのネイティブアメリカンは、ドングリを砕いて粉にし、水にさらして渋みを抜いた後、パンケーキやスープの材料に使っていた。
地中海沿岸では、ドングリを焙煎して粉にし、飲み物や食材として利用していた。 ドングリのアク抜きには時間と手間が掛かるが、古代の人々はその栄養価を活かすために様々に工夫を凝らしていた。
(旧石器時代の人々がドングリを食べていた可能性は研究されているが、直接的な証拠は未だ十分に発見されていない。一部の研究では、どんぐりの渋味成分タンニンを除去する方法や調理法が、古代人にも利用されていた可能性が示唆されている。また、縄文時代にはドングリが重要な食料資源であったことが知られているが、旧石器時代においても油とたんぱく質含量の多い肉とドングリの組み合わせ調理で、タンニン量が減少し、また官能検査でも渋味も顕著に低下している。古代人も、ドングリと肉の組み合わせで焼き上げれば渋味が軽減できることに気がつけば、渋いドングリでも食用としていたと思われる。 )
ナウマンゾウの絶滅の理由については、更新世後期に気候が急激に温暖化し、ナウマンゾウが適応していた寒冷な環境が失われたことが影響した。北大西洋地域で温暖化が起こったベーリング‐アレレード期(約1万4,700年前に起こった事象、最終氷期末期の区切りとなる。)に、その気候変動に伴う植生の変化が、ナウマンゾウの食料供給に影響を与えた可能性が高い。また当時の人類が狩猟の対象とし、ナウマンゾウの個体数が減少した可能性も指摘されている。これらの要因が複合的に作用し、約1万4,700年前頃に絶滅したとされている。
ナウマンゾウの化石は明治時代以降、全国各地で発掘されている。多数のナウマンゾウが北海道から九州にいたる日本各地に生息していたことが分かる。最後の化石として知られる、その日本橋浜町でナウマンゾウの化石が見つかったのは、1976年2月、都営新宿線の掘削工事中に浜町駅のホーム北端から30mの地点(日本橋浜町二丁目)で発見された。発掘された化石は、頭蓋や下顎骨をはじめ100点近く。その出土した地層から、約15万年前の東京周辺にはゾウの群れがいたことが分かった。発見された化石は、現在、八王子市教育委員会の管理下に移され、稲荷山行政資料保管施設に保管されている。この化石は「浜町標本」と名付けられ、地下約22mの地点から出土した。この地層は約1万5,000年前のものとされている。 ナウマンゾウは約1万5000年前に絶滅したと考えられており、この化石はその時代のものと一致している。
「浜町標本」と名づけられた化石には、少なくとも3個体分の骨が含まれ、さらに、同一個体で頭蓋・体軸・体肢といった全体骨格が出土した。全国でも初めてのことで、調査後に復元作業も行われ、肩と腰の位置が高く、ずんぐりとしたナウマンゾウの体型的特徴がより明確になった。復元されたナウマンゾウは、肩の高さは1.9mと、比較的小柄。頭骨・牙、そして臼歯の生え変わり方や体格などから40歳代の雌であるとされた。学術的にも非常に貴重な標本とされて、浜町標本に関する研究は現在も行われており、例えば2022年には「化石研究会会誌」に追加資料が発表されている。この研究では、浜町標本の骨格や周辺環境についての詳細な分析が行われていた。
ナウマンゾウPalaeoloxodon naumanniは、絶滅したゾウ科の一種で、パレオロクソドン属Palaeoloxodonに分類されている。この属は、アジアゾウElephas maximusと近縁であることが遺伝学的および形態学的な研究から明らかになっている。パレオロクソドン属は、新生代の新第三紀時代の最後の時代鮮新世が活動期であった。前時代の中新世終期の約550万年前に始まり、260万年前に最高潮に達していた。この時代から発見された嚆矢となる化石は、人類学の観点から評価高く重要視されている。アフリカ大陸に生息する最初の人類アウストラロピテクスの出現を示す。
この時代に、動物や植物の多くの種が、気候条件や地殻変動などに制約されながら、様々な地域に生息地を広げ始めた。これは植物と動物の両方のレベルで生物多様性の点で重要な変化の時代となった。その活動の場は、多くの場合、今日まで維持されている。
約500万年前の鮮新世前期には、地球規模での環境変化が勃発し、これがパレオロクソドン属を初め多くの動物の進化に影響を与えた。鮮新世時代になると大きな造山活動は終息したが、大陸の漂流は続いていた。大陸は既存の海からのゆっくりとした移動を続け、彼らが現在持っている場所のほんの数kmに到達していた。
この時代の最も重要なのが、北アメリカと南アメリカを結びつけるパナマの地峡の形成であった。これは地球全体の気候に多大な影響を与えた。地質学的に、これは『超越的現象』と言われる。約500万年前 パナマ地峡形成前では、現在では陸続きとなっているパナマやコスタリカなど、中央アメリカを構成する国々は、島々として分散していた。パナマ地峡が、北アメリカ大陸と南アメリカ大陸に接続した時期は、鮮新世の約300万年前だとされる。パナマ地峡は、文字どおりパナマプレート上に位置しており、環太平洋火山帯に含まれる。火山の噴火や地震、プレートの沈みこみによる反動からの跳ね上がりなどが、海底を隆起させた。
また、中新世後期の約700万年前からは、大気中二酸化炭素濃度の低下による寒冷化から、パナマ海峡付近を浅海化(海退)していた。パナマ地峡の最短部の幅は、わずか64kmという小さな陸地だが、地球に及ぼした影響は相当に大きい。パナマ地峡によって、北アメリカ大陸と南アメリカ大陸の間を通っていた海流が遮断された。「そのような大規模な海流の変化が、氷河時代の要因になったのではないか」とも考えられている。つまり、鮮新世終期の約300万年前に形成されたパナマ地峡が、現在の更新世(氷河時代)の要因のひとつとして考えられる。
地球規模の気候変動と生物生態系に大きな影響を与えた。この地峡の形成により、海流の流れが劇的に変化し、南北アメリカの生物多様性を誘引した。
パナマ地峡が形成されることで、大西洋と太平洋をつなぐ海流が遮断された。この結果、北大西洋の海流が強化され、現在のような熱塩循環が確立さ、寒冷化が促進した。両方の海、特に大西洋の冷却を引き起こした。北極海域への熱輸送が増加し、氷床の拡大が進んだ。これが地球規模の寒冷化を促進した。一方で、熱塩循環の確立により、地球の気候がより安定し、現在の気候パターンの基盤が形成された。
パナマ地峡の形成により、南北アメリカの動植物が互いの大陸に移動する「アメリカ大陸間大交差」が起こり、新しい生態系が形成され、多くの種が進化した。南アメリカから北アメリカへ移動した動物は、アルマジロやナマケモノなど、北アメリカから南アメリカへ移動した動物は馬やクマなどが、新しい競争環境が生まれ、一部の種が絶滅する一方で、新しい種が進化した。
極レベルでは、南極海と北極海の水温が急激に下がり、今日では地球上で最も寒くなった。
この間に海面に顕著な降下があり、これは現在水没している土地の残存断片として現れている。ロシアとアメリカ大陸を結ぶ陸橋の場合では、現在、これは殆ど水没しており、ベーリング海峡として知られている地域によって占められている。
ザンクリエンズ洪水
新生代新第三紀鮮新世時代の前の中新世末期、そのメッシニア時代Messinian ageの後半の約596万年前~533万年前に、地中海は部分的またはほぼ完全な乾燥のサイクルに入った。メッシニアンの塩の危機Messinian Salinity Crisisと呼ばれている。その間、地中海は、山岳地帯の出現により閉鎖され、ほぼ完全に干上がったとされている。
 |
| マルタ島は、地中海の中央にあるマルタ共和国領の島、シチリア島の南93 kmほどのところにあり、面積は246 km2、最高標高点は253 m。それでも同国内で最も大きな島である。島の東部には首都バレッタがあり、市街は世界遺産となっている。 海岸沿いの入り組んだ入り江が、温暖な港の景観を創りだし、高台のある丘に密集した人家の風景が、.典型的な地中海性気候が、年間を通して温暖で快適な生活を保障していることを物語る。 マルタ島の4月の天気は、曇りがちな日もあるが、全体的には穏やかで過ごしやすく、平均気温は18〜21℃程度で、最低気温も12〜14℃、降水量は少なく、雨の日は月のうち10日程度、マルタの風景に新しい息吹を吹き込む春、街路樹トベラの花がその鮮やかな色合いで再生を象徴する。 マルタヤグルマギクは、春から初夏(4月~6月)にかけて咲きく、特に5月が最盛期で、鮮やかな青やピンクの花が見頃を迎える。この時期は、マルタ島の自然が色とりどりの花々で彩られる美しい季節になる。マルタの国花であるヤグルマギクには「幸福」「優雅」「信頼」という花言葉がある。この花が、マルタの穏やかで美しい自然を象徴している。 庭園や田園地帯を彩り、明るい黄色の花が春から初秋にかけて咲かせるオオキバナカタバミのクローバーのような葉は、夜や日陰になると葉を閉じる習性がある。 ピンク色の花を咲かせる半つる性の低木で、地中海料理に欠かせないケッパ の蕾も採れる。 夏(6月~9月)は、暑く乾燥しており、平均気温は約28~31℃、湿度が低いため、日本の夏よりも過ごしやすい。この時期は晴天が続く。 鮮やかなピンクや白の花を咲かせる常緑低木セイヨウキョウチクトウは、乾燥や暑さに強いため、夏のマルタでよく見られる。 青い花が特徴のプルンバゴ(ルリマツリ)は、庭園や公園で涼しげに咲いている。 トベラは秋になると実が三つに裂け、鮮やかな赤い種が現れる。この特徴的な実は、秋の風景に彩りを添える。 <トベラ: 秋になると実が三つに裂け、鮮やかな赤い種が現れます。この特徴的な実は、秋の風景に彩りを添えます。セイヨウキョウチクトウ: 夏から秋にかけて咲き続けるこの花は、ピンクや白の鮮やかな色合いで目を引きます。 br> 冬(12月~2月): 温暖で雨が多い季節、平均気温は約10~15℃で、霜や雪はほとんど見られないが、雨の日が増える。それでも寒さは厳しくはない。 マルタの気候は、夏の乾燥した暑さと冬の穏やかな雨が特徴で、春には黄色や白、赤系統の花々が咲き乱れ、特にクラウンデイジーやオオキバナカタバミなどが目立つ。これらの花々は石灰岩地質に適応しており、マルタの自然環境に溶け込んでいる。 |
 |
これらの蒸発岩の状態と分布を研究することで、地中海が干上がったとされるメッシニアン塩分危機の詳細を解明し、それにより、当時の気候や海洋条件の変化を深く理解する手がかりとしている。
地中海周辺の地層には、塩分濃度が非常に高い環境で形成された堆積物が見つかっている。当時の気候は乾燥しており、蒸発が進んだ結果、地中海の水位が大幅に低下したと考えられている。しかも、降水量が少なかったため、地中海への水の供給が不足し、干上がりが急速に.進行していた。 この危機が、ジブラルタル海峡が地殻変動によって閉じたことが主な原因であり、その後のザンクリアン洪水によって再び海水が流入し、地中海が元の状態に戻ったことを明らかにしている。
地殻変動により、地中海と大西洋をつなぐ唯一の通路であるジブラルタル海峡が閉ざされた。これにより、大西洋からの海水供給が途絶えた。地中海に閉じ込められた海水が蒸発し、塩分濃度が急激に上昇した。塩分濃度が高すぎて、多くの海洋生物が生存できない環境になった。その一方で、陸上動物が干上がった地中海を渡り、新たな生息地に移動する機会が生まれた。
鮮新世時代開始の約533万年前、ジブラルタル海峡が再び開通し、大西洋から大量の海水が流入する「ザンクリアン洪水」が発生した。この洪水により地中海は再び海洋としての姿を取り戻した。いわゆるザンクリウス洪水が発生した。大西洋から地中海を占領していた場所に水が戻ることになった。
その間、隙間を通って流れた水の作用は、小さな水路を形成するまでそれを浸食していた。地中海で水位が安定して通常に戻るまで、水流は維持されていた。
中新世後期から鮮新世にかけて、地球全体で寒冷化が進行した。気候は寒冷化し、南極大陸は中新世よりもさらに氷床を拡大していた。北半球での氷床の発達もこの時代に既に始まっていた。ヒマラヤ山脈などの大山脈の形成される過程で、その上昇が同時に激しい岩石の浸食・崩顔を招き、これによって大量のカルシウム塩が海に流入していった。このカルシウム塩が大気中の二酸化炭素を吸収し海洋に石灰として固定し激減させ、これが寒冷化の進行を促した。この間、海洋水域には重大で大きな変化があり、パナマの地峡の出現の結果、大西洋と太平洋の間のコミュニケーションの断絶であった。
この寒冷化に伴い、森林が縮小し、草原やサバンナが広がった。この新しい環境が、長鼻目(ゾウ類)を含む動物たちの進化を促した。収集された化石によると、鮮新世時代に最初の人類が出現した。現代の動物相につながるものがほぼ出現している。寒冷化に伴い、 長鼻目(ゾウ類)などの中新世の温暖な環境で繁栄した生物は多様性を減少させていった。類人猿をはじめとする霊長目(サル類)も全体的に生息域や多様性を減少させるも、その中から中新世末期にチンパンジーの祖先と分かれていたヒトの祖先(ヒト亜族)がこの時代に発展した(いわゆる猿人)。アウストラロピテクスAustralopithecus、それは同属の最初の標本から始まったので、この人類は人類の起源において超越的であった。「南の」の意味のラテン語australisと「サル」の意味のギリシャ語pithēkosからの造語で、「南部の猿」「南猿」の意で、霊長目(サル目)ヒト科の絶滅した属であり、化石人類の一群である。約400万年前 ~約200万年前に生存していたとされる。石器はほとんど作っていないようだが、最後期の種のアウストラロピテクス・ガルヒでは原始的な石器、つまり無加工な自然石などを使っていた。かつては猿人と呼ばれていた。
長期間孤立した島大陸だった南米大陸が北米大陸に繋がったことで、多くの生物の両大陸間の行き来が可能になった(アメリカ大陸間大交差)。これによって北米の生物との生存競争にさらされた南米原産の生物は衰退し、絶滅したものも多かった。
これらの環境の激変が、パレオロクソドン属のような新しい種の誕生と適応を可能にした。パレオロクソドン属は、更新世にユーラシア大陸を中心に広く分布していたゾウ科の一群である。この属には、ナウマンゾウを含む多くの種が存在し、肩高4mを超える巨大な種から、島嶼化によって進化した小型種まで多様性が広い。
遺伝子解析や頭蓋骨の形態学的特徴から、パレオロクソドン属はアジアゾウ属Elephasと非常に近い関係にある。特に、頭蓋骨の構造や臼歯の形状が類似しており、これが近縁性の証拠とされている。
パレオロクソドン属は、アフリカで進化したゾウ科の祖先から分岐し、ユーラシア大陸に広がった。その過程で、環境適応や地理的隔離により多様な種が様々に進化した。ナウマンゾウは特に日本列島を中心に生息しており、寒冷な気候に適応した特徴を持っている。
ナウマンゾウは約1万年前に絶滅したとされている。その原因として、気候変動と人類の狩猟圧が挙げられている。
中国北部に生息していたナウマンゾウの絶滅理由には、以下のような要因が考えられている。更新世後期末期のヤンガー・ドリアス期(約12,900年前~11,700年前)、約1万年前に、地球規模での気候変動が進行した。氷期から間氷期への移行に伴い、寒冷な環境に適応していたナウマンゾウは生息地の縮小や食料不足に直面した。気候変動により森林が縮小し、草原やサバンナが広がることで、ナウマンゾウの生息環境が大きく変化しました。この環境変化が生存に影響を与えた。
更新世後期末期の約1万年前の中国北部では、農耕や牧畜はまだ本格的に行われていなかったと見られている。この時期の人々は主に狩猟採集を行い、自然環境に依存した生活を送っていた。狩猟技術は進化しており、細石器を用いた弓矢などが普及していた。小型で鋭利な石器を弓矢の鏃として使用し、効率的な狩猟が可能になった。集団狩猟の際に協力して大型動物を捕獲する技術が発展た。ナウマンゾウは大型哺乳類として狩猟の対象となり、個体数が減少した。これらの要因が複合的に作用し、ナウマンゾウは絶滅に至った。
農耕や牧畜は、完新世初期の約1万年前以降に徐々に発展した。中国北部では、後に新石器時代に入ると、農耕や牧畜が広がり、定住生活が始まる。この時代の人々の生活様式は、環境や技術の進化と密接に関連していたのが興味深い。
ナウマンゾウが日本列島に渡り、そして絶滅していった過程は、自然環境と人類の歴史が交差する興味深いテーマでもある。
ナウマンゾウの化石は日本各地で発見されており、特に長野県の野尻湖周辺で多くの化石が見つかっている。その同じ地層から大量の花粉が採取され、その当時の植生から気象状態や環境も推測されている。これにより、ナウマンゾウの分布や絶滅の過程についての研究が進められている。
ナウマンゾウの食料供給に影響を与えた植生変化としては、気候変動により、広葉樹と針葉樹の混交林が減少し、寒冷な環境に適応した針葉樹が優勢になった可能性がある。これにより、ナウマンゾウが好んで食べていた広葉樹の葉や果実が減少したと考えられる。
 野尻湖 野尻湖 |
| ナウマンゾウの化石は、長野県ではここの信濃町野尻湖以外では、信濃町柏原・長野市・中野市・上田市・青木村・小諸市・佐久市・佐久穂町・ 南牧村・富士見町など、11カ所ほどで発見されている。 |
ナウマンゾウは草食性で、主にイネ科の植物を好んで食べていたとされている。ゾウであれば、木の葉や果実も食べる。湿地や森林に生育する植物が重要な食料源である。ナウマンゾウが水草を食べた直接的な痕跡については、現在のところ明確な証拠は見つかっていないようだ。ただ、ナウマンゾウが湿地や湖沼周辺に生息していたことから、水草を含む水生植物を食べていた可能性が考えられている。特に、湿地環境に適応した植生がナウマンゾウの食料供給の一部を担っていたと推測されている。これらの植物は、ナウマンゾウが生息していた寒冷な環境に適応した食性の一部だったと考えられている。
これらの植生変化が、気候変動や地形の変化によるものとされている。ナウマンゾウの絶滅には、こうした食料供給の減少が重要な要因の一つだった可能性がある。
ナウマンゾウの大きな特徴は、前頭頭頂隆起(ぜんとうとうちょうりゅうき)にある。その特徴は頭の形は、横から見ると、四角く やや角張っている。アジアゾウは丸く、アフリカゾウはややとがった三角形である。体の大きさは、背までの高さでオスが2・4m~2・8mあり、メスは2mくらいで、メスの方が小柄であった。
そして、背中のカーブが肩と腰のところに高まりがあるのも特徴の一つ。
千葉県印旛沼・北海道幕別町忠類・東京都日本橋浜町・神奈川県藤沢市などから、まとまった化石が見つかって骨格復元が行われている。瀬戸内海からは、おびただしい量の化石が見つかっている。中国の一部にナウマンゾウとしている化石が見つかっているが、日本のナウマンゾウとまったく同じ種であるのかどうかは未だ不明。
目次へ
北海道に生息していたケナガマンモスやナウマンゾウの天敵として考えられる動物には、主に大型の肉食動物が挙げられる。例えば、更新世の時代には、シベリアや北アメリカから渡ってきた可能性のあるホラアナライオンPanthera spelaeaや、オオカミの祖先である大型の肉食性イヌ科動物が睥睨していた。
日本列島には、更新世後期の約2万年前以前に世界最大級の巨大なオオカミが生息していたことが化石記録から知られている。これらの大型肉食性イヌ科動物は、ケブカマンモスやナウマンゾウのような大型草食動物と同じ時代に存在していたが、直接的な天敵であったかどうかは不明である。ただこの巨大なオオカミは、後に日本列島に渡来した新しい系統のオオカミと交雑し、小型化したニホンオオカミCanis lupus hodophilaxの祖先となったと考えられている。
一般的に「オオカミ」と呼ばれているものは、「ハイイロオオカミ」のことで、学名はキャニス・ルプスCanis Lupus、実はユーラシアと北アメリカ原産の犬である。イヌやディンゴなど、30以上の亜種が認識されているが、一般的に理解されているように、ハイイロオオカミは自然に発生する野生の亜種のみで構成されている。オオカミの毛皮は通常、白・茶色・灰色・黒のまだら模様であるが、北極圏の亜種はほぼすべてが白であり、北半球に広く分布している。シンリンオオカミは北米の北西部に分布するハイイロオオカミで、ホッキョクオオカミは北米大陸最北部のツンドラ地帯に分布する体毛が白いハイイロオオカミである。また、タイリクオオカミは、ヨーロッパから極東まで、ユーラシア大陸の中緯度を東西にまたがって広く分布するハイイロオオカミを指す。このように、ハイイロオオカミは地域により、体が大きかったり小さかったり、体毛が変わっていたりする。こうした違いに着眼して、他の地域の仲間たちと区分されたが、すべてハイイロオオカミの「亜種」である。 この区分は微妙であることが多いので、分類学者によって意見が分かれ、いろいろな説が出されている。北米に生育するハイイロオオカミは、以前は26亜種に区分されていたが、細かく分けすぎたために区別が難しくなって、最近では5亜種にまとめ直されている。いずれにせよ、「種」は、地域ごとに形態の特徴が異なる「亜種」によってできあがっている。「種」が異なれば、基本的には交雑して子孫を残すことはできない。同じ「種」に属する「亜種」どうしは、自由に交配して子孫を繁栄させることは可能である。
ハイイロオオカミとイヌの共通の祖先は、絶滅した東アジアのハイイロオオカミの集団と考えられている。イヌは更新世後期の約4万~2万年前にこのオオカミの集団から分岐し、家畜化されてきた。この過程で、人間との関係が深まり、イヌは現在のような多様な犬種へと進化した。
興味深いのは、イヌの祖先が特定のオオカミの亜種に由来するのではなく、複数の地域で家畜化が行われた可能性があり、このため、イヌの遺伝的背景には様々なオオカミの影響が含まれている。
ハイイロオオカミとイヌは遺伝的に非常に近い関係にあるため、交雑が可能である。実際、彼らは遺伝的に非常に近い関係にあり、同じ種Canis lupusと見なされることもある。そのため、ハイイロオオカミとイヌの間で生まれる雑種はウルフドッグ(狼犬)と呼ばれる。ただし、「すべて」という表現に関しては補足が必要で、交雑が成功するかどうかは、個体の健康状態や繁殖能力、そして環境条件に左右される。野生環境でもオオカミとイヌの交雑が確認されており、特にオオカミの個体数が減少した地域では、こうした交雑が増える傾向がある。交雑によって生まれるウルフドッグ(狼犬)は、性格や行動が個体ごとに異なり、飼育には特別な配慮が求められる。そのため、交雑が可能である一方で、その結果や影響については十分な理解が重要になる。
ハイイロオオカミ(タイリクオオカミ)は、北アメリカ大陸やユーラシア大陸の北半球に広く生息している。彼らは非常に社会的な動物で、通常5~10頭ほどの群れを形成して生活している。群れはリーダーとなる夫婦を中心に序列があり、協力し合いながら群れを支えている。 狩りの際には群れで行動し、大型動物を狙うことが多い。例えば、獲物の親子を分断させたり、交替で襲い掛かって消耗させるなど、高度な連携を駆使して狩りを成功させる。また、獲物を捕らえた後はリーダーから序列の高い順に食べる。
夜行性で、夕方から夜にかけて活発に活動し、シカやウサギ、昆虫や鳥など様々な動物を食べる。繁殖期は冬で、群れのリーダー夫婦のみが繁殖活動を行い、子育ては母親が主に担当し、群れ全体でフォローする。
ハイイロオオカミは、ケブカマンモスやナウマンゾウのような大型動物にとって直接的な天敵ではないようで、それらの動物は非常に大きく、オオカミのような捕食者にとっては狩りの対象としてはリスクが高すぎる。これらの巨大な動物は、体格や力の面でオオカミを圧倒しており、オオカミが単独で襲うことはほとんど不可能である。 ただし、オオカミの群れが協力して、弱った個体や幼い個体を狙うことはあり得る。特に、食料が不足している厳しい環境では、オオカミがリスクを冒して大型動物に挑むことも考えられる。しかし、こうしたケースは非常に稀であり、通常はシカやウサギなど、より小型の獲物を狙うことが一般的で、マンモスやナウマンゾウのような動物は、むしろ人間の狩猟活動によって絶滅に追いやられたとされている。
オオカミはその生態系の中で重要な役割を果たしていたが、これらの巨大動物の主な脅威ではなかった。オオカミは群れで行動するため、弱った個体や幼い個体を狙うことがある。特に、ナウマンゾウやマンモスの子供たちは、群れから離れてしまった場合、捕食されるリスクが高かった。
また、オオカミは主に中型から小型の動物を狩ることが多く、マンモスやナウマンゾウのような巨大な草食動物は、むしろ間接的な生態系の一部として関わっていたと考えられる。
ロシア、シベリアの永久凍土の奥深くから、氷河時代の更新世後期を生きた2頭のホラアナライオンの子どもの死骸が見つかった。セミユリャフ川semyuelyakh-river沿いのシベリアの北極圏の洞窟で発見された冷凍の子グマスであった。すぐそばで一緒に発見されたため、最初はきょうだいだと思われていたが、放射性炭素年代測定によって、2万7,962年前と4万3,443年前に生きていた。いずれも生後1~2ヶ月頃に死んだことが分かった。死骸が荒らされた形跡はなかったが、頭蓋骨はひび割れ、肋骨は折れ、全身は異様に変形していた。ただ、子グマの毛皮やひげまでもは損傷していない。彼女の歯・皮膚・軟部組織・臓器はミイラ化しているが、すべて無傷であった。検死から、2頭は約1万5000年を隔てて起こった別々の土石流で死んだのではないかと考えられている。更新世後期、ホラアナライオンは、東シベリアに広く分布していたが、この種が生息していた痕跡は、ユーラシア大陸の大部分や、現代の北米アラスカあたりでも発見されている。
更新世時代の大型動物の多くと同様、ホラアナライオンもおよそ1万4,000年前、最終氷河期の終わりの更新世後期に起きた「第四紀の大量絶滅時代」に滅んだ。
更新世後期の最も凶暴な捕食者の1つであるホラアナライオンは、アラスカ州ユーラシア大陸とカナダ北西部で、先史時代の馬や先史時代の象など、様々な哺乳類のメガファウナMegafauna(大型野生動物の総称)を食べていた。
ホラアナライオンはよく繁栄し、現在のスペインからユーラシア大陸を横断し、北米のアラスカやユーコンまで分布していた。その姿は先史時代の洞窟壁画に広く描かれている。ヨーロッパの洞窟を支配していたホラアナライオンは、今から約3万年前~1万年前の間に生息していた。その名の通り、洞窟を住処としていたこのライオンは、現在のライオンよりも一回り大きかったとされている。体長は約2.5mにも及び、体重は約300kgに達すると推定されている。その強靭な体格は、厳しい自然環境下での狩猟に適していた。広大な草原や森林で狩りを行っていたと考えられている。彼らの獲物には、馬や鹿、さらには大型の草食動物であるマンモスが含まれていた可能性がある。
ホラアナライオンは、強力な顎と鋭い爪を武器に、多様な大型獲物を捕食していた。ホラアナライオンの狩猟スキルと肉食性は、彼らをその時代の生態系における頂点捕食者の地位に置いていた。ホラアナライオンが生きた時代は、氷河期であった。この時代のヨーロッパは、現在とは異なる厳しい気候条件の下でありながら、多様な動植物が共存していた。ホラアナライオンはこのような環境で進化し、繁栄した。ホラアナライオンの絶滅は、氷河期の終わりと密接に関連していると考えられている。気候の温暖化により生息環境が変化し、彼らの主な獲物であった大型草食動物の数が減少したことが、絶滅の一因とされている。
ベーリング‐アレレード期の温暖化が大型草食動物の減少に影響を与えた背景には、植生の変化が大きく関与している。この時期の急激な気候変化により、北アメリカを含む多くの地域で植生が大きく変化した。温暖化に伴い、森林が拡大し、草原が縮小した。大型草食動物は草原に依存していたため、食料資源が減少し、生息地が制限された。この植生の変化は、食物連鎖全体に影響を与えた。草食動物の減少は、肉食動物にも影響を及ぼし、生態系全体が不安定化した。
この時期は、ベーリング‐アレレード期の温暖化により、人類が農業を始める条件が整い、これに伴い、森林や草原が耕作地に変えられ、人類の活動もより活発化し、狩猟や農耕や家畜の放牧が広がることで、その土地利用の変化が、野生の草食動物が食料を確保するのが困難になった。人類が定住生活を始めることで、集落が形成され、それに伴い野生動物の生息地がさらに失われた。人類は燃料や建材を得るために森林を伐採した結果、植生が変化し、草食動物が依存していた植物が減少した。これらの土地利用の変化が、気候変動と相まって大型草食動物の生存を困難にし、絶滅を加速させた。これらの要因が複合的に作用し、大型草食動物の減少となりホラアナライオンの絶滅に繋がった。
北海道では、かつて大型の肉食性イヌ科動物が生息していた。特にエゾオオカミCanis lupus hattaiが注目されている。この亜種は、ハイイロオオカミの一種であり、北海道の生態系において重要な役割を果たしていた。エゾオオカミは、陸上動物を主な獲物としながらも、一部の個体は海産物を利用していたことが研究で明らかになっている。多くのエゾオオカミは専らエゾシカなどの陸上動物を食べていたが、一部のエゾオオカミ個体群では、海産物に強く依存した食性を備えていたことも明らかになっている。彼らが自然状態で海産物を多く利用していたとすれば、北海道にも「海辺のオオカミ」が存在していたことになる。海産物を多く食べている個体がいることが明らかになった。カナダ沿岸の一部地域では、サケなどの海産物を多く利用する「海辺のオオカミ」が知られているが、北海道のエゾオオカミもこれに近い生態を持つ。
化石については、北海道大学植物園や苫小牧市美術博物館などにエゾオオカミの骨格試料が所蔵されており、これらを用いた研究が進められている。また、縄文時代やアイヌ文化の遺跡からもエゾオオカミの骨が出土しており、これらの化石は彼らの生態や食性を復元するための重要な資料となっている。
かつての北海道では、イヌ科の大型哺乳類であるエゾオオカミが生態系の頂点に君臨していた。しかし、入植に伴ってエゾオオカミの駆除が行われたことや、1879年の大雪によるエゾシカの大量死でエゾオオカミの餌が不足したことなどが重なり、20世紀の初頭に絶滅した。エゾオオカミのように食物連鎖の頂点に立つ頂点捕食者は、生態系をコントロールする重要な役割を持っていた。
彼らの生存を脅かす要因としては、クロマニヨン人などの人類が狩猟を行い、ホラアナライオンの個体数に直接影響を与えた可能もある。また、大型の草食動物、マンモスやナウマンゾウを、日本列島の旧石器時代の人々は、主要な食料源としていた。特にナウマンゾウは、更新世後期の約4万年前から1万5千年前までの間、日本列島に広く生息しており、旧石器時代の人々にとって貴重な狩猟対象であった。
ナウマンゾウは、現代のアジアゾウよりもやや小型で、寒冷な気候に適応して全身が体毛で覆われていた。狩猟によって得られた肉は食料として利用され、骨や牙、そして毛皮は道具や衣類.、装飾品や威信財.として活用していた。ナウマンゾウの毛皮は旧石器時代の人々によって防寒用や敷物として使われていた可能性は高い。毛皮は寒冷地での生活において非常に重要な資産であり、保温性が高く、耐久性にも優れている。衣服や敷物、さらには住居の断熱材として利用されることが多かったとされている。
ただし、ナウマンゾウに関する具体的な考古学的証拠は限られており、毛皮の使用については推測の域を出ない。それでも、その用途の広さと同時代の他の地域での大型動物の毛皮利用の例から考えると、ナウマンゾウの毛皮も同様に活用されていた可能性は十分に想定できる。
重要なことは、狩猟には集団による役割分担が不可欠であり、優れた指揮者の存在も重要.であり、このような活動を通して社会的な結束が強まったとも考えられ、落とし穴など使う狩猟方法や槍のみならず、やがて弓矢の使用など狩猟具も急速に進化した。その一方で、氷河期の終わりに伴う気候の温暖化が、獲物の生息地の縮小に繋がり、獲物の減少を招いたとされている。加えて森林の拡大や草原の減少が、ホラアナライオンの生態的ニッチに深刻な影響を与えた。これらの要因が複合的に作用し、約12,000年前にホラアナライオンは絶滅した。興味深いことに、シベリアの永久凍土から保存状態の良いホラアナライオンの子供のミイラが発見されており、彼らの生態や絶滅の原因についての研究が進められている。
ヒグマは森林地帯を中心に活動し、しかも雑食性であるため、植物や小型動物、さらには魚類など多様な食物を捕獲していた。このように、ホラアナライオンとは食性が異なっていたため、直接的な食物の競合は少なかった可能性が高い。
ただし、厳しい環境条件や獲物の減少が進む中で、限られた資源を巡る間接的な競争があった可能性は否定できない。また、気候変動や人間の狩猟活動が両者の生存に影響を与え、最終的にホラアナライオンは絶滅し、ヒグマは食性の多様性からも、その後も生き延びたと考えられる。
北海道の環境では、厳しい気象条件のみならず、食料競争も彼ら自体が捕食の対象にもなり生存を脅かされる要因になった。人間も彼らの生存に影響を与えた重要な要因となり、その狩猟技術の向上により、マンモスやナウマンゾウ、ホラアナライオンやヒグマは人間による捕獲の対象となり、ヒグマ以外の肉食大型動物の絶滅の一因となった。これらの要因が複合的に作用し、最終的に『第四紀の大量絶滅時代』に至ったとされている。
旧石器時代以前の日本列島には、牛や馬は自然に生息していなかったとされている。これらの動物は、後の時代に人間によって持ち込まれたもので、例えば、馬は弥生時代以降に日本に導入され、農耕や移動手段として利用されるようになった。旧石器時代の日本列島では、ナウマンゾウやオオツノジカなどの大型動物が生息しており、これらが主な狩猟の対象となっていた。牛や馬のような家畜化された動物は、後の時代に農耕文化の発展とともに重要な役割を果たすようになる。
目次へ
トバ火山の大噴火
新生代第四紀更新世(氷河期)後期(約129,000年前〜約10,000年前)にあたる約74,000年前、インドネシアのスマトラ島にあるトバ火山が地球史上最大級の噴火を勃発させた。この噴火は「破局噴火(地下のマグマが一気に地上に噴出し、壊滅的な被害や寒冷化を引き起こす超巨大噴火)」レベルの火山爆発指数(VEI)で最大の8に分類される。この噴火により、約2,800km3もの火山灰や溶岩が放出され、現在のトバ湖という巨大なカルデラ湖が形成された。
 成層圏オゾン層の破壊 成層圏オゾン層の破壊 |
| 成層圏にオゾン層が形成されるのは、先カンブリア時代太古代の約35億年前、海洋に光合成を行うシアノバクテリアが誕生したことに始まる。これらのバクテリアが酸素を生成し、大気中に放出することで、オゾン層の材料となる酸素分子が蓄積された。 その活動が地球の大気組成を大きく変える契機となり、この現象が「酸素革命」や「大酸化事変」と呼ばれる程、原生代初期の約24億年前に本格的に進行し、酸素濃度が急激に上昇したことで成層圏のオゾン層形成が整った。 その後、紫外線が酸素分子を分解し、酸素原子と結合してオゾンを生成するプロセスが進行する。 成層圏の上層部の酸素分子O2に太陽光(紫外線)が当たると分解され、 O2 + hν(太陽光などの紫外線) ⇒ O + O 酸素原子ひとつずつに分離する。 分離された酸素原子Oは、放っておくと酸素分子 O2に結びつき O + O2 ⇒ O3 オゾンになる。 それは、成層圏の上層部で、酸素分子O2が光を吸収して、酸素原子Oに分かれて下層に落ちて行き、下層部にある酸素分子 O2と反応し、中間領域でオゾンO3 が生成され、オゾン層になる過程をたどる。 このオゾン層が地球の生命を紫外線から守る役割を果たすようになるまでには、さらに長い時間がかかり、酸素濃度が十分に高まる必要があった。この過程は地球の歴史の中で非常に重要な転換点となり、生命が陸上に進出する条件を整えることになる。 オゾン層が生命を紫外線から保護するようになったのは、古生代約4億7500万年前のオルドビス紀(約4億8500万年前~約4億5000万年前)とされている。この時期には、成層圏のオゾン濃度が十分に高まり、地表に到達する有害な紫外線を効果的に遮断できるようになった。これにより、生命が水中から陸上へと進出する条件が整い、多様な生物が進化する道が開かれた。 古生代のデボン紀(約4億1,920万年前~約3億5,890万年前)終期の約3億5900万年前に、オゾン層の破壊が始まった可能性が指摘されている。デボン紀はシルル紀に引き続き温暖で、海洋温度は現代と同じくらいの25℃前後、地域による温度差は少なかったと見られる。 「魚の時代」とも呼ばれ、強力なアゴを持つようになった魚類が脊椎動物として始めて生態系の頂点に立つ。その後期には4本足を持つ脊椎動物『両生類』が現れた。体長6mのダンクルオステウスは肉食性の魚で、アゴに並んでいるギロチンみたいな歯は、実は歯ではなくて歯の役割をするために変化したアゴの骨であった。 「デボン紀末の大量絶滅」の時代には、海の酸素濃度が極端に低くなる「海洋無酸素事変」が何度か繰り返されたと考えられている。そのデボン紀末には大規模な火山活動が頻発し、これにより大量の二酸化炭素が放出された。これが地球の温暖化を引き起こし、海洋表面の水温が上昇したため、水に溶け込む酸素の量が減少した。これは、温かい水は酸素を保持する能力が低いためである。 この温暖化によって海洋の層構造が安定化し、表層と深層の水の混合が減少し、その結果、深海への酸素供給が制限され、深海の酸素濃度が低下した。また海洋表層の水温が上昇すると、植物プランクトンの成長が促進され、これにより、プランクトンの死骸や排泄物が増加し、これらの有機物が微生物に分解される過程で酸素が消費され、無酸素状態が拡大した。 更に陸上植物が急速に進化・繁栄したことで、大量の有機物が海洋に流れ込み、この有機物が分解される過程で酸素が大量に.消費された。 このデボン紀後期に、地球から約65光年離れた場所で発生した超新星爆発が、宇宙線や紫外線を地球に送り込み、オゾン層を長期間にわたり破壊した可能性がある。この影響で、紫外線が地表に到達し、生物に遺伝子損傷を引き起こしたと考えられている。このオゾン層を破壊した影響は大きかった。 これらの要因が複合的に作用し、生物の大量絶滅を引き起こしたと考えられている。 成層圏の温度構造は高度によって変化しており、これが化学反応の速度と成分の分布に重要な役割を果たしている。 その化学反応の速度は通常、温度が高いほど速くなる。この関係はアレニウスの式(Arrhenius Equation)で表されるが、 成層圏では、地上から約20~50kmの間で高度が上がるほど温度が上昇する「逆転層」の特徴がある。このため、オゾン分解反応や生成反応の速度は高度によって変化する。 例えば、オゾンが紫外線を吸収して分解される際、温度が高いほどその速度が上がる傾向がある。温度が反応速度に影響を与えることで、O3・O2・Oの配置も温度の違いに応じて変化する。 しかも、成層圏での気温が安定していることにより、これらの成分の分布が局所的な反応条件によって調整される。 温度が変わると、分子や原子の運動エネルギーも変化する。これが特定の化学結合を壊したり、新しい結合を形成したりするエネルギーに影響し、反応過程や生成物の種類も変化する。 このように、成層圏の温度構造は化学的な現象全体に大きな影響を与えるため、オゾン層や大気の化学組成にも重要な役割を果たなど、温度が変わると分子や原子の運動エネルギーも変化するため、配置構造を一律に示すことは困難を伴う。しかも特定の化学結合を壊したり、新しい結合を形成したりするエネルギーに影響し、反応過程や生成物の種類が変化する。 成層圏での気温が安定しているが、これらの成分の分布が局所的な反応条件によって調整されいるため予見は不可能である。 以下は、オゾン層破壊に関与する代表的な主な化学反応式 - オゾンの分解 - O3 + hν(太陽光などの紫外線)→ O2 + O O3 + O → 2O2 hνは、光子のエネルギー、つまり光の形でのエネルギーを指す。この記号は主に化学反応で、光化学反応を表す際に使用される。 例えば、オゾン分解の第一段階では次のような反応が起こす。 O3 + hν → O2 + O ここで、hνは太陽光などの紫外線を意味し、このエネルギーがオゾン分子を分解するためのエネルギー源として使われる。 オゾンO3は非常に反応性が高い分子で、酸素原子Oが近くに存在すると、酸素分子O2を形成する反応が起こることがある。 O3 + O → 2O2 この反応は、オゾンが分解する際に発生する酸素原子が他の酸素分子と結合することで進行する。 具体的には、オゾンが紫外線や熱などのエネルギーを受けると分解し、酸素分子O2と酸素原子Oに分裂する。この酸素原子は非常に反応性が高いため、近くに存在する酸素分子と速やかに結合して新たな酸素分子を形成する。このプロセスは、オゾン層の形成や破壊において重要な役割を果たしている。 NOxサイクルは、窒素酸化物(NOとNO2)が触媒として働き、オゾンO3を分解する一連の化学反応を指す。 このサイクルは成層圏で重要な役割を果たす。窒素酸化物NOxは、主に人間活動、自動車や航空機の化石燃料の燃焼による排出や、また自然現象、雷などのによっても生成される。 成層圏に到達したNOxは、オゾンと反応を開始する。NOがオゾンO3と反応して二酸化窒素NO2と酸素分子O2を生成する。 NO + O3 → NO2 + O2 次に、このNO2が酸素原子Oと反応して再びNOと酸素分子O2を生成する。 NO2 + O → NO + O2 このNOのリサイクルが繰り返されることで、オゾンは効率的に分解される。 この反応は触媒的であるため、NOx自体は消費されず、何度も反応に関与できるため、オゾン破壊の効率が非常に高くなる。 |
この噴火が引き起こした大規模な森林火災は、確かに塩化メチルや臭化メチルなどの化合物を大量に放出した可能性がある。これらの化合物は、バイオマス燃焼によって自然に生成されることが知られており、成層圏に到達することでオゾン層に影響を与える。ただし、当時のオゾン層への具体的な影響については、直接的な証拠が限られており、科学的な議論が続いている。
トバ火山の大噴火が引き起こした大規模な森林火災は、確かに塩化メチルや臭化メチルなどの化合物を大量に放出した可能性があり、成層圏に到達することでオゾン層に影響を与える可能性もある。ただし、当時のオゾン層への具体的な影響については、直接的な証拠が限られており、科学的な議論が続いている。
また、トバ噴火による「火山の冬」現象が地球の気候に大きな寒冷化をもたらしたことは広く認識されているが、これがオゾン層の化学反応にどの程度影響を与えたかについては、さらなる研究が必要と言われている。
火山噴火によるエアロゾルaerosolには、主に火山灰や硫酸エアロゾルが含まれている。その火山噴火によるエアロゾルの影響は非常に広範囲で、地球規模の気候変動や生態系に大きな影響を与えることが知られている。特に、硫酸エアロゾルは、火山活動によって放出される二酸化硫黄SO₂が大気中で酸化され、硫酸H₂SO₄の微粒子として形成される物質で、非常に微細な液体または固体の粒子(PM2.5μm【マイクロメート】以下など)で大気中に浮遊する。自動車の排気ガスや自然現象(黄砂など)などが主な発生源でもあり、硫酸塩・硝酸塩・炭素成分・有機化合物などの化学物質を含んでいる。非常に微細であれば、肺の奥深くまで入り込み、呼吸器系や循環器系に悪影響を及ぼす。また成層圏に長期間滞留し、太陽光を散乱させ地球表面の温度を低下させる「日傘効果」により気候に悪影響を与える。例えば、1991年の20世紀最大級のピナツボ火山の噴火(フィリピンのルソン島西側にある)では、大量の硫酸エアロゾルが成層圏に放出され何か月も残留した。それにより全球規模の気温が約0.5℃下がり、オゾン層の破壊も著しく進んだ。
更新世後期の「トバ火山の冬」により、地球の平均気温が約5℃低下し、寒冷化が数千年続いたとされている。また、火山性エアロゾルは成層圏オゾン層にも影響を与え、紫外線の吸収能力を変化させる。これにより、ホモ・エレクトゥスやホモ・エルガステルなどの人類のみならず大型哺乳類が依存していた食物資源が減少し、生息環境が大きく変化した可能性がある。寒冷化により植生が変化し、多くの動植物が生息環境を失い、これがマンモスやマストドンなどの大型哺乳類の生存にも大きな影響を及ぼした。
ヴュルム氷期(約11万5千年前〜約1万年前まで続いた最終氷期)のトバの超巨大噴火は、ホモ属全体に壊滅的な影響を与えたと考えられている。ホモ・エレクトゥスやホモ・エルガステルなどの傍系の種は、これらの厳しい環境の中で生存し続けることができず、最終的に絶滅に至った可能性が高い。
トバ噴火後の急激な気候変動は、人類の歴史における最大級の試練の一つとされています。この時期、現生人類も過酷な環境に直面し、その生存を維持すること自体が非常に困難だったとされています。総人口がわずか数千人~1万人程度にまで減少したとする「人口ボトルネック効果」は、現生人類の遺伝的多様性の低下につながりましたが、これによって特定の遺伝的特性や適応力が強化された可能性も指摘されている。
このように、ホモ属の他の種が絶滅する中で、現生人類が生き残ることができた背景には、生物学的要因だけでなく、道具の使用や社会的な協力、コミュニケーション能力の発展といった文化的・行動的な要因も大きく寄与したと考えられます。これが現在の人類の進化の土台を築いたと言えるでしょう。
紫外線の増加は、特に、成層圏オゾン層の破壊によるUV-B【280~315nm】の増加が問題視されている。紫外線は、太陽光に含まれる不可視光線で、波長によってUV-A・UV-B・UV-Cの3つに分類される。UV-Bはその中間に位置し、地球のオゾン層によって一部が吸収されるが、残りは地表に到達する。UV-Bは人体においてビタミンDを生成する必須物質であるが、過剰な曝露はDNAやタンパク質に損傷を与え、皮膚癌や白内障の原因となる。また、植物の光合成系にダメージを与え、光合成効率を低下させ、葉の脱色など、成長が阻害される。例えば、植物プランクトンの減少は海洋生態系に大きな影響を与えるばかりか、それを餌にする動物プランクトンをも含む生態系全体に影響を及ぼす。
トバ噴火は「トバ・カタストロフ理論Toba catastrophe theory」でも知られるように、人類の進化に大きな影響を与えた。この理論によれば、噴火後の寒冷化により人類の総人口が一時的に激減し、遺伝的多様性が大幅に失われたと考える。この現象を「ボトルネック効果」と呼び、現代人類の遺伝的特徴が均質化させたと言う。これは、少数の生存者から現代の人類が進化したことを示唆している。
生存の危機に直面した人類は、環境適応力を高めるために技術や文化を進化させた。衣服の使用や洞窟壁画の制作など、文化的な躍進がこの時期に始まったとされる。
トバ火山噴火Toba supereruptionは、約74,000年前の更新世後期に、現在のインドネシアのスマトラ島にあるトバ湖で勃発した超火山噴火である。それは、少なくとも4回のカルデラ形成噴火の最後にあたり、それ以前のカルデラはカラブリアン期の約120万年前に形成された。これは、最後の噴火で、推定火山爆発指数が8であり、第四紀で最大の既知の爆発性火山噴火であり、地球史上、既知の爆発性噴火で最大であった。
この超巨大噴火が、世界的な寒冷化を引き起こしたとする説が「トバ・カタストロフ理論」と呼ばれる。火山灰やすすが空を覆い、南アジアでは長期にわたって森林が失われた。トバ火山噴火による火山灰は、東南アジア・南アジアを中心に厚く降り積もった。ベンガル湾を越えたインド・パキスタンでは、トバ火山由来のものとされる約7万年前の火山灰が2mもの層厚で堆積している。グリーンランドの氷床コアの酸素同位体比からもこの時期の急激な気候の寒冷化が推定されている。これは北半球で気候の寒冷化が生じた証拠とされている。一方、南極の氷床コアにはこの気候変動は記録されていない。
グリーンランドの氷床コアの酸素同位体比を解析することで、過去の気候変動を復元する方法が用いる。δ18Oとは、酸素の安定同位体である18Oと16Oの存在比を表す量。つまり、この解析では、氷床コアに含まれる酸素同位体(主に酸素-16と酸素-18)の比率を測定する。この比率は、過去の気温や降水量の変化を反映しており、過去の気候や環境変化を解析する。
グリーンランドの氷床コアは、過去の環境を記録している試料として採取する。この値は、試料が形成された時期の温度や水循環の状況を反映している。δ18Oの変動パターンを解析し、気候変動や環境変化の証拠を特定しておけば、温暖化や寒冷化の時期を推定することが可能となる。比較と統合、つまり他の地域や試料のデータと比較し、地球規模の気候変動や地域特有の環境変化を明らかにする。この方法は、古気候学や地球化学の分野で広く利用されており、過去の気候変動の復元や水循環の理解に役立っている。
具体的な手法として、グリーンランドの氷床から深層コアを掘削し、過去数十万年にわたる氷の層を取得する。各層の年代を決定するために、氷床コアの層構造や含まれるガス成分を解析する。同位体比の測定では、氷の中に閉じ込められた水分子の酸素同位体比(δ18O)を分析し、この比率が、過去の気温や降水パターンを示す指標と照合する。同位体比のデータを気候モデルと組み合わせて、寒冷化や温暖化の具体的な時期や原因を特定する。この方法により、約74,000年前の寒冷化が確認され、これが火山活動による変化などの要因によるものとされた。
インド中央部の人類はこの激甚な環境変化の中でも生き延びた。インドのマディヤ・プラデシュ州にあるダバの発掘現場では、8万~6万5000年前の堆積物の層から、太古の石器が見つかり、2月25日付けで英科学誌「ネイチャー・コミュニケーションズ」に掲載された論文によると、噴火の前後で同じタイプの石器が使われ続けた。したがって、1つの継続した集団がトバ噴火の影響下でも生き残っていた。
しかし、この劇的な気候変動は、当時生存していたホモ属に壊滅的な打撃を与えたと考えられている。ホモ・エルガステルやホモ・エレクトゥスなどの傍系種は絶滅し、現世人類のホモ・サピエンスも、総人口が1万人以下にまで激減したと推定されている。これは、遺伝的多様性の著しい減少、いわゆる「ボトルネック効果」を引き起こしたと考えられている。興味深いことに、衣服の起源もトバ火山と関連付けられている。ヒトジラミの遺伝子解析から、アタマジラミとコロモジラミの亜種分化が約7万年前に生じたことが分かっている。このことから、トバ事変後の寒冷化を生き抜くために、人類が衣服を着用するようになったという説が提唱されている。
ヘリコバクター・ピロリ菌の遺伝子解析は、現世人類がアフリカから世界各地へ拡散していった時期を示唆している。少なくとも5万8000年前以降に、人類はアフリカを出て世界各地へと移動していったと考えられている。トバ・カタストロフ理論を支持する研究者の中には、トバ火山噴火後に再びアフリカから人類が拡散したという主張もある。
ヘリコバクター・ピロリ菌(ピロリ菌)は、人類の進化と拡散の歴史を解明する上で重要な手がかりを提供する。この細菌は、約10万年前から人類の胃に寄生しており、人類の移動とともに世界中に広がった。
ピロリ菌は、胃の粘膜表面に生息している菌で、胃液には塩酸が含まれている。これは非常に強い酸性であり、ほとんどの菌は生息できない。ピロリ菌も同様に酸性の中では生きられないが、それでもピロリ菌は胃の中で生き続け。これは、ピロリ菌が「ウレアーゼ」という酵素を出し、そのウレアーゼは、胃の中にある尿素を分解してアンモニアというアルカリ性の物質を作り出す。このアンモニアにより塩酸が中和され、環境が酸性ではなくなる。これにより、ピロリ菌は胃の中でも生き続けることができる。
ピロリ菌の遺伝子型の多様性により、遺伝子型は地域ごとに異なり、これを解析することで人類がどのような経路をたどって移動してきたかを科学的に推測できる。しかもピロリ菌の遺伝子は人類の遺伝子よりも突然変異率が高いため、短期間(数千年〜数万年)の詳細な変化が解析できる。ピロリ菌は約6万年前にアフリカを離れた人類とともに進化し、アジア・ヨーロッパ・アメリカ大陸へと広がった。
ピロリ菌の遺伝子解析により、人類がアフリカから他の大陸へ移動した経路や時期が明らかになっている。ピロリ菌の遺伝子型は、人類の食生活や環境に適応して進化しており、地域ごとの人類の生活様式や歴史を反映している。ピロリ菌の遺伝子解析は、人類学や進化生物学の研究において非常に重要な役割を果たしており、遺伝子配列を解析する「次世代シーケンサー」の普及によってさらに詳細な解析が進んでいる。
トバ・カタストロフ理論は、「トバの超巨大噴火によって『火山の冬』が訪れ、氷河時代の寒気が一段と激甚化し、大気と景観に甚大な影響が及び、生物の生態系も波瀾を極めた。その劇的な寒冷化はおよそ6,000年間続いたとされる。その後も気候は断続的に寒冷化するようになり、地球はヴュルム氷期へと突入する。」と説く。まさに破局的な規模であり、噴出物の量は2,000km3を超え、1980年のセント・ヘレンズ山噴火の約3,000倍ものエネルギーが放出されたと推定されている。
この大噴火により、大量の火山灰が成層圏にまで達し、地球全体を覆い尽くした。太陽光を遮断した結果、『ヴュルム氷期』と呼ばれる、約11万年前〜約1万年前まで続いた最終氷期で、地球全体が寒冷化し、氷河が広がった。この時期には、気候の変動が人類の生活環境に大きな影響を及ぼした。それでも、ここにいた狩猟採集者たちは、変化に適応できていた。ダバの遺物が、これまでにアフリカ中期旧石器時代(約28万5000年~5万年前)のアフリカやオーストラリア、アラビア半島の遺跡で出土した同様の石器とほぼ一致すると考えられている。こうした石器技術の共通性から、研究チームは、ホモ・サピエンスが従来の説よりも早くアフリカから進出しており、ダバの遺物はそれをさらに強く裏付けるとみている。
このヴュルム氷期は、完新世の始まりとともに終了する。ヨーロッパアルプス地域では、その間、現在よりも平均で約6℃低かったと推定されている。地球全体が寒冷化し、北ヨーロッパ・北アメリカ・アジアの一部が厚い氷床に覆われた。海水が氷床として陸上に固定されたため、海面は現在よりも約120m低下した。ツンドラや針葉樹林が広がり、現在の温暖な地域でも寒冷な気候が支配的となった。その一方、この氷期は、地球の気候や生態系に大きな影響を与え、人類の移動や進化を促している。たとえば、海面低下により陸橋が形成され、人類や大型哺乳類が新たな地域に移動する契機となった。
ナウマンゾウが中国東北部から日本列島へ移動してきたのは、チバニアン期の約43万年前から30万年前の間と推定されている。この時期も氷期にあたり、海面が低下して陸橋が形成されていたため、ナウマンゾウが移動可能だったと考えられている。氷橋を渡ったという説もあるが、陸橋の形成が移動の主要な要因となった。この陸橋がユーラシア大陸と日本列島をつなぎ、ナウマンゾウが移動可能になったと考えられている。ナウマンゾウは、寒冷な気候に適応するために皮下脂肪を発達させ、全身が体毛で覆われていたとされている。この適応が、寒冷な日本列島での生息を可能にした要因の一つで、ナウマンゾウはその後、日本列島全域に広がり、約1万5000年前まで生息していたとされている。
トバ噴火による「火山の冬」は、食料や生息地の不足を招き、動物が新しい環境を求めて移動する要因となった可能性がある。例えば、寒冷化による植生の変化が草食動物の移動を促し、それに伴い捕食者も移動した可能性が高い。
この寒冷化は、動植物の生息地に影響を与え、大型哺乳類の移動を促したようだ。ただし、具体的な証拠や詳細な研究は限られており、噴火が直接的に大型哺乳類の大移動を引き起こしたかどうかは明確ではない。 マンモスは寒冷なツンドラやステップ地帯に適応しており、広範囲にわたって移動した。特に、食料を求めて季節ごとに移動することが一般的であった。氷期の間、海面が低下し、シベリアと北アメリカを結ぶベーリング陸橋が形成されたため、マンモスはこのルートを通じて新たな地域に進出した。
ナウマンゾウは比較的温暖な地域に生息しており、日本列島を含む東アジアで広く分布していた。氷期の寒冷化により、彼らも生息地を移動し、食料や適した環境を求めて活動範囲を広げていた。マンモスは長い体毛や厚い脂肪層を持ち、寒冷地での生活に適応していたが、ナウマンゾウはマンモスほど寒冷な環境には適応していない。それでも現代のゾウよりも寒冷な気候に耐える能力があった。ヴュルム氷期の終わりに伴う気候の温暖化や人類の狩猟活動が、これらの動物の絶滅に影響した。特に、マンモスは食料資源の減少や生息地の縮小により、最終的に絶滅した。
ヴュルム氷期の間、人類の移動は地球規模で大きな変化を遂げた。この時期の寒冷化と海面低下により、新たな移動ルートが形成され、人類の分布に大きな影響を与えた。海面が約120m低下したことで、現在のシベリアとアラスカを結ぶベーリング陸橋が出現した。このルートを通じて、現生人類(ホモ・サピエンス)がアジアから北アメリカ大陸へ移動したと考えられている。
東南アジアでは、海面低下によりスンダランド(現在のインドネシア周辺)やサフルランド(オーストラリアとニューギニアを結ぶ陸地)が形成され、人類がこれらの地域に進出した。ヨーロッパでは、氷床の南端に沿った地域に人類が集中し、比較的温暖な中東地域を経由して移動が行われた。厳しい寒冷環境に適応するため、衣服の使用や火の利用が進化した。また、洞窟や簡易的な住居を利用することで寒さをしのぎ、大型哺乳類(マンモスやナウマンゾウなど)の狩猟が主要な食料源となり、移動生活が一般化した。
北アメリカでは、約12,800年前に起きたとされる「ヤンガードリアス期」の急激な寒冷化も、火山活動や隕石衝突が原因と考えられている。この時期には、ケナガマンモスやマストドンを含む多くの大型哺乳類が絶滅したとされている。寒冷化に伴う植生の変化による食物供給の減少が、これらの動物の生存を困難にした。
ベーリング‐アレレード期(約13,700年前)やヤンガー・ドリアス期(約12,900年前~11,700年前)は、地球の気候が急激に変化した時期であり、マストドンの生態や生息環境にも大きな影響を与えた。マストドンは主に北アメリカに生息していた大型の草食動物で、森林や湿地帯を好んでいた。彼らは葉や枝、果実を食べることで生態系の一部を形成していた。
ベーリング‐アレレード期は温暖化が進んだ時期で、森林が拡大し、マストドンにとって適した環境が広がった可能性がある。ヤンガー・ドリアス期は寒冷化が進んだ時期であり、これによりマストドンの生息地が縮小し、食料の確保が困難になったことが絶滅の一因とされている。
チバニアン期(約77万年前から12万年前)は地質学的な時代区分であり、末期には火山活動が活発だった可能性がある。ベーリング‐アレレード期やヤンガー・ドリアス期はチバニアン期の終期の時代にあたる。火山活動が気候変動に影響を与えた可能性は否定できない。例えば、火山噴火によるエアロゾルの放出が寒冷化を促進した可能性がある。
ヤンガー・ドリアス期の寒冷化は、主に北大西洋の熱塩循環の弱体化や彗星衝突などが原因とされている。これらの要因が急激な気候変化を引き起こしたと考えられているが、チバニアン期の火山活動が直接的に影響を与えたという証拠は現在のところ見つかっていない。ただし、火山活動が気候変動に与える影響は広く認識されており、例えば火山噴火によるエアロゾルの放出が地球の寒冷化を促進する。ヤンガー・ドリアス期の寒冷化においても、火山活動が間接的に関与していた可能性は完全には否定できない。
アガシー湖は、かつて北アメリカ大陸の北半分に存在した巨大な氷河湖で、現在のカナダとアメリカの国境付近に広がり、マニトバ州・オンタリオ州北西部・ミネソタ州北部・ノースダコタ州東部・サスカチュワン州を覆っていた。アガシー湖は更新世終期の最終氷期(約3万年前~1万年前)の末期に形成された。氷床が溶けることで大量の融解水が集まり、氷河前縁湖として誕生した。湖の最大面積は約44万平方km2に達し、現在のカスピ海や黒海に匹敵する規模であった。約8,200年前にハドソン湾の氷が溶け、湖水がほぼ全て排水された。この湖の形成と消失は、海面上昇や地球の気候、地形などに大規模な影響を齎した。アガシー湖の名残として、ウィニペグ湖やマニトバ湖などが現在も存在している。
ヤンガー・ドリアス期の寒冷化は、北大西洋の熱塩循環の弱体化が大きな要因になっている。最終氷期の終了に伴い、北アメリカのローレンタイド氷床が溶け、大量の淡水がアガシー湖から北大西洋に流れ込んだ。この淡水が海水の塩分濃度を低下させ、熱塩循環を弱体化させた。結果として、北大西洋の暖流が停止し、ヨーロッパや北半球の寒冷化を引き起こした。
彗星衝突による塵やエアロゾルが大気中に放出され、太陽光を遮断し、寒冷化を促進した可能性がある。衝突地点としては、北アメリカのオクラホマ州・ミシガン州・サウスカロライナ州、カナダのアルバータ州などが挙げられている。ヤンガー・ドリアス期の彗星衝突に関しては、複数の地点で衝突が起きた可能性が指摘されている。これは、彗星が地球に接近した際に分裂し、複数の断片が異なる場所に落下した結果と考えられている。
彗星の断片が衝突することで、広範囲にわたる影響が生じ、地球規模の寒冷化を引き起こしたとされている。さらに、衝突地点の地層からはナノダイヤモンドや溶融ガラスなど、非常に高温・高圧の条件下で形成される物質が発見されており、これが彗星衝突の証拠とされている。
このような複数地点での衝突は、地球の気候や生態系に大きな影響を与えたと考えられている。これらの要因が複合的に作用し、ヤンガー・ドリアス期の急激な寒冷化を引き起こしたとされている。
(ナノダイヤモンドは、炭素原子が正四面体構造で結合したダイヤモンドの結晶構造を持つナノサイズの粒子で、その化学成分は主に炭素で構成されており、天然のダイヤモンドと同様の性質を持っている。
ナノダイヤモンドは、その特性を活かして、医療や生物学の分野で幅広く応用されている。ドラッグデリバリーシステム【DDS】では、ナノダイヤモンドは薬剤を運搬し、標的部位で徐放するためのキャリアとして使用される。
ナノダイヤモンドは遺伝子を運搬するためのキャリアとしても利用され、幹細胞治療や臓器再生を目指した研究が進められている。また、ナノダイヤモンドの蛍光特性を活用し、細胞や組織のラベリングやイメージングに使用される。これにより、病変部位の可視化や診断が可能になる。ナノダイヤモンドは抗菌性を持つため、医療機器やインプラントの表面コーティングに利用され、感染リスクを低減する。)
ローレンタイド氷床Laurentide ice sheetは、現在のカナダとアメリカ合衆国の北半分を覆う巨大な氷床で、氷河の跡は五大湖や氷河湖として見られる。ローレンタイド氷床は、最終氷期(約2万年前)に北アメリカ大陸の広範囲を覆った巨大な氷床であった。その中心は現在のハドソン湾周辺で、南は五大湖地域まで広がり、面積は約1,300万平方km2に達していた。
この氷床は、地球規模の気候変動に大きな影響を与えた。特に、氷床の融解やサージ(急速な氷河の前進)によって北大西洋に大量の氷山や淡水が流れ込んだ。この淡水の流入は、北大西洋の塩分濃度を低下させ、海洋の熱塩循環を弱めた。その結果、北半球の気温が急激に低下し、寒冷化が進行した。この現象が「ヤンガー・ドリアス期」なども寒冷期を引き起こしたとされている。
また、氷床の形成と融解は、地球の軌道変化や太陽放射量の変動、さらには大気中の温室効果ガス濃度の変化とも密接に関連している。
研究チームは、2万年前の氷河期に北米のほぼ全域を覆っていたローレンタイドLaurentide氷床が、予想をはるかに超えるスピードで融け、数十億トンの水を放出していたことを突き止めた。この溶解が「地軸のブレ」という自然要因による温暖化によって引き起こされたことから、人為的な温室効果ガスによる昨今の温暖化が、グリーンランドの氷床にどのような影響を及ぼすかに関心が集まっている。
その結果、氷床は、約6,500年前に完全に消滅するまでの間に、急速な溶解を2回ほど経験していたことが分かった。1回目は約9,000年前で、海面は年に1.3cmずつ、約7m上昇した。2回目は約7,500年前で、海面は年に1cmずつ、5m上昇した。ちなみに、現在の海面上昇幅は年に約3.3µm。
ローレンタイド氷床の融解は、ベーリング‐アレレード期の温暖化と密接に関連している。この温暖化期の約14,700年前は、最終氷期から間氷期への移行期にあたり、地球全体の気温が急激に上昇した。この気候変動により、ローレンタイド氷床を含む大陸氷床が融解し始め、大量の淡水が海洋に流れ込んだ。
この淡水流入は、北大西洋の熱塩循環を弱め、気候に大きな影響を与えた。その結果、ヤンガー・ドリアス期の寒冷化が引き起こた。 ローレンタイド氷床の融解による淡水化は、北大西洋の深層水循環(熱塩循環)に大きな影響を与えた。この循環は、塩分濃度と水温による密度差で駆動されるもので、地球規模の気候を調整する重要な役割を果たしている。 氷床の融解によって大量の淡水が北大西洋に流れ込むと、海水の塩分濃度が低下し、密度が減少する。その結果、冷たい海水が深層に沈み込む能力が弱まり、深層水循環が停止または減速する可能性が高まる。これにより、北大西洋の熱輸送が減少し、北半球の気温が急激に低下する寒冷化が引き起こされた。この現象も、ヤンガー・ドリアス期などの寒冷期の原因とされている。
ローレンタイド氷床の融解による淡水流入は、メキシコ湾流を含む北大西洋の熱塩循環大西洋南北熱塩循環Atlantic Meridional Overturning Circulation(AMOC)に影響を与えた。メキシコ湾流は、熱帯地域から北大西洋へ暖かい海水を運ぶ重要な海流であり、北半球の気候を温暖に保つ役割を果たしている。
しかし、氷床の融解による大量の淡水が北大西洋に流れ込むと、海水の塩分濃度が低下し、密度が減少する。この変化により、メキシコ湾流を含むAMOCの流れが弱まり、場合によっては停滞することもある。その結果、北半球の気候が寒冷化し、氷期のような環境になる。
ベーリング‐アレレード期やヤンガー・ドリアス期
ベーリング‐アレレード期やヤンガー・ドリアス期と関連が深いマストドンMastodonは、ゾウ目マムート科マムート属に属する大型の哺乳類の総称である。原始的なゾウ類でありマンモスや現生のゾウに似ている。新生代古第三紀始新世の約4,000万年前から更新世(氷河時代)後期の「大量絶滅時代」の11,000年前まで生存していた。
 更新世後期の「大量絶滅時代」は、ベーリング‐アレレード期やヤンガー・ドリアス期と関連が強いと考えられている。ベーリング‐アレレード期は、最終氷期の終了期の温暖化の始まりを指す。約14,700年前~12,900年前まで続いた。この期間は、チバニアン期の終期(約14,700年前〜12,900年前)にあたり、地球の気候が大きく変動した時期として知られている。この期間は、氷期から間氷期への移行期にあたり、急激な温暖化が特徴的であった。
更新世後期の「大量絶滅時代」は、ベーリング‐アレレード期やヤンガー・ドリアス期と関連が強いと考えられている。ベーリング‐アレレード期は、最終氷期の終了期の温暖化の始まりを指す。約14,700年前~12,900年前まで続いた。この期間は、チバニアン期の終期(約14,700年前〜12,900年前)にあたり、地球の気候が大きく変動した時期として知られている。この期間は、氷期から間氷期への移行期にあたり、急激な温暖化が特徴的であった。この温暖化は、北半球の氷床が急速に縮小し、海面が上昇したことと関連している。また、気温の上昇に伴い、植生や動物の分布も大きく変化した。急激な気候変動が特徴で、北半球では温暖化が進行したが、南極では「南極寒冷期Antarctic Cold Reversal」と呼ばれる寒冷化が同時期に発生した。
北大西洋では、この時期に海洋循環が活発化し、暖流が高緯度まで到達するようになっていた。この循環の再開は、北半球の温暖化を促進し、ヨーロッパや北アメリカの氷床を著しく縮小させた。
ベーリング‐アレレード期は、最終氷期の終わりを告げる重要な気候イベントであり、地球規模での気候変動のメカニズムを理解する上で非常に重要な時期とされている。この時期の後、ヤンガードリアス期という短期間の寒冷化が続き、最終的に完新世の安定した気候へと移行した。
北大西洋地域で温暖化が起こったベーリング‐アレレード期(約1万4,700年前に起こった事象、最終氷期末期の区切りとなる。)に、南極では寒冷化しており、北大西洋地域が寒冷化したヤンガー・ドリアス期(約10,800年前〜9,600年前)は、「寒の戻り」と言われる。氷床は前進し、動植物は後退し、南極は温暖化するなど、北半球高緯度が温暖化する時期は、南極が温暖化する時期に比べて千年スケールで遅れていた。 このように、温暖化と寒冷化が南北両半球高緯度地域で逆になる現象は、バイポーラーシーソーbipolar seesawと呼ばれるメカニズムで説明される。
バイポーラーシーソーは、千年スケールの温暖化・寒冷化の気候変動のメカニズムに関する考え方で、その原因に南北両半球高緯度における深層循環の変動が関係しているとされている。例えば、北大西洋における深層循環が止まると、それまで熱帯から運ばれていた熱エネルギーの行き場を失い、その余った熱が南大西洋に再分配され、南極域が温暖化するという。バイポーラーシーソーメカニズムは、地球規模での熱エネルギーの再分配から、地球の気候システムを制御していることから、その原因とされる深層循環変動の解明がいかに重要かを端的に示した。
北太平洋における深層水は、北大西洋高緯度域と南極海双方で沈み込んだ海水が起動する。南極海で形成された深層水は、南極海周辺を回る深層水(CDW)と北大西洋からやってくる深層水が混合する。
そのCDWからの流れの一部は、さらに南太平洋西岸を北上して、中央太平洋海盆をへて二つに分岐し、下部太平洋深層水になる。さらに南極海で新たに形成され、南太平洋西岸、フィリピン海をへて北西太平洋海盆からハワイ諸島方面へ東向きに流れる上部太平洋深層水(UCDW)の二つの海流から構成される。北太平洋深層水は、南極底層水起源が約70%、北大西洋起源が約25%と報告されている。
それに伴い生態系が大きく変化し、多くの種が適応できずに絶滅した。 ヤンガー・ドリアス期は、ベーリング‐アレレード期の後に訪れた寒冷期で、約12,900年前から11,700年前まで続く。この時期には、急激な寒冷化が起こり、北半球の高緯度地域で特に顕著であった。この気候変動は、熱塩循環の弱体化や彗星衝突などが原因とされており、生態系にさらなるストレスを与え、多くの種の絶滅を引き起こした。 これらの気候変動は、地球の環境を劇的に変化させ、人類の進化や文化の発展にも影響を与えた重要な時期でもある。
目次へ
熱塩循環
海洋大循環は「熱塩循環」とも呼ばれ、海水の温度と塩分の変化が循環を駆動する。海洋大循環の出発点となるのは、北大西洋のグリーンランド沖と、南極のウェッデル海の2か所とされている。この2か所で、大気による冷却と海水の高塩分化により密度を高め重くなった表層の海水が深層へと沈み込む。
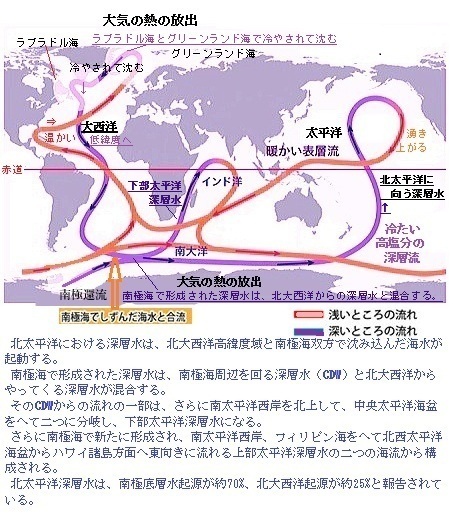 グリーンランド沖で沈み込んだ海水は図にあるとおり、大西洋を南下して南極海に達する。ここで、さらに沈み込んできた南極還流と合流した後、深層海流となってインド洋や太平洋へと北上する。北上とともに上層の海水と混合し、暖まり密度が低くなるに伴い上昇し、今度は表層海流としてインド洋からアフリカ大陸南端を流れ、大西洋を北上して、出発点であったグリーンランド沖へ戻る。グリーンランド沖を出発した海水が元の場所へ戻るまでの周期は、数千年にも及ぶと考えられている。
グリーンランド沖で沈み込んだ海水は図にあるとおり、大西洋を南下して南極海に達する。ここで、さらに沈み込んできた南極還流と合流した後、深層海流となってインド洋や太平洋へと北上する。北上とともに上層の海水と混合し、暖まり密度が低くなるに伴い上昇し、今度は表層海流としてインド洋からアフリカ大陸南端を流れ、大西洋を北上して、出発点であったグリーンランド沖へ戻る。グリーンランド沖を出発した海水が元の場所へ戻るまでの周期は、数千年にも及ぶと考えられている。海洋大循環はコロンビア大学教授.地球化学者ウォーレス・ブロッカーWallace Smith Broeckerにより、1980年代に提唱された。図で示される赤や青のリボンは「海洋のベルトコンベアーOcean conveyor belt(海洋大循環)」と呼ばれている。海洋大循環は極域から低緯度域へ向けて冷たい海水を、また低緯度域から極域へ暖かい海水を運ぶため、地球全体として温和な気候の実現する。それは極めて大きな貢献と言える。
「海洋のベルトコンベヤー」と海流は密接に関連しているが、一致しているわけではない。海流は主に風や地球の自転による力(コリオリの力force de Coriolis))で表層を流れるものが多く、それが黒潮や親潮のような名前で呼ばれる。一方、ベルトコンベヤーは深層海洋循環を含む、より広範な地球規模の流れを指す。この循環は、海水の温度や塩分濃度の違いによる主に密度差で駆動されるため、表層海流とは異なるメカニズムが働いている。
例えば、北大西洋で冷却された海水が沈み込むことで深層循環が始まり、赤道を越えて南極まで流れる。この流れは非常にゆっくりで、全体の循環には数千年を要する。具体的には、北大西洋のような高緯度地域の北極海で、海氷が形成される過程で放出された塩分を取り込んだ表層の海水が冷却されて、より密度が高く重くなり、深層へ沈み込む。この沈み込んだ水は、深層海流として赤道を越え、南極やインド洋、太平洋へと流れていく。そして、これらの地域で再び表層に浮上し、表層海流として循環を続ける。一方、表層海流は風の影響を受けて比較的速く動きやすい。つまり、ベルトコンベヤーは海流の一部を含むものの、その深層と表層の流れが異なるメカニズムで動いているため、完全に一致することはあり得ない。
ただ未解明な点も多いが、海洋が循環するメカニズムには「熱塩循環」以外に、「風成循環」がある。風成循環とは、風が海面を引きずる作用によって起こる海洋の循環である。地球の自転により、低緯度では東風(貿易風)、中高緯度では西風(偏西風)が強く。そのため、緯度が高くなるほど西風が強まる(東風が弱まる)。緯度15~45度の地域においては、北半球では時計回りの循環が、南半球では反時計回りの循環が海洋に形成される。
約45度より高緯度側では、緯度が高くなるにつれて西風が弱まる地域においては、それとは逆向きの循環が形成される。約45度より高緯度側で西風が弱まるのは、地球の大気循環とコリオリの力(地球の自転による影響)に関連している。中緯度地域では、偏西風(西風)が強く吹くが、緯度が高くなると、地球の自転速度が強まり、西風のエネルギーが分散されるため、西風が弱まる。
また、高緯度地域では、極地の冷たい空気と中緯度の暖かい空気がぶつかることで、風のパターンが変化しやすくなる。
風成循環
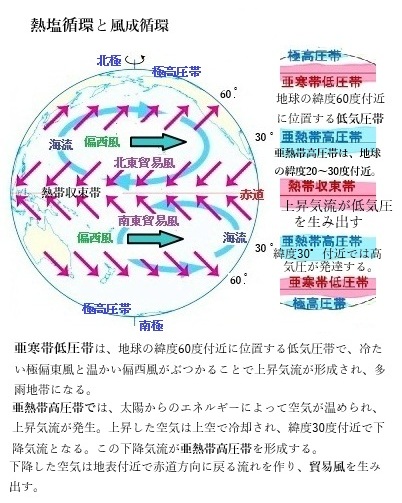 風成循環wind-driven circulationは、海面を吹く風が海水を引きずることで生じる海洋の循環であり、地球の自転の影響も受け、低緯度では貿易風(東風)、中高緯度では偏西風(西風)が強く。この風の作用により、北半球では時計回り、南半球では反時計回りの海流循環が形成される。日本近海の黒潮や親潮は、風成循環の典型的な例である。
風成循環wind-driven circulationは、海面を吹く風が海水を引きずることで生じる海洋の循環であり、地球の自転の影響も受け、低緯度では貿易風(東風)、中高緯度では偏西風(西風)が強く。この風の作用により、北半球では時計回り、南半球では反時計回りの海流循環が形成される。日本近海の黒潮や親潮は、風成循環の典型的な例である。黒潮は北太平洋の亜熱帯循環の一部であり、日本南岸に沿って流れる強い北向きの流れ(西岸境界流Western Boundary Current)となる。一方、親潮は亜寒帯循環の一部であり、冷たい海水を南に運ぶ流れとなる。これらの海流は、風成循環によって駆動され、海洋表層の循環を構成する。風成循環の深さは海面から数百m程度に限られるが、これらの海流は熱や塩分の輸送を通じて気候や生態系に大きな影響を与えている。
海面から1,000m深さ程度までの海洋の流れを、長期間にわたって平均すると、海洋にはいくつかの大規模な水平方向の流れがある。全球規模でみると、南極大陸周囲に流量の大きい東向きの海流(南極環流)があり、太平洋・大西洋・インド洋などの亜熱帯域などに大規模な循環が存在し、例えば、北半球亜熱帯域では時計回りの循環、南半球亜熱帯域では反時計回りの循環となっている。このような海洋表層の循環は、主に海上を吹く風が海面に及ぼす力(風応力)によって駆動されており、また、地球の自転による効果によって『西岸強化』と呼ばれる現象が見られる。
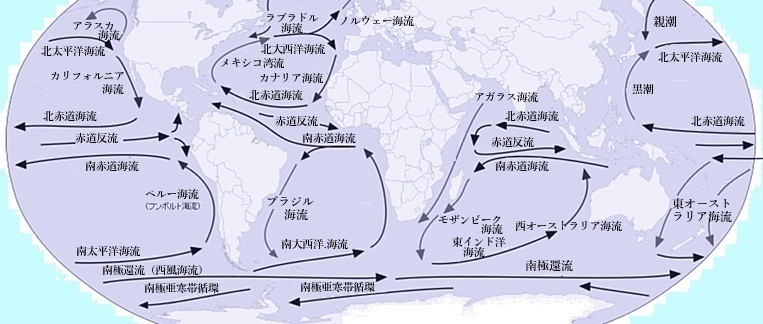
北半球の亜熱帯循環を例にすると、西岸付近に幅の狭い強い北向きの流れ、つまり『西岸境界流』と呼ばれ、北太平洋では黒潮、北大西洋ではメキシキ湾流があり、その東側はゆっくりとした南向きの流れがある領域になっている。
北太平洋海域をより詳細に見ると、熱帯域には反時計回りの熱帯循環、亜熱帯域には時計回りの亜熱帯循環、亜寒帯域には反時計回りの亜寒帯循環があることがわかる。このような循環は、いくつかの海流により構成され、亜熱帯循環では、北太平洋海流・北大西洋海流・黒潮および黒潮続流、カリフォルニア海流などの海流により、亜寒帯循環であれば親潮や亜寒帯海流などの海流により構成されている。
亜寒帯循環に関連する海流として、親潮以外にもノルウェー海流・ラブラドル海流・アラスカ海流が含まれる。これらの海流は亜寒帯循環の一部を形成し、寒流として特徴づけられている。ノルウェー海流は北大西洋の亜寒帯循環に属し、北極海へ向かう流れとなる。ラブラドル海流はカナダ東部沿岸を南下する寒流で、北大西洋亜寒帯循環の一部、アラスカ海流は北太平洋亜寒帯循環に属し、アラスカ湾を反時計回りに流れる暖流である。
アラスカ海流が暖流であるのは、主にその起源となる北赤道海流の特性にある。この海流は亜寒帯海流の一部でありながら、比較的暖かい海水を含んでいる。アラスカ湾内では、周囲の海水と比べて温度が高いため、暖流としての特徴を示す。アラスカ海流は反時計回りに循環し、暖かい海水を北米の沿岸地域に運ぶ。この流れが、カナダやアラスカの気候を温和にする。
亜寒帯低圧帯
亜寒帯低圧帯は、地球の緯度60度付近に位置する低気圧帯で、冷たい極偏東風と温かい偏西風がぶつかることで形成される。この地域の大気の流れの特徴は、冷たい空気と暖かい空気の衝突、つまり極地方から吹き出す冷たい極偏東風と、低緯度から吹き込む温かい偏西風が緯度60度付近で衝突する。この衝突により、冷たい空気が暖かい空気の下に潜り込み、暖かい空気が上昇する。これにより上昇気流が形成される。この上昇気流は、雲の形成や降水を引き起こし、この地域を多雨地帯にする。
極偏東風は、地球の極地方で見られる恒常風(卓越風)の一種で、この風は、北極や南極の極高圧帯から寒帯前線(または亜寒帯低圧帯)に向かって吹く東風を指す。地球の自転によるコリオリの力の影響を受けて、東寄りの風となる特徴がある。ただ、極偏東風は地表から約1~3kmの高さで吹き、風の強さや方向は不規則な場合もある。この風は、極地方の冷たい空気が原因で発生し、極循環と呼ばれる大気の循環システムの一部を形成する。
この上昇気流が発生することで、地表付近の気圧が低下し、低気圧が形成される。この低気圧こそが亜寒帯低圧帯の特徴となる。この地域には極前線と呼ばれる気温差の大きい空気の境界が存在し、気象現象が活発になる。
亜寒帯低圧帯は、地球の大気循環の一部であり、極循環やフェレル循環Ferrel circulationと密接に関連している。この循環は、地球全体の気候バランスを保つ重要な役割を果たしている。そのメカニズムは、極循環との関係では、極地方で冷たい空気が下降し、地表を低緯度方向に移動することで形成される。この冷たい空気が緯度60度付近で温暖な空気と衝突し、上昇気流を生じさせる。この上昇気流が亜寒帯低圧帯を形成する要因の一つとなる。
フェレル循環は、亜熱帯高圧帯(緯度30度付近)から亜寒帯低圧帯(緯度60度付近)に向かう空気の流れを含む。亜寒帯低圧帯では、極循環からの冷たい空気とフェレル循環からの温暖な空気が衝突し、低気圧が発生しやすい環境を作る。極地方の冷たい空気と中緯度の温暖な空気の温度差が、亜寒帯低圧帯の形成を促進する。この温度差により、極前線や寒帯前線が形成され、これが低気圧の発生を助ける。
フェレル循環によって、極東風や貿易風とは正反対の向きに風が発生する。これが偏西風と呼ばれる。偏西風はフェレル循環と極循環の境界付近で最も強くなり、ジェット気流と呼ばれる強い西風となる。
極高圧帯
「極高圧帯」という名称は、地球の気候や気象における特定の現象を示すために名付けられた。この名称は、地球科学や気象学の分野で使用される専門用語であり、極地方の気圧分布を説明するために適切な表現として選ばれた。この一帯は、地球の極付近に位置し、非常に高い気圧を特徴にする。極高圧帯は、冷たい空気が収縮し、地表付近で高気圧を形成することで生じる。この現象は、極地方の気候や風のパターンに大きな影響を与える。 極高圧帯の形成は、地球の極地方における特有の気象メカニズムによるものである。
極地方では、太陽エネルギーの受容が非常に少ないため気温が低くなる。冷たい空気は密度が高く重いため、地表付近に沈み込む性質がある。この沈み込む動き自体が、より収縮を強める。しかも、冷たい空気が地表付近に沈むことで、下降気流が形成される。この下降気流は、空気中の水蒸気を地表に押し戻し、雲の形成を抑えるため、晴天が多くなる。その一方、下降気流によって地表付近の空気が圧縮されるため、気圧は上昇する。これにより、極地方には高気圧が形成される。
極高圧帯は、地球の大気循環の一部であり、極循環と呼ばれる大気の流れとも関連している。この循環が、極地方から低緯度地域への冷たい空気の移動を促し、地球全体の気候のバランスを維持する。極循環も、地球の高緯度地域おける大気の循環パターンであれば、極地方の冷たい空気がどのように移動し、循環するかを説明するものとなる。
極地方では、太陽エネルギーの受容が少ないため、その大気は非常に冷えている。この冷たい空気は密度が高く重いため、地表付近に沈み込む。この沈み込む動きが下降気流を形成する。この地表に沈んだ冷たい空気は、極地方から低緯度の約60度付近に向かって移動する。それが風となるが、.直線的に吹くのではなく、地球の自転による「コリオリの力」が働くため、東から西への流れを作り出す。 この地表での空気の流れが、「極偏東風」と呼ばれる。北極や南極の極高圧帯から亜寒帯低圧帯(高緯度低圧帯)に向けて吹く風になる。 極偏東風は、地表の回転速度がより速い低緯度地域に向けて吹くため、地球は西から東に自転するため、地球上にいると、見かけ上は地表より遅い速度で西から東に移動(=回転)している。
このため、北半球では極偏東風は北東から南西に向けて吹くようにみえ、南半球では南東から北西に向けて吹くように見える。
緯度60度付近では、極地方から移動してきた冷たい空気と、低緯度から移動してきた暖かい空気が衝突して上昇気流が発生させる。この上昇気流は、上空で極地方に向かって移動する。この移動中に空気は冷却され、再び下降気流を形成するために極地方に戻る。
このように、極循環は地表と上空で異なる方向に空気が移動することで成り立っている。この循環により、地球全体の気候バランスを保つ重要な役割を果たし、極地方の乾燥した気候や低緯度地域の暖かい空気との湿熱交換に寄与する。
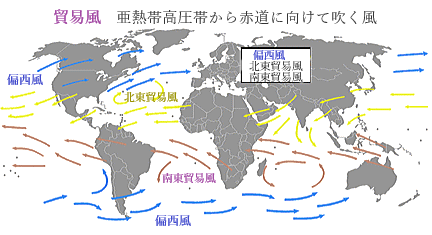
貿易風
亜熱帯高圧帯から赤道に向けて吹く風を貿易風と呼ぶ。貿易風はその名の通り、蒸気船登場前の風で動く帆船の時代にインド洋の交易に使われていた。赤道付近の熱帯収束帯から亜熱帯高圧帯(中緯度高圧帯)にかけては南北の温度差が少ないため、貿易風は地球の自転の影響を大きく受ける。南北の温度差が少ない理由は、北回帰線と南回帰線の間であり、季節によっては太陽がその真上に来る場所だからである。
 |
| バングラデシュのモンスーンは、熱帯モンスーン気候の典型的な特徴を示す。バングラデシュは、熱帯モンスーンの影響を強く受ける地域であり、雨季(通常6月〜9月)には大量の降雨が発生する。この期間中、年間降水量の約70%が集中する。その影響で、バングラデシュでは短期間に大量の雨が降り、このため、洪水が頻発.する。 モンスーンの終了期(10月頃)、世界最大のベンガル湾では、サイクロンが発生しやすくなり、これが降水量をさらに増加させる要因となる。バングラデシュのモンスーンは、地域の気候や生活に深く関わっており、特に低地や河川沿いの地域で洪水が起きやすく、洪水や農業の生産性に重大な影響を与える。 バングラデシュでは、貿易風とモンスーンが洪水に密接に関係している。貿易風はインド洋から湿った空気を運び、モンスーンの形成に関与する。 モンスーンの豪雨は河川の水位を急激に上昇させ、洪水を引き起こす主な原因となる。バングラデシュは低地が多く、河川が密集しているため、洪水が発生しやすい地形であるため、モンスーンの豪雨や貿易風の変動により、洪水が頻繁し、農地や住居に甚大な被害をもたらす。 バングラデシュの乾季(通常11月〜4月)は、平均気温は20℃~30℃程度で、比較的過ごしやすい時期とされている。乾季は降水量が非常に少なく、晴天の日が多いので、観光や屋外活動に適した時期とされている。 |
北半球では北東から南西に風が吹くため北東貿易風といい、南半球では南東から北西に向かって風が吹くため南東貿易風と言う。貿易風は太平洋東部のように開けた海洋では一年中一定方向に吹くが、陸地が近いインド洋北部や太平洋西部では季節風(モンスーン)によって乱される。
恒常風に分類されるが局所的にインド洋では季節風の影響で、船の往路と復路を別の季節にすることで帆船での航海に利用していた。低緯度での貿易風のメカニズムは、地球の大気循環とコリオリの力(転向力)によって形成される。赤道付近では太陽からのエネルギーが集中し、地表の空気が温められて上昇するため、この地域は「熱帯収束帯Intertropical Convergence Zone」と呼ばれ、頻繁に低気圧が発生する。この地域では、赤道付近の強い日射によって地表が暖められ、上昇気流が形成される。この上昇気流が低気圧を生み出し、熱帯収束帯は常に低気圧地帯として特徴づけられている。
特に、この地域では湿った空気が上昇するため、雲が形成されやすく、頻繁に降雨が発生するため、熱帯収束帯は熱帯雨林気候やモンスーン気候に大きな影響を与えている。
「熱帯収束帯」で上昇した空気は上空で冷やされ、緯度30度付近で下降する。この地域は「亜熱帯高圧帯」と呼ばれ、高気圧が形成される。そのため亜熱帯高圧帯から赤道に向かって空気が流れる。この風が貿易風と呼ばれる。
地球の自転により、北半球では風が進行方向の右側に、南半球では左側に曲がる。その結果、北半球では北東から南西に、南半球では南東から北西に風が吹く「北東貿易風」と「南東貿易風」が形成される。
貿易風は地球の大気循環(特にハドレー循環)と地球の自転による力の影響で生じる。
地球の自転が風の向きを変える仕組みは、「コリオリの力(転向力)」によるものである。地球は西から東へ自転している。このため、地表の速度は赤道付近で最も速く、極地に近づくほど遅くなる。
自転によって、移動する風や海流などは、北半球では進行方向の右側に、南半球では左側に曲がるように見える。これは、地球の自転による見かけの力で、実際には物体が曲がっているわけではなく、地球の回転が原因でそう見える。例えば、北半球で赤道から北極に向かって風が吹く場合、赤道付近の地表速度が速いため、風はその速度を保ちながら北に進む。しかし、北に行くほど地表の速度が遅くなるため、風は相対的に右側に曲がるように見える。
この現象は、地球規模の気象パターンや海流の形成に大きな影響を与えている。
インドの雨季(通常6月~9月)は、南西モンスーンの影響で非常に雨量が多く、時に荒れることがある。特に豪雨が短期間に大量の雨を降らせるため、河川の氾濫や都市部での洪水が頻発する。山岳地帯では、豪雨によって地盤が緩み、土砂崩れが発生する。更に、ベンガル湾で発生するサイクロン(熱帯低気圧)が雨季に重なると、さらに激しい暴風雨をもたらす。
インドの冬季(11月~2月)には、乾燥した北東貿易風が吹き、この風は大陸から海洋に向かって吹くため、インド全土で乾燥した気候が続く。これにより、農業用水の不足が懸念される。特に北インドでは、乾燥した空気が寒冷な気候をもたらし、夜間の気温が大きく下がることがある。
北東貿易風がベンガル湾を通過する際に水蒸気を取り込み、南インド、特にタミル・ナードゥ州では降雨をもたす。この現象は「北東モンスーン」とも呼ばれる。インドの気候は地域によって大きく異なるため、具体的な影響は場所によって変わる。
モンスーン(季節風)は、特定の地域で季節によって風向きが変化する現象を指す。この現象は、主に大陸と海洋の温度差によって惹起される。モンスーンの主な特徴は、風向きの変化で、夏季には海洋から大陸へ湿った風が吹き込み、雨季をもたらす。 - 冬季には大陸から海洋へ乾燥した風が吹き、乾季もたらす。その 気候への影響では、 モンスーンは、南アジアや東南アジアなどの地域で雨季と乾季を明確に分ける重要な要因となる。特にインドやバングラデシュでは、モンスーンの雨が農業や水資源に大きな影響を与える。
「モンスーン」の語源:は、アラビア語の「mawsim(季節)」に由来し、元々はアラビア海で見られる季節風を指していた。 モンスーンは、地域の気候や生活に深く関わる自然現象であり、特に農業や水資源管理において重要な役割を果たしている。
目次へ