バクテリアとプランクトン
(地球誕生史)
| TOP | 車山お知らせ | 歴史散歩 | 太陽系の形成 |
|
|


 生物学における「生物」の定義とは?
生物学における「生物」の定義とは?定義1) 細胞からなり、遺伝情報としてDNA(デオキシリボ核酸)を有している。
定義2) 生殖を行って増殖する。
定義3) 代謝してエネルギーを生産し、ATPにそのエネルギーを蓄える。
(代謝metabolismとは、生物細胞の生存の維持に関与する一連の生化学反応の総量である。)
定義1で、生物を定義する上で最も基本的なことは、どのような生物も細胞で構成されていることで、その細胞は細胞膜で覆われており、細胞膜はリン脂質とタンパク質でできている。その細胞の内部には、二重らせん構造をしたDNAを持っていることが重要なポイントで、DNAには遺伝子が含まれており、生命活動を行う上で必要となるタンパク質を作ることができる。DNAに書き込まれた遺伝情報を発現するには、まず遺伝子の塩基配列がRNAに転写される。
(細胞膜がない生物はいるのだろうか。それが生物か非生物かは別にしてインフルザウィルス、HIVウィルスなど病気を拡散させるウィルスは、実はその外殻はタンパク質でできている。したがって、ウィルスは細胞膜を持たないが、単純ヘルペスウイルスやインフルエンザウイルス、ヒト免疫不全ウイルスなど一部のウイルス粒子に見られる膜状の構造のエンベロープ envelopeが、ウイルス粒子【ビリオン】の最も外側に位置しており、ウイルスの基本構造となるウイルスゲノムおよびカプシドと呼ばれるタンパク質の殻・層を覆っている。エンベロープは、ウイルスが感染した細胞内で増殖し、そこから細胞外に出る際に細胞膜あるいは核膜などの生体膜を被ったまま出芽することによって獲得される。
実はウィルスが生物か非生物かという問題は、現在でも議論が分かれている。そもそもウィルスは、タンパク質の殻のなかに遺伝子だけを入れ込んだ存在だ。呼吸もしなければ代謝もしない。そのため、ウィルスをただの遺伝子を運ぶための粒子であり、生物ではないとする意見もある。その.一方で、自己の遺伝子を他の細胞に寄生することで複製しているという点から生物の多様性、進化の成果に衝撃を受ける。)
定義2の意義は、生物は子孫を残すために生殖を行って増殖する、という特徴がある。生殖には無性生殖または有性生殖があり、無性生殖は分裂や出芽などによって、有性生殖は2つの配偶子が接合することによって新個体が作る方法を言う。
無性生殖で増殖するときでも、有性生殖で配偶子を作るときでも、必ず細胞分裂を伴う。細胞分裂を行うために必要なのが、DNAを複製して分配するといった作業である。DNAの複製などには多くのタンパク質が関与するが、それらのタンパク質を合成する情報もDNAに含まれている。つまり、自分自身の力で次世代に遺伝情報を受け継いでいくことができる、それが生物の重要な特徴と言える。
(ウイルスは、タンパク質や核酸の合成に必要な素材を保有していない。従って、単体あるいは工培地では増殖できず、必ず生体細胞を必要とする。 生体細胞内に寄生して、細胞の代謝を利用して増殖し、材料や宿主細胞の代謝酵素、タンパク質合成のための宿主細胞リボソームを利用して自己に必要な成分を合成する。
細菌は基本的に2分裂によって増殖するのに対し、ウイルスは1つの粒子が感染した宿主細胞内で一気に数を増やす。 この時にウイルスが自分のコピーを複数増やすが、希にミスコピーが起きる。 これが俗に言う突然変異の原因となる。)
定義3では、生物は自身の生命維持や活動に必要なエネルギーを、自分の力で生産しているという特徴が重要となる。生命活動、例えば物質の合成や輸送、そのため筋肉の収縮などを行う際はエネルギーが必要となる。
エネルギー源であるATPの合成ために有機物を分解する反応(異化)である「呼吸」と、ATPを用いて有機物などの複雑な有機物質を合成する反応(同化)である「光合成」など、代謝を行うことで生命活動を維持している。
( ウイルスは、タンパク質質や核酸の合成に必要な素材を持っていない。 そのため単体あるいは工培地では増殖できず、必ず宿主細胞を必要とする。その生体細胞内に寄生して、細胞の代謝を利用して増殖し、材料、宿主細胞の代謝酵素タンパク質合成のための宿主細胞リボソームを利用して自己成分を合成する。)
生物の定義の1つでも当てはまらないものは「無生物」と呼ぶ。生物と無生物の境界は、「生物の定義」の基準を満たしているかというのが現在の生物学での見解である。ただ、生物か無生物かは科学者が便宜上定義したもので、自然界では線引きが難しい。今後新たな生物の発見などによって、生物と無生物の境界はより曖昧になっていくものと思われる。何よりも「生物の多様性」には、
☆ 同じ種でも遺伝子が異なる「遺伝的多様性」
☆ 様々な生物種が存在する「種の多様性」
☆ 様々な生物の相互作用から構成される様々な生態系が存在する「生態系の多様性」
「生物多様性」とは、簡単にまとめると「全ての個々の生物は互いに違いがあり、互いに繋がっている」。
ウイルスが生物かどうかは議論が分かれている。「ウイルスは宿主を利用するとはいえ、増殖して子孫に遺伝情報を受け継いでいることからも生物といえるのではないか」と言う専門家も多い。それでも学術界でもウイルスが生物かどうかは未だに議論が分かれている。生物か無生物かは科学者が便宜上定義したもので、自然界では線引きが難しい部分もあり、より研究が進めば、今後新たな生物の発見などにより、生物と無生物の境界はより曖昧になっていく。
ウイルスは種類によって様々な形状をしている。しかもシンプルである。外殻はカプシドと呼ばれるタンパク質の殻でできており、その内部にDNAもしくはRNAのどちらか一方だけを持っている。しかも、カプシドの周りをエンベロープという脂質二重膜が包み、そのエンベロープにスパイクタンパク質と呼ばれるタンパク質が存在する場合もある。また、ウイルスは細胞で構成されていないが、細胞膜とは異なる成分の殻を持っている。
ウイルスは生殖を行わない。また自身の力で増殖できない。しかも、ウイルスの持つ遺伝情報には最小限の情報しかなく、自身のDNAやRNAを複製したり、タンパク質を合成したりというシステムが備わっていない。そのウイルスの増殖は、「他の細胞に感染して代わりにウイルスを増殖してもらう」。感染する細胞のことを宿主と言うが、ウイルスは宿主のDNAの中に自身の遺伝情報をこっそり組み込み、ウイルスの遺伝情報やタンパク質を合成してもらうことで、大量に子孫を増殖する。
ウイルスは代謝を一切行わない。タンパク質の合成やエネルギーの生産、消費などの生命活動を全く行わない。必要最低限の遺伝子のみを持ち、まるで子孫を増やすことだけに専念している。生物とは、個体として機能するあらゆる生き物であるが、このような定義は、解決するよりも多くの問題を提起する。それは少なからず、個体としての概念が難しいからである。生物とは何かを定義するために、多くの基準が提案されているが、そのうちのほとんどが広く受け入れられている、最も一般的なものは、「生物が自律的な繁殖、成長、および代謝を行っている」ということにある。その「自律性の.多様性」が「生物多様性」の大きな要因となり、しかも多くの可能性を秘めている。
プランクトン
プランクトンplanktonは、水や空気の中を漂うが、海流や風に逆らって積極的に推進することができない生物でありながら多様な生物群系biomeがある。プランクトンを構成する個々の生物はプランクターと呼ばれる。海では、二枚貝・魚、シロナガスクジラなど、多くの大小の水生生物に重要な食料源になっている。「プランクトン」には、光合成により自ら栄養を獲得する植物プランクトン、自ら運動し外から栄養を獲得する動物プランクトンが存在する。
海洋プランクトンには、バクテリア・微視的真菌・古細菌・藻類・原生動物、および海の塩水や河口の汽水に生息する漂流動物または浮遊動物などが含まれる。淡水プランクトンは海洋プランクトンに似ているが、湖や川に生息している微小な生物群である。
これらのプランクトンは、水中の生態系において非常に重要な役割を果たしている。プランクトンには、植物プランクトンと動物プランクトンの二種類がある。
植物プランクトン
植物プランクトンは、光合成を行うことができるため、水中の一次生産者として機能する。これにより、酸素を生成し、有機物を作り出す。代表的な例としては、
クロレラ ; 緑藻の一種であり、淡水性単細胞緑藻類の総称。栄養価が非常に高く、ビタミン・ミネラル・、タンパク質、そしてクロロフィルが豊富に含まれている。
ケイ藻 ; 二酸化ケイ素SiO2(シリカ)でできた美しい殻を持つ非常に多様性に富んだ微小な単細胞の一群の珪藻類。ケイ藻は水中で光合成を行い、酸素を生成するとともに、食物連鎖の基礎を形成し、.多くの水生生物にとって重要な食料源となる。
シアノバクテリア ; 藍色細菌cyanobacteriaは、酸素発生型光合成を行う細菌の一群で、系統的には真正細菌に属する原核生物。陸上植物のものも含めて全ての葉緑体は細胞内共生において取り込まれたシアノバクテリアに由来する。単細胞、群体、または糸状体と、原核生物としては極めて複雑な体を持つものもいる。
動物プランクトン
動物プランクトンは、植物プランクトンや他の小さな動物プランクトンを捕食して生きている。一部の動物プランクトンは、他の有機物を摂取してエネルギーを得ている。これらのプランクトンは、魚や他の動物にとって重要な餌となる。代表的な例としては、
クラゲ類 ; 体がゼリー状で、透明なものが多い。クラゲの一部は成長して大きなサイズになるが、若い時期や小型の種類は動物プランクトンとして分類される。
オキアミ類 ;「魚の小さな稚魚」を意味するノルウェー語のオキアミに由来するが、オキアミは甲殻類であり、エビやアカザエビに関連する魚ではない。海洋生態系で重要な食物連鎖の一部となっている。
カイアシ類 ;海洋・淡水・湿地などの水中を浮遊する微小な甲殻類で、広範な分布を持ち、非常に多様な生物群であり、数百種類以上が知られている。様々な海洋生態系において見られる。極地から熱帯まで、さらには深海や淡水環境にまで広く生息している。
ミジンコDaphnia ; 淡水に生息する小さな甲殻類の一種で、微細な動物プランクトン、多くの水生生物の餌となる。ミジンコは生息する湖沼の環境が良いときは、単為生殖という方法で卵(単為生殖卵)を産み、子孫を増やす。この単為生殖は子孫を増やす際に雄を必要としない。つまり卵と精子の受精を行わないため非常に効率よく多くの子孫を増やすことができる。そして、この時の子孫は全て雌で、これを雌性単為生殖と呼ぶ。これは、魚などに捕食されるばかりのミジンコにとってはとても理にかなった子孫の増やし方と言える。
アメーバ ; アメーバとは、基本的に鞭毛や繊毛を持たず、仮足で運動する単細胞の真核生物である。移動の際は細胞内の原形質流動によって、進行方向へ細胞質が流れるに従い、その形を変えるようにして動く。この運動をアメーバ運動と言う。この時に原形質流動によって突き出される部分を、足になぞらえて仮足(かそく)または偽足(ぎそく)と呼ぶ。
淡水プランクトンは、魚や水生昆虫など多くの水生生物の餌になっており、生態系の底辺を支える重要な生物といえる。ツリガネムシVorticellaは、淡水に生息する、繊毛虫と言う動物的単細胞の真核生物の一群である。全身に繊毛という毛を持ち、これを使って移動する。多くのものが細菌や有機物の微粒子などを摂食するが、他の単細胞生物を捕獲して食べるなど、明確な捕食者として振る舞うものもある。ゾウリムシやラッパムシは、細胞にたくさんの繊毛を持ち高速遊泳や、急激な方向転換などを可能にしている。殻を作ってその中で生活をする有鐘類もいる。単細胞藻類を細胞内に共生させる種も知られている。
繊毛で水流を起こし、水中の有機物を食べることで水質の浄化にも一役買っている。淡水プランクトンは、湖や川の生態系の健全性を維持するために不可欠な存在で、水質の変化や汚染などがこれらのプランクトンに影響を与えることがあり、環境保護の観点からも重要な研究対象となっている。
ほとんどの場合、プランクトンは流れが彼らを導くところに漂うだけであるが、クラゲのようにゆっくりと泳ぐものもあるが、一般的に海流の影響を克服する程に十分な速さではない。プランクトンは一般的に水に生息していると考えられているが、大気中を漂流して生活する空中バージョンairborne versionsも存在している。これらの気生プランクトンaeroplanktonは、風に運ばれて空中に漂う小さな生命体で、その生物のほとんどは、サイズが非常に小さいか微視的なサイズのために識別するのが難しい。植物の胞子・花粉、風に散らばった種子などに含まれいる。また、陸域の砂嵐から空気中に流された微生物や、海の飛沫によって空気中に流された海洋プランクトンもいる。主に約40,000種類の真菌、数百種の原生生物Protist(真核生物のうち、菌界にも植物界にも動物界にも属さない生物の総称)、そして藻類・コケ類・苔類などの微生物が含まれ、それらのライフサイクルの一部がエアロプランクトンとなり、また多くの場合、胞子・花粉、風に散らばった種子と共に生きている。陸域の砂嵐から微生物が大気中に流され、さらに大量の空中浮遊する海洋微生物が飛沫として大気中に放出され、地球上のあらゆる地域に、1 ㎡あたり、毎日、数億個のウイルスと数千万個のバクテリアが堆積され続けている。
小さな漂流するエアロプランクトンは、大気中のいたるところで、1㎥あたり最大106個の微生物細胞の濃度に達する。この気団循環により、膨大な数の浮遊する空中生物を地上に分散させ、大陸間を移動し、やがて遠隔地で生き残り定住することで新たな生物地理学的なパターンを作り出す。この奔放さが各地域の環境を植民地化されるだけでなく、これらの生物の世界を飛び回る現象が、各地の生物環境を変えていく。
空気中の微生物は、雲の形成や降水など気象環境に影響し、しかも栄養循環にも関与し、広大な陸生生息地である既存の葉圏phyllosphere(微生物の生息地となる大地.から上部全体)の新たな形成となり、ひいては既存の生物に環境適応をせまり、それに応えられた生物が進化を遂げたとして命脈を保つ。
クラゲ
クラゲは、世界に存在に3,100種類以上、日本でも200種類はいる。大きさも様々で小さなクラゲは、5mmほどのベニクラゲから、世界一長い生物、触手の一部が37mにも達するキタユウレイクラゲなどがいる。毒針のある無数の細い赤銅色の触手がからまったような特異な形状をしている。この触手がライオンのたてがみに見えるため、英名では「Lion’s mane jellyfish(ライオンのたてがみクラゲ)」と名付けられている。クラゲは刺胞動物で毒液を注入する針(刺糸)を備えた細胞内小器官を持つ。クラゲの中でも傘を持ってその傘を閉じたり開いたりして、海を漂う。
クラゲの血液は海水で全身が心臓、脳は必要としない。クラゲは日常的に反射運動の繰り返しであれば、脳からの命令は必要としない。よって脳自体がない。しかも心臓と血液もない。クラゲは体内に取り入れた海水を循環させることで、全身に栄養を行き渡らせている。また海水を循環させる心臓ポンプの役割は、傘を開いたり閉じたりしながら泳ぐあの動きそのもの、つまりクラゲは全身が心臓のようなもの、その内臓もいたってシンプルで、消化器官と循環器官がひとまとめになった「胃水官系」という器官がある。傘の中心部についた口から海水とエサのプランクトンを取り入れると、この胃水官系を通して消化を行い、また口から吐き出す。必要な栄養を取り入れて、いらなくなったものを吐き出すまでの動きがひとつの器官で完結している。イソギンチャクやサンゴなども同じ仕組みの内臓を持つ。彼らを総称して「刺胞動物」と言う。
クラゲは大人になって性別が別れ、交尾を終えると死ぬ。その死体は水に溶けて完全に消失してしまう。クラゲは体を構成するものの97%が水分からできており、命が尽きると細胞がすべて水に溶ける。ただ生命をやり直す「ベニクラゲ」は、大人になると死なずにまたポリプpolypの状態に戻る。ポリプは、刺胞動物の体の構造のひとつであり、イソギンチャクのように基質上に固着して触手を広げるものを言う。ほぼ円筒形であるが、細長いもの、平らなもの、円錐形に近いものなど、その形は様々ある。
クラゲは、夜の海に浮上して水面のプランクトンやオキアミ類を食べている。あの大きな越前クラゲ(傘の直径 2 m・湿重量 150 kg)もプランクトンを食べている。たっぷり食べて海中に深く潜り、ゆっくりと食べ物を消化するクラゲもいる。しかも、カツオノエボシPhysalia physalis (Portuguese man o' war)などは魚を餌にしている。大西洋とインド洋でも発見されたが、主に太平洋に生息する太平洋のマンオブウォーやブルーボトルと同じ種と考えられている。それは多数の微細な毒の刺胞細胞を持っており、魚のみならず人間を殺すのに十分な強力な痛みを伴う刺し傷を負わせる。表面的にはクラゲに似ているが、ポルトガルのマン・オブ・ウォーPortuguese man o' warは、ゾイドZooidと呼ばれる多くの小さなユニットで構成されている。それぞれの形態にはかなりの差違がありながら、遺伝的に同一で、これらの異なるタイプのゾイドZooidが、それぞれが狩猟・消化・繁殖などで特殊な機能を果たすが、その群体が一緒になって単一の個体として機能している。約30億年を優に超える生物進化による多様性は止まること知らない。群体生物の一部のゾイドZooidのサイズと環境温度の間には相関関係があり、化石の群体内のゾイドZooidサイズの変化は、地質学的過去の海の温度と季節性の指標として使用されている。
生物の複雑な細胞組織やほぼすべての多細胞生物が、真核生物に含まれることから、真核生物の誕生は、生命の進化における画期的な出来事であった。真核生物の最終共通祖先は、現生するすべての真核生物の起源となれば、単一の個体によるのではなく生物学的な集団であったと見られる。2022年、アスガルド古細菌が複雑なアクチンベースの細胞骨格を持つことが明らかになり、真核生物の祖先が古細菌であることを示す最初の直接的な視覚的証拠が得られた。
(リボソームは、すべての生物の細胞内に存在する。タンパク質とRNA分子からなる小さな構造物で、リボソームは、mRNAをくわえ込み、アミノ酸の運ぶtRNAと相互作用しながら、タンパク質の合成【翻訳】)を行う重要な役割を果たす。特に、粗面小胞体の表面には多くのリボソームが付着しており、設計図に基づいて必要なタンパク質を合成する。)
目次へ
「合成生物学Synthetic Biology」の進展により、その有用性に着目し、従来の天然資源に依存した発見から、人為的に作成した多様性の中から新しいものを見出すことへとアプローチに変化している現状が、今日の合成生物学の背景にあると考える。医療や生命科学研究に有用なタンパク質を合成するために、生きた細胞にタンパク質の設計図を読みこませて、新たな細胞に作ってもらう方法が採られる。本来なら生き物が使わない新規なアミノ酸をも使えるように改変した細胞を用いると、目的のタンパク質に目印をつけたり、薬剤と連結したりすることが容易になる。また、これまでにない新しいタンパク質の形や働きを実現することができるかもしれない。新規アミノ酸のような人工のコンポーネントを含んだタンパク 質の生産技術の開発は、創薬に対しても最先端の技術支援にもなっている。
 新規アミノ酸を既存のタンパク質の望み部位に自在に導入する技術を使い、タンパク質の修飾を容易にしている(タンパク質の部位特異的修飾法)。今後のバイオ医薬品は、タンパク質に手を加えた修飾法で効用を向上させたり、昆虫細胞を遺伝的に改変して架橋アミノ酸のアジドフェニルアラニンAzidophenylalanine
(AzF)の導入を可能にしたりしている。しかも、AzFを3つの異なるタンパク質に組み込んだ後も、そのタンパク質の架橋が依然として観察されたことを示している。タンパク質の分析にこの新技術を活用して、タンパク質の機能の詳細な解析を可能にしている。
新規アミノ酸を既存のタンパク質の望み部位に自在に導入する技術を使い、タンパク質の修飾を容易にしている(タンパク質の部位特異的修飾法)。今後のバイオ医薬品は、タンパク質に手を加えた修飾法で効用を向上させたり、昆虫細胞を遺伝的に改変して架橋アミノ酸のアジドフェニルアラニンAzidophenylalanine
(AzF)の導入を可能にしたりしている。しかも、AzFを3つの異なるタンパク質に組み込んだ後も、そのタンパク質の架橋が依然として観察されたことを示している。タンパク質の分析にこの新技術を活用して、タンパク質の機能の詳細な解析を可能にしている。例えば、非天然のアミノ酸を酵素に組込むことで構造の安定化が実現できる「タンパク質の構造安定化技術」により、酵素の立体構造を安定化させるために、リジン残基の間にもともと存在していたスペースに非天然のアミノ酸を嵌め込んで酵素を安定化させもしている。非天然のアミノ酸をタンパク質の複数部位に導入することでタンパク質の構造安定性を高めることなどが、既に実用化されている。
「普遍遺伝暗号」という用語が存在するように、少数の例外はあるが、ほとんどの生物で遺伝暗号は共通であり、このことは生命進化の初期に既に遺伝暗号が成立したことを示唆する。つまり、アミノ酸のレパートリーが30億年以上ほとんど変化していないことになる。
いくつかのコドンを非天然のアミノ酸に割り当てるということを、基本的な開発コンセプトとして、大腸菌を使って非天然のアミノ酸をタンパク質に導入する手法により様々なものが開発されている。非天然のアミノ酸の導入位置は、遺伝子上でUAGコドンによって指定される。このとき、UAGコドンは非天然のアミノ酸を指定するだけでなく、本来の翻訳終止の働きもする。UAGコドンまでタンパク質合成が進んだところで、非天然のアミノ酸が伸長ポリペプチド鎖に取り込まれると、タンパク質合成は次の終止コドン(UAAかUGA)まで進み、期待される長さのタンパク質産物が得られる。現在、この大腸菌株の開発を持って、人為的な遺伝暗号の改変が本当の意味で成功したと考えられている。この大腸菌の中ではUAGコドンは、遺伝子中で何度使用しても良く、非天然のアミノ酸が何か所でもタンパク質に導入できることになり、しかもUAGコドンはもはや終止コドンとして働くことがないので、不完全長のタンパク質産物が生じることはなく、現在、非天然のアミノ酸が導入された完全な長タンパク質が合成されている。
代謝工学および遺伝子工学技術を使用して、生きた細胞に新しい機能を付与することが行われるが、この方法は、普遍的な遺伝子と比較し、必須でない遺伝子を排除して基本的なゲノムを作成することで、既存の細胞の複雑さを軽減しようとしている。これらの取り組みは、細胞生命の単一の起源、いわゆる「最後の普遍的な共通祖先」という仮説に基づいており、すべての生物を生み出した普遍的な最小ゲノムの存在を支持している。
しかし、最近の研究では、生命の系統樹を構成する真核細胞と原核細胞が、単一の細胞から分岐したのではなく、より遡る原始細胞のグループから進化した可能性が提起されている。その結果、普遍的な「最小ゲノム」の追求が難しくなる中、本質的でない機能をいくつも切り捨ててしまうと、生物の適応度が損なわれ、「もろい」ゲノムになってしまう。
「最小ゲノム細胞」というと、これこそ最初の生命に近いと考えるかもしれないが、実際進化可能な最初の細胞が地球上に生まれたときは、決してミニマルではなく、ある程度遊びがあったと考えられている。ミニマル・セルMinimal Cellだとたった一つの遺伝子に変異が起きただけで、進化どころか死んでしまうと想定されるからだ。
ところが、1999年Science誌の最終号に、あたかも来たる21世紀の先駆けとしての使命を背負うかの様に、900あるマイコプラズマMycoplasmaの遺伝子を500近くに減らしてもなお自律的に生きているMinimal Cellが報告されている。
2023年7月5日 Nature オンライン掲載論文の.「最小ゲノム生命その後」では、23年を経てこの最小ゲノム細胞を研究しているインディアナ大学から、最小ゲノム細胞でもある程度の進化は可能であることが示された。
「Evolution of a minimal cell(最小ゲノム細胞の進化)」の研究では、2,000世代まで細胞を維持してきた集団を、元の細胞と比べ、変異や適応が起こっているかを調べている。
(マイコプラズマ肺炎は、「肺炎マイコプラズマ(Mycoplasma pneumoniae」という細菌に感染することによって起こる呼吸器感染症で、小児や若い人の肺炎の原因として比較的多い。例年、患者として報告されるもののうち約80%は14歳以下で、1年を通じてみられ、秋冬に増加する傾向があると言う【厚生労働省.】。)
マイコプラズマは、自己複製可能な原核生物としての最小ゲノムを持っている。そのゲノムは通常、グアニン(G)とシトシン(C)含有量を比較的低くしDNAの再生を容易にしている。細胞膜には真核生物にしか見られないコレステロールも含まれている。マイコプラズマは、真核生物を宿主とする寄生細菌として、ヒトや脊椎動物の多くの病気の原因となっている。細菌とウイルスの中間のような性質を持つのが特徴である。実験室レベルでは、栄養培地で培養可能な種もある。培地で培養可能な最小の生物と位置づけられている。寄生虫または共生生物として、宿主生物から脂肪酸・アミノ酸、核酸の前駆体などの必須代謝成分を受け取る。これにより、ゲノムのサイズを小さくし、寄生生活様式に適合させている。そのため、原則として、宿主を殺さない。むしろ、それらは慢性感染症を引き起こし、宿主との適応関係を維持し、非常に成功したタイプの寄生虫を具現化している。
細胞内および細胞外細菌であり、主にヒトや脊椎動物に.寄生し、多くの病気の原因となっている。最小ゲノム細胞Minimal Cell(MC)だけでなく、元となったマイコプラズマにしても、突然変異の数が普通の細胞より100倍高い。さらに驚くのはMCでも同じ様に変異は起こっており、一般に考えられている様に変異の許容力が著しく低下するわけではない。さらに驚くのは、一塩基変異のみならず、挿入や欠損を伴う変異も、同じ程度に起こっており、それでも生きてきている。実際、変異の数や種類で見るとほとんどMCと既存の細胞間では差はない。ではこれらの変異は、細胞の進化を伴っているのか。ここではストレスを与える選択を行うのではなく、ただ増殖率を1代目と2,000代目で比較している。驚くなかれ、全く同じ培地を用いて培養を続けていても、増殖率が1代目と比べて2倍に上昇している。
すなわち、変異が細胞の適応性をさらに高めていることがわかる。ではどの様な遺伝子の変異が適応力に寄与しているのかを調べるため、アミノ酸配列が変化する様々な変異が集中している遺伝子を調べると、14種類の遺伝子が見つかり、たとえば細胞分裂の場所を示す ftsZC遺伝子変異を1代目の細胞で発生させると、増殖が高まることを明らかにしている。これは、MCでも、まだまだ合理化が可能であることを示している。
アミノ酸変化が起こる遺伝子は、有機合成に関わる酵素、特に脂肪代謝にかかわるものが多く、今後一つ一つ調べられると思う。 これまでの原核細胞進化研究では進化と共に細胞の大きさが上昇することが観察されており、既存の細胞でも2,000代を経ると、体積で10倍になるが、MCではこれは見られない。 おそらくゲノムを合理化して、許容力がなくなった結果がこんなところに見られるのだろう。 以上が主な結果で、MCも変異をベースに進化することがわかった。次は遺伝子が複雑化して遊びや許容力が生まれると、進化の質が変化するかが検討されるだろう。
合成生物学は、器官組織・細胞・遺伝子、そして分子といった生物の構成要素を部品と見なし、それらを組み合わせて生命機能を人工的に設計したり、人工の生物システムを構築したりする学問領域である。生物による創発的な行動を再現し、人工生命体を構築するために、非天然の化学物質が使用されもする。もう一方は、生物学的システムから交換可能なコンポーネントを探し出し、自然に存在しないシステムを組み合わせて作成もする。いずれの場合でも、分析によって容易に解決できない問題に関与し、解決するために、研究者に新しい領域への挑戦に挑む。それにより、新しいパラダイムparadigmが、分析だけでは容易には実現できない形で生まれもする。
従来の分子生物学では、生物個体から組織・細胞・分子・遺伝子へと細分化し、その機能を解析的なアプローチで取り組んできたが、合成生物学では、分子生物学などで蓄積された知見を生かしながら遺伝子を設計し、目的物質を生産するための代謝経路を構築し、それが機能する細胞や生物システムを作り出すという、構成的なアプローチが主流になる。それによって、本来交配できない生物種の遺伝子同士を一つの細胞内で働かせる。その組み合わせによるアウトプットは無限となり、更には種の壁を超えることも可能にした。
従来の手法だと手間やコストや時間などの技術的な制約から、「細部に分解して調べる」ことから始めたが、近年の技術革命により合成生物学の手法により「様々な物質を組み合わせて調べる」ということを容易にした。既に、細胞をプログラムして治療薬として利用したりする遺伝子治療や細胞治療、生物をプログラムして治療薬として利用する生物医薬品、植物や動物などの生物をプログラムしたりする遺伝子組み換え植物や、遺伝子組み換えサケなどが実現している。
「青い血液」を持つカブトガニの青い血は、細菌と接触したときに固まる機能を持っている。そのため、カブトガニの血液は抗菌や除菌を徹底する医療の現場で、大いに活用されている。
合成生物学の傘下に、各種の研究領域が統合されることは重要である。代謝調節・必須要素・生化学的構成などでは、生命の様々な側面に焦点を当てているが、これらの戦略はすべて、新しいタイプの生物を生み出すという同じ目的に向かっている。さらに、その様々な方法論は、多数の方法論的アプローチから始まり、やがて合成生物学のアプローチの多様性に繋がる。
合成生物学者は、様々な方法論・技術・研究機器を使用するにもかかわらず、新しい生命体を設計し、生産するという同じ基本目的を持っている。合成生物学は、単一の分野や技術だけではなく、多くの異なる科学分野から引き出され、そこからインスピレーションを得る学際的な分野が重要で、合成生物学の評価は、倫理的、法的、または安全性など調査領域は極めて広い。例えばリスクの問題は各技術に固有のものである一方で、合成生物学全体を考慮に入れる必要があるという事実と、その予測範囲を真摯に広げて検討しなければならないほどリスクは特別高い。
「植物修復法phytoremediation」は、植物を利用して、様々な供給源から環境に放出された化合物を固定化し取り込み、毒性の低減や安定化、または分解する。しかも、費用対効果が高く環境に優しい技術で修復できることが示されている。重金属・有機汚染物質・放射性核種・抗生物質・農薬などは、植物を使用して、有機および無機汚染物質から土壌と水を持続可能かつ経済的に修復する信頼できるソリューションになる。
原子力発電所の事故などが起こると、周囲に放射性セシウムなどが放出され、土壌が汚染される。その解決策として注目されているのが、植物に有害物質を吸収させる「植物修復法」である。 2021年には岩手大学と島根大学、東京大学大学院農学生命科学研究科の共同研究によって、世界で初めて「セシウムを効率的に取り込む植物タンパク質」の同定に成功した。この研究では、植物の遺伝子を編集し、セシウム輸送タンパク質を過剰に発現させることで、セシウムに汚染された土壌の浄化に役立つと期待されている。
植物の根の細胞にセシウムを効率的に吸収できるABCタンパク質ファミリーに属する2つのタンパク質、ABCG33とABCG37が発見された。今後は、この2つのタンパク質を過剰発現する植物を作成することで、土壌汚染で深刻な問題になっている放射性セシウムを効率的に除去できる「植物修復法」が実現できる可能性が高まった。
(本研究成果は、米国Cell Press社が発行する科学雑誌「Molecular Plant」に2021年2月13日【日本時間】に公開された。)
目次へ
アクチンは、重要なタンパク質で、すべての真核細胞に存在し、筋肉収縮の主要成分としても知られている。アクチンは、そのアミノ酸配列と機能によって主にβ(ベータ)、γ(ガンマ)など複数の形式に分類される。βアクチンは、細胞骨格の主要な構成要素であり、細胞の形状を維持する。γアクチンは細胞内外への物質の輸送、細胞の運動、および細胞間の接着にも不可欠な役割を果たす。これらは細胞の動的な構造と機能を支える基盤を形成し、生命現象を理解する上で中心的な役割を担っている。
真核生物の起源を特定するのは困難であるが、16億3,500万年前に生息していた原始的な緑藻、その最古の多細胞真核生物である微視的な化石が中国北部の古代中国の岩石層(チュアンリンコウ累層)で発見されたことは、真核生物が原生代中期まで遡れることを示している。約16億5,000万年前に生息していた最初期の明確な単細胞真核生物も中国北部で発見されている。
2015年以降、中国科学院南京地質古生物学研究所の古生物学者であるMaoyan Zhu氏とLanyun Miao氏は、同じ地域から岩石を収集し、それらを溶解し、最終的に279の微細な化石を発見した。化石は最大20個の円筒形の細胞のひもで構成されており、隣接する細胞壁は植物のように、顕微鏡で暗いリングとして見えると報告している。諸化石には胞子があり、独自の細胞壁があり、フィラメントが特殊な生殖構造を持っていたことを示唆している。そのフィラメントはUrospora属Urospora wormskioldii種のグリーンランドの海藻類などの現代の真核生物に似た緑藻類である可能性が高いと結論付けられた。
地球の総歴史(46億年間)に対する原生代の割合を計算すると、次のようになる。
原生代の期間 / 地球の総歴史 ≈ 45億−5億4100万 / 46億年
原生代は地球の総歴史(46億年間)の約86%を占めている。
累代(eon, aeon)は、地質時代の区分で最大のもので冥王代・太古代・原生代・顕生代の四つの累代がある。例えば顕生代(顕生累代)の名称は「肉眼で見える生物が生息している時代」という意味である。三葉虫をはじめとする化石として残りやすい殻や骨格を有する生物などが多く誕生し始めた時代であるカンブリア紀以後を指す。顕生代はカンブリア紀の始めから現在までのことで、約5億4100万年の期間であれば、地球誕生が約46億年前と考えられているので、顕生代は地球年齢の約1割ほどに過ぎない。
累代は複数の代eraからなり、その代は紀periodからなり、紀は世epochで構成される。現在の我々の時代は顕生代の新生代第四紀完新世に当たる間氷期にある。
累代(eon, aeon) ⇒ 代era ⇒ 紀period ⇒ 世epoch
環境
初期の生命の証拠は、西オーストラリア州ピルバラ・ブロック東部のマクフィー・リワード地域にある太古代前期の34億7000万年前における大陸地殻、ワラウーナ層群中の炭酸塩鉱物の重晶石bariteで発見されている。この鉱物の化学組成は硫酸バリウムBaSO4、結晶系は斜方晶系、21.1%もの硫黄分画を示し、これは硫黄の同位体の硫黄 32Sを硫黄34Sよりも容易に代謝する硫酸塩還元細菌の証拠と言う。
硫酸還元菌は、硫酸還元細菌と硫酸還元古細菌からなるグループであり、いずれも硫酸SO42-を最終電子受容体として、硫化水素H2Sへと還元する嫌気性呼吸を行う。これらの硫酸還元菌の多くは、亜硫酸塩(SO32-)、亜ジチオン酸塩(S2O42-)、チオ硫酸塩(S2O32-)、三チオン酸塩(S3O62-)、四チオン酸塩(S4O62-)、元素状硫黄(S8)およびポリサルファイド(Sn2-)などの無機硫黄化合物を還元する。
最終電子受容体として、好気呼吸では水H2Oの酸素分子O2を還元するのるはなく、無機硫黄化合物を還元するため硫酸を「吸う」のである。硫酸還元菌は35億年前までさかのぼることができ、地球上に生命が誕生して間もなく硫黄循環に貢献し、最も古い微生物に属すると考えられている。
 |
| 縞状鉄鉱床 先カンブリア時代の海底に堆積した酸化鉄を主体とする堆積鉱床である。主に産出するのは太古代初頭の38億年~原生代中期初頭の19億年前の年代の地層で、それ以後は原生代終期の約10億年前~5億4200万年前に一時期堆積した(新原生代酸化イベント)。 現在の世界の鉄鉱石の需要(年間約6億トン)の大部分を縞状鉄鉱床から産出する鉄鉱石がまかなっている。 縞状鉄鉱床の名は、鉄鉱石に富む部分と主にケイ酸塩鉱物から成る部分が、各々厚さ0.5~3cm程度の縞状に細かく互層していることから名づけられた。 特に27億年前~19億年前の時期に非常に大規模な鉱床が形成された。例えば北アメリカ大陸のカナダ東部のラブラドールからケベックにかけての鉄鉱床は1,000kmに達し、その先は切れ切れながらアメリカ合衆国の五大湖周辺まで約2,000km以上も続いている。その他オーストラリア(Hamersley)、旧ソ連(KurskとKryvyi Rih)、南米、インド、中国(吉林省)、南アフリカ、北ヨーロッパなどに大規模な鉱床がある。 『新原生代酸化イベント』は、約10億年前~5億4200万年前の原生代終期に起った。この時期には、地球の大気中の酸素濃度が大幅に増加し、生命の進化に大きな影響を与えた。地球全体が氷に覆われる「全球凍結(スノーボールアース現象)」が複数回発生した。この極端な環境変化が生物の進化を促進した。 エディアカラ生物群(南極大陸を除くすべての大陸で発見されている地球最古の動物の1つ)などの多細胞生物がこの時代に登場し、後の『カンブリア爆発』への道を開いた(多細胞生物の出現)。大気中の酸素濃度が増加し、真核生物や動物の進化に必要な環境が整った。 エディアカラ動物群は、約5億3900万~5億年前に絶滅して、原生代の終わりと顕生代の始まりの象徴となっている。顕生代は地球の歴史上「現生動物」が支配する時代である。 |
太古代の大気は、遊離酸素がほぼ完全に欠如していた。酸素濃度は現在の大気レベルの0.001%未満であり、一部の分析では現代のレベルの0.00001%まで低かったことを示唆している。30億年より前の海水温度も60~120℃という高温であったと推定されている。
しかし、酸素濃度の上昇の一時的な前兆は、この太古代の約29億年前以後は氷河堆積物が見つかるようになった。太古代を通じて大気中には酸素はなく窒素と二酸化炭素が主体であったが.、30億年前頃には、酸素発生型の光合成を行うシアノバクテリアが出現していた可能性がある。シアノバクテリアが形成したとおぼしき大規模なストロマトライトが広く分布していた。ただし、放出された酸素は縞状鉄鉱床の形成などに消費されていたと推測され、大気中酸素濃度の上昇にはつながらなかった。全地球規模での酸素濃度の上昇は次の原生代まで待つこととなる。
次の27億年前と25億年前にも前兆があったことが知られている。特に27億年前と25.01億年前での酸素化の増加の前兆は、どちらも大酸素化イベントの潜在的な出発点と見なされている。しかし、ほとんどの学者は、原生代初期の約24億年前に始まったと考えている。
約24億年前
ヒューロニアン氷期Huronian glaciationの開始は、24億年前のシデリアン(25億年前~23億年前にあたる古原生代最初の地質時代)から21億年前のリィアキアン(23億年前~20億5,000万年前にあたる古原生代二番目の地質時代)までの原生代に起こった氷期で、全球凍結状態(スノーボールアースSnowball Earth)に近い、きわめて深刻な氷期だったと考えられている。マクガニン氷期Makganyene glaciationとも呼ばれる。地球が史上初めて全球凍結状態に陥り、生物の大規模な絶滅を引き起こした。生物は辛うじて火山の付近のような熱水活動のある場や低緯度海域で生存していたと見られる。原因はシアノバクテリアが発生させた膨大な酸素がメタンと反応して水と二酸化炭素を生じ、強力な温室効果を持つメタンが減少したことによると見られる。
メタンは完全燃焼すると空気中の酸素と反応して、二酸化酸素と水になる。
CH4 + 2O2 → CO2 + 2H2O
この長い氷期の間に大気中へ放出された二酸化炭素が消費されず、温室効果を引き起こしてアイス・アルベド・フィードバックIce-albedo feedbackを突破して解氷する。アイス・アルベド・フィードバックは、地球の気候システムにおける重要なプロセスで、氷や雪は太陽光を反射する能力が高く、これをアルベド効果と呼ぶ。氷や雪が溶けると、反射率が低い地表、例えば海や陸が露出し、より多くの太陽光を吸収する。この結果、地球の表面温度が上昇し、さらに氷や雪が溶けるという正のフィードバックループfeedback loopが発生する。 このアルベド・フィードバックを突破するとは、このフィードバックループが進行し続け、気温の上昇が加速することを意味する。このフィードバックループは、地球温暖化の一因として重要な役割を果たす。
スノーボールアースは、先カンブリア時代後期の原生代のいくつかの地球規模の氷河作用に関する地球科学的な仮説であり、その最終段階は原生代終期の約5億8000万年前に終了した。これらの氷河期には、氷河が極から赤道に進み、海の大部分が凍りつき、地球のほぼ全面が氷で覆われた。
この仮説の地質学的証拠の信頼性と解釈に疑問が投げかけられている。なぜなら、当時の全氷期に対する明確な兆候があるが、それが赤道にまで及ぶ氷河作用の提案の物理的メカニズムに疑念が拭いきれないからである。
ティライト tilliteは、氷河によって運ばれた堆積物から成る岩石の一種で、氷河が解けることによって堆積された土砂や岩片が固まったもので、その特性は、科学者たちが氷期の年代を特定するのに役立つ。主に細かい粒子の土と石から構成され、地質学的な時代において氷河の存在を示す重要な証拠となる。ティライトなどの艶出し堆積物は原生代後期から受け継がれており、多くの場合、厚さはわずか数mであるが、場所によっては最大2,000mの厚さもある。
これらの堆積物は、南極大陸を除くすべての大陸で検出されている。堆積物の直接的な年代測定は不可能であるが、その形成時間は、その上にある岩石とその下にある岩石によって推定できる場合もある。古地磁気の復元によると、対応する堆積場所の少なくとも一部は、原生代後期を通じて赤道近くに位置していた。さらに、それらは、炭酸塩岩、赤い堆積物、蒸発岩など、より熱帯の条件下での形成を示す岩石と密接に関連している。この発見は、この時期に地球が赤道まで氷に覆われていたという推定に繋がる。
(『岩石に刻まれた磁気からたどる過去の地球』
地球はひとつの大きな磁石であるが、そのN極とS極は、数十万年に1回程度の頻度で逆転を繰り返している。岩石には、形成された時点での地球磁石の向きが記録されている。この磁気の記録を利用して地球の過去の姿を調べる。例えば、磁気をたどることで、具体的に大陸がどのように移動したのかも調べることができる【プレートテクトニクスの理論の確立】。
しかも、地球磁場の源は、外核における電磁流体現象によるため、常に変動しており、地球表面の 各地で異なる値を観測する。古地磁気学は地球上の1点における過去のある時点での地磁気の方位や強度の情報を記録していた岩石などの残留磁化から当時の地磁気の様子を復元し研究解析する。)
7億5000万年~5億8000万年前の原生代後期の少なくとも4つの氷河期に関しては、世界のほぼすべての地域で痕跡が検出されている。地球の全氷河期のうち少なくとも2つ、スターチアン氷河期(約7億1700万年~6億6000万年前)とマリノア氷河期(約6億5000万年~6億3500万年前)ばかりか、さらに、はるかに初期の氷河作用の痕跡も検出されている。
北アメリカにある五大湖のうちの1つヒューロン湖周辺の岩石から検出できる、いわゆるヒューロン氷河期は、原生代初期の約23億年~22億年前の気象現象である。このカナダの岩石からの古地磁気の発見は議論の余地があり、この時代の他の氷河岩については、赤道近くの堆積物が議論されている。
この 氷河期の発生原因一つが、当時存在していた超大陸ロディニアの分裂であった可能性がある。当時の降水は、超大陸の大きさのために、乾燥して砂漠のようだった地域まで到達することができない。その結果、そこの表面の岩石は物理的および化学的風化chimical weatheringにさらされた。雨水に溶解した大気中の二酸化炭素CO2は、ケイ酸塩鉱物 CaSiO3 の岩石の分解に重要な役割を果たす(炭酸風化)。 その過程で、二酸化炭素は炭酸水素塩HCO3−に変換され、川に流され、最後に流出した雨水とともに海に流れ込み、石灰 CaCO3として沈殿して海底に堆積した。大気中から温室効果ガスが除去されたため、気温が下がり、地球全体に氷河作用を及ぼした。
CaSiO3 + 2CO2+ H2O → Ca2++2HCO3−+SiO2 (1)
(二酸化珪素 SiO2濃度は雨水量に強く依存してはいない)
Ca2+ + 2HCO3− → CaCO3+CO2+H2O (2)
(化学的風化は、主として大気中 や土壌中の二酸化炭素が降水や地下水に溶け込んで炭酸H2CO3となり、それが岩石中に含まれる鉱物を溶解したり水和するような現象を言う。)
マリノアン氷河期の名称は、南オーストラリア州の州都アデレードの郊外Marianoにちなむ。氷河はおそらく全球を被覆するスノーボールアース事象が発生した。マリノアン氷期における特徴的な氷河の堆積物は地球が史上最も厳しい氷河期にあったことを示す。氷河はリズミカルなパルスで成長と後退を繰り返し、赤道付近まで到達した。マリノアン氷河期は短いが、それでも世界的な氷河作用であった。他の極低温氷河期は、マリノアン氷河期やスターチアン氷河期に比べて、おそらく小さく、全球的ではなかった。
氷河期の終焉は、火山による二酸化炭素の放出とメタンハイドレートの溶解によって温室効果が上昇する原因となり、また赤道永久凍土からのメタン放出によって加速された可能性もある。
地質学的な変化によって多くの証拠が失われているにもかかわらず、現地調査では、中国やスヴァールバル諸島、南オーストラリア州でマリノア氷河作用の証拠が示されている。中国の貴州省では、火山灰の層の下にジルコンZrSiO4鉱物を含む氷河岩があり、その放射性同位元素による年代測定が可能となった。
(ジルコンには放射性元素であるウランやトリウムが多く含むため、精度の高い年代測定が可能になる。)
目次へ
「微生物microorganism」とは、肉眼では観察できない極めて微視的なサイズの生物であり、単細胞の形で、または細胞のコロニーとして存在する。微生物は非常に特異な生息地を持ち、極から赤道まで、砂漠・間欠泉・岩・深海のいたるところに生息している。非常に暑いまたは非常に寒い条件などの極端な環境に適応するものもあれば、高圧の条件ばかりか、脱水・真空・酸に耐えることができる好極性微生物も知られている。生物のゲノムは、その環境と直接的または間接的に相関している。様々な微生物は、太古代の苛烈な嫌気性環境を含む様々な環境で生存し、存続するために進化してきた。そして、その多くの遺伝子は、過去に存在した微生物の進化の初期に重要な役割を果たしてきた可能性がある。ところが、現在、様々な生態系に位置を占める数え切れないほどの微生物のうち、人が発見できたのは全体の1%にすぎない。
微生物集団の大部分は、極限環境に適応する。例えば、人間の消化管・土壌・熱水噴出孔・深海底などには多くの嫌気性細菌が存在する。近年の技術の進歩により、これらの環境における細菌の生態の解析が進んでいる。現在、嫌気性細菌は、バイオ燃料・発酵、・酵素・ワクチン・ビタミン・乳製品などの関連産業を発展させてきた。それには好極性微生物のゲノム解析が、大きく貢献している。
デイノコッカス・ラジオデュランズDeinococcus radiodurans(属:デイノコッカス,種:D.ラジオデュラン)のように高放射線環境に適応する細菌も存在する。10グレイ(Gy:1グレイは物質1kg当たりに1ジュールのエネルギーを放射線から受けたということを意味【J/kg)】。)の放射線でヒトを、60Gyの放射線で大腸菌を殺す。Deinococcus radioduransは5,000Gyを浴びても死滅せず、15,000Gyでも37%は生き残る。となれば、放射線に強い好極性微生物の中に含まれ、その強さの鍵となるタンパク質についての分子の立体構造と、その機能発揮に重要な役割を果たすアミノ酸分子を生成できれば、放射線で傷ついた遺伝子を修復するタンパク質を原子レベルで開発し、放射線障害やがん化抑制の医療への開発が期待できる。
Deinococcus radioduransは、一本鎖DNAと二本鎖DNAの両方を修復できる独自の機能を備える。細胞に損傷が生じた場合、損傷したDNAはコンパートメントリング(複数の区画)状の構造に運ばれ、そのコンパートメントの外側から修復されたDNAと内側の損傷したDNAと融合させることができる。しかしながら、この放射線に対する「強さ」を獲得したDeinococcus radioduransが、放射線から身を守り、DNAを修復する優れた能力を発揮するためのメカニズムについては、未だ多くの謎が残されている。これまでの研究から、この細菌にはPprAと呼ばれるその細菌の仲間しか持っていない特有のDNAに結びつくタンパク質が存在することが知られている。このため、まずPprAの立体構造を知ることが課題となっている。
2020年8月、科学者たちは、国際宇宙ステーションInternational Space Station (ISS)で行われた研究に基づいて、地球のバクテリア、特にDeinococcus radioduransが宇宙空間で3年間生存することが判明したと報告されている。これらの知見は、宇宙ダスト・流星体・彗星・隕石・小惑星、そして汚染された宇宙船など、宇宙全体に生命体が分布しているという仮説であるパンスペルミアpanspermia(宇宙播種説)の概念、つまり地球の生命の有機的な構成要素は、地球外、つまり宇宙に起源を持つという説が支持される所以になっている。生命に使用される小さな有機分子の多くが宇宙で発生し、それが地球のような天体に到達し進化する可能性を示唆した。
極限環境微生物には多くの種があり、しかも世界中に生息域を広げている。例えば、地表から奥深くにある熱い岩石の内部に生息するサーモコッカス・ガンマトレランスThermococcus gammatoleransは、超好熱性の古細菌である形態的には1~2μmほどの直径を持つ球菌で、多数の鞭毛を持つ場合が多い。かなり強いガンマ線に耐える種もいくつか含まれている。
極限環境微生物のグラム陰性古細菌gram-negative archaeonであり、現存する最も放射線耐性のある生物として知られている。グラム陰性古細菌は、地球上の生命を支えているあらゆる環境に適応している。細菌は伝統的に、グラム染色反応に基づいて、グラム陽性菌とグラム陰性菌に分類される。そのグラム染色反応は、細菌種をグラム陽性菌とグラム陰性菌の2つの大きなグループに分類するために使用される染色方法で、真菌感染症の診断にも利用されるが、この名前は、1884年にこの技術を開発したデンマークの細菌学者ハンス・クリスチャン・グラムに由来している。
膜が1つしかないグラム陽性菌は単胚葉菌とも呼ばれ、膜が2つあるグラム陰性菌は二葉菌としても知られている。このカテゴリーには、モデル生物の大腸菌や緑膿菌(洗い場などの水回りで増えやすいがどこにでもいる)、クラミジア・トラコマチス(主な性感染症のひとつである性器クラミジア感染症)、エルシニア・ペストなどの様々な病原菌が含まれている。
Thermococcus gammatoleransは、1991年のグアイノート巡航中に、メキシコの西部にある半島バハカリフォルニア半島の沖のグアイマス盆地の水深約2,600mの海底熱水噴出孔から潜水艇ノーチユによって採取された。また、2003年にもグアイマス盆地(北緯27度1分、西経111度24分)の熱水煙突から採取されたサンプルから発見された。Thermococcus gammatoleransは、55〜95°Cの温度で繁殖し、88℃前後が最適な増殖を示す。つまり全種が好熱性、偏性嫌気性の従属栄養生物であり、熱水域でペプチドや多糖類を分解して生活している。水素により強い増殖阻害を受けることから、代謝の結果生じる水素を硫黄Sと反応させ、硫化水素H2Sに.して除去している。このため単体硫黄や硫化金属が多量に存在する環境をよく好む。しかも、放射線に対して最も強い耐性を持つ既知の生物であり、最大30,000グレイGyの線量と最大5,000グレイGyの瞬間線量に耐えることができ、生存能力を失うことはない。
Thermococcus gammatoleransの細胞は、最適な条件と栄養素の欠如により、放射線抵抗性が低下するが、染色体DNA修復システムは、成長期の指数関数的な成長段階にある細胞を、より急速にDNAを再構成する。つまり苛烈な環境下でThermococcus gammatolerans sは、生存能力を失うことなく、損傷した染色体をゆっくりと、時には迅速に再構築することができる。
Thermococcus gammatoleransのDNA修復メカニズムは、より複雑なゲノムに組み込むことで、それを鋳型にしてDNAを修復改善し、細胞の老化を減少させることができる。高温に耐性のある新しい酵素マーカーの開発への応用と、発がん研究およびミトコンドリア疾患の発症の研究への応用について研究が行われている。
目次へ
 |
 |
| ストロマトライト シアノバクテリアが分泌する粘液に細かい堆積物が海水中の炭酸カルシウムとともに沈着する。シアノバクテリアは、日中は光を求めて沈着物の表面に出、夜間は活動を休止する。この繰り返しによって炭酸カルシウムを含む固い層状構造が形成された。 |
保存状態が良好な太古代中期の34億6,000万年前以前のバクテリアと思われる化石が西オーストラリア州のピルバラで発見されている。さらに、同じピルバラで、最も古い生命体とされる、メタン生成菌が約35億年前に存在していたことを示す証拠を、東京工業大学の上野雄一郎グループが発見しており、2006年10月23日、英科学誌ネイチャーで報告している。
ピルバラ地域の地質環境は学術的に見て極めて古いものからなり、化石化したストロマトライトや30億年前まで遡る花崗岩などが存在する。ピルバラ地域の主産業は、鉄鉱石採掘を筆頭に、原油採掘を含む鉱業ならびに産出物の輸送である。オーストラリアの鉄鉱石産出量の大半はこの地域からのものであり、特にトムプライス、ニューマン周辺に産地が集中している。
このオーストラリア大陸北西部のピルバラ地方ハマスレー地域には、太古代後期の27億年前の縞状鉄鉱層が広く分布している。広大な大地一面が、厚さ数100mにもおよぶ厚い赤茶けた鉄鉱層でできている。太古代に、海水中に解けていた鉄分と遊離酸素が結合し酸化鉄になり沈殿してできたものである。地球上に大量の酸素が発生したことを物語る証拠でもある。現代では、大規模な鉄鉱石の産地としても知られている。
現在、リオ・ティント社は、「ピルバラ」のような遠い場所を含む規模の鉄道網を自動運転化し、業務の安全性と効率化をし、200両以上の貨車を機関車で牽引し、16か所の採鉱場から4か所の港湾施設へ、1,700km以上の距離を自動走行して鉄鉱石を運んでいる。
(原文は2020年1月に公開
「日立レールSTS社のプロダクト・マネジャーのロズリン・スチュアートは、「このプロジェクトが成功した理由の1つは、チームメンバーの存在です。現メンバーは、2012年のプロジェクト開始当時からほとんど変わっていないのです」と、振り返ります。リオ・ティント社と日立レールSTS社は今後も、さらなるシステム改善のために協創を続けていきます。」 )
2025年2月19日 、[東京 19日 ロイター] は三井物産は19日、豪州西部ビルバラ地域のローズリッジ鉄鉱石事業の権益40%を取得すると発表した。
温室効果ガスとは、水蒸気・二酸化炭素・メタン・フロン(フッ素Fと炭素Cの化合物の総称)など、赤外線を吸収して温室効果、つまり地上から宇宙方向へと出ていく熱を大気中に留める効果をもたらす気体のことあるが、その一方、二酸化炭素やメタンなどの温室効果ガスは他の気体よりも赤外線を吸収しやすく、しかも太陽光や地上から放射された赤外線を吸収して、その温度が上がった温室効果ガスからもその温度に応じた強さの赤外線を周囲に放射する。
二酸化炭素CO2は2個の酸素原子に1個の炭素原子が結合しているが、メタンCH4は4個の水素原子に1個の炭素原子が結合している。
メタンを研究しているニューヨーク大学の環境科学者マシュー・ハエックは指摘する。「このため、赤外線を受けたり吸収したりする際の原子間の結合による振動や、赤外線を再放出する方法もより多いことになります」
「1ポンド(約450g)のメタンは、排出された時点で二酸化炭素の100倍以上の熱を閉じ込めます」と、だが、メタンのほうが消失が早い。
赤外吸収スペクトルinfrared absorption spectrumは、試料に赤外線をあてて得られる吸収スペクトルを測定する分析手法である。原子同士の結合は硬く固定されているわけではなく、バネで繋がるかのように、ある程度の柔軟性をもっている。
 およそ有機化合物と呼べるものは、すべからく赤外領域に固有の吸収スペクトルを持っている。分子に赤外領域のエネルギーを与えると、そのエネルギーを吸収して化学結合間に振動が生じる。このとき、吸収された赤外線エネルギー量を測定することで、化学結合の種類などを知ることができる。
およそ有機化合物と呼べるものは、すべからく赤外領域に固有の吸収スペクトルを持っている。分子に赤外領域のエネルギーを与えると、そのエネルギーを吸収して化学結合間に振動が生じる。このとき、吸収された赤外線エネルギー量を測定することで、化学結合の種類などを知ることができる。赤外吸収スペクトルのピークをみても、大まかな構造式が分からない。少なくとも、赤外分光法だけで構造式を推測するのは不可能である。大まかなデータしか得られないという欠点からも、正確な分析手法としては重要度は低いが、分子の振動や回転の状態を変化させるのに必要なエネルギーは、物質の化学構造によって異なり、物質に吸収された赤外光を測定すれば、蓄積されたデータと比較して化学構造や状態に関する情報が得られる。
二酸化炭素は温室効果ガスと呼ばれて、地球温暖化の原因物質と言われている。これは、二酸化炭素が赤外線を吸収するからで、赤外線(光粒子)は物質を温める作用があり、物質に光(電磁波)を当てると暖かくなるのは、吸収された光粒子が分子の振動や回転を活発化する熱エネルギーに変換しするからである。また二酸化炭素が増えるほど地球温暖化が進行するのは、二酸化炭素が赤外線を吸収すると分子が活発に動き回り、周辺空気全体を温めるからである。
酸素や窒素ではなく、なぜ二酸化炭素が温室効果ガスなのかについて、赤外線の性質や分子が振動する原理によって理解できる。実際には未知化合物の構造決定で赤外分光法を利用することはほぼないが、既知物質を解析するときには有用である。
あらゆる物質で単結合を有し、単結合では500~1500cm-1(センチメートルのマイナス1乗と読む)でピークが見られる。このエリアは単結合によるあらゆるピークが混ざり合い、かなり複雑なピークとなっている。また、わずかに構造式が違うだけでもピークの形が変わる。
(500~1500cm-1の場合の赤外光の波長𝜆は、0.00067~0.002 cm【6.7~20 μm】 の範囲になる。赤外線分光の分野では、この範囲が分子の振動による特有の吸収を示す重要な領域とされている。)
ヒトの体に赤外線を当てると暖かくなるのは、赤外線のエネルギーが吸収され熱に変換されるからである。あらゆる分子は単結合を有するが、酸素や窒素、水素など左右対称の分子は赤外線を吸収しない。つまり、赤外吸収スペクトルが観測できない。
それでも水素を含む単結合のピークは、赤外吸収スペクトルでは、水素を含む単結合は2500cm-1(0.0004 cm=4 µm))以上に伸縮振動のピークが観察される。以下の結合に関するピークがこのエリアで見られる。
C-H
O-H
N-H
これら水素を含む結合の中でも、重要なのが3000cm-1(0.000333 cm=3.33 µm)以上にあるピークで、赤外吸収スペクトルでは、3000cm-1の付近に着目する。それ以外でも物質であれば、その多くが赤外線を吸収するため、ピークを観測できる。これら単結合のピークは500~1500cm-1(0.00067~0.002 cm=6.7~20 μm)でピークが見られる。
アミノ基の-NHについても、3300cm-1 (0.000303 cm=3.03 µm)の周辺にピークが表れる。ヒドロキシ基の-OHとアミノ基の-NHはどのように見極めるのかは、-NHのほうがピークの形が鋭くなり、見分け方はこれだけである。また-NH2のように、窒素に水素原子が2つある場合、3300cm-1の周辺に2本のピークが表れる。
赤外吸収スペクトルでは、1300cm-1 (0.000769 cm=7.69 µm)以下の部分を指紋領域と呼ぶ。人間は固有の指紋を有している。人によって指紋の形が異なることから、犯人を捜すときに指紋を調べる。指紋と同じように、化合物ごとに独自のピークの形があるので、既知化合物については、指紋領域を確認することで同じ物質かどうかを判別できる。
実際には未知化合物の構造決定で赤外分光法を利用することはほぼない。しかし既知物質を観察するときに極めて優れている。
大気中にとどまっている期間は、メタン.はわずか10年ほどで消滅する。、一方、二酸化炭素は数世紀にわたって大気中にとどまり累積される。
実は、メタンガスは、二酸化炭素の約25倍の温暖化効果があり、太古代終期の25億年前に地球の寒冷な気候を緩和し生物が生息しやすい環境を作ることに貢献していた。
(「地球温暖化係数Global Warming Potential」の指標は、CO2を基準にして、ほかの温室効果ガスがどれだけ温暖化する能力があるか表した数字で、地球温暖化係数として現時点での数値が報告されている。CO2:1、メタン:25。)
天然ガスの主成分はメタンCH4であるため、石炭や石油などと同じく化石燃料であるが、一定量のエネルギーを生み出す際に排出されるCO2は、石炭や石油に比べてる格段に低い。近年、火力発電において石炭火力が忌避され、天然ガス火力への移行が進む理由はまさにこの点にある。しかも、天然ガスはその名の通り常温で気体であるから、陸上ではパイプラインが使える。しかし、日本のように海外から輸入する際には、輸送時の体積を軽減するために液化が必要になる。液化天然ガスLiquified Natural Gas(LNG)として輸送する。LNGとよく似た略称に液化石油ガスLiquified Petroleum Gas(LPG)がある。LPGはプロパンC3H8とブタンC4H10を液化したものであり、プロパンボンベとして流通してる。LPGが常温で液化するのに対してLNGではー160℃という極低温が必要になる。これが天然ガスをLNGとして輸送する際のコスト増の要因となっている。 メタンは実は身近なガスでもあり、有機物が発酵する際に生成するありふれたガスと言える。牛のゲップもその一例であり、これを意識的に利用し、生ごみなどの有機廃棄物の発酵処理によりメタンを生成させ、廃棄物削減と同時に燃料回収を達成するバイオガスのプロジェクトが日本各地で進行している。水には溶解せず、ガスとして容易に回収できるというメタンの特性を活かした試みでもある。当然、大気中に放散させない注意が必要となる。
上野雄一郎グループは、西オーストラリア州のピルバラの約35億年前の古い地層で、石英に閉じ込められていた気泡の中に生物が作ったメタンがあることを突き止めた。上野雄一郎グループはあきらめずに研究を続け、西オーストラリアのピルバラ地塊で得られた約35億年前の熱水沈殿にメタンを含んだ流体包有物の痕跡を抽出した。流体包有物とは、鉱物が熱水から沈殿して成長していく際に周りの熱水から取り込んだもので、鉱物が形成された当時の環境を推測するために分析したりする。その分析により炭素同位体組成により、このメタンが微生物起源であることが解析された。これは、メタン細菌の存在を示す最も古い痕跡となり、これまでの地球化学的な状況証拠より約7億年も遡る。
その石英は、地下の.熱水が岩盤を上昇し海底に達した時に溶けていた成分が結晶化してできた。炭素には原子量が12Cと13Cの2種類があり、生物は軽い12Cの方が取り込みやすいため、メタンCH4の炭素を調べると生物が作ったものかどうかがわかる。このメタンは海底の熱水中の菌が作った。
太古代では、既に原始微生物による代謝過程の多様性が気候変動に重大な影響を及ぼしていた。太古代の海洋において、水素と鉄という異なる電子供与体を利用する複数種の酸素非発生型光合成細菌が「共存」することによってはじめて、大気中に高濃度のメタンを維持することが可能になった。ここでいう光合成は、より原始的な酸素非発生型の代謝過程を踏まえ、水の水素を電子供与体とする酸素発生型の光合成とは異なり、酸素非発生型の光合成は水素分子や2価鉄Fe2+、硫化水素H2Sなどの物質を電子供与体として利用する。2価鉄イオンFe2+は2つの電子を失い、2価の鉄イオンとなり水に溶解するため、3価鉄イオン3+よりも酸化・還元反応に対して反応性が高い。
硫化水素は好気性生物の多くにとっては有毒であるが、酸素非発生型光合成、すなわち水素源として水ではなく硫化水素を用いることで、酸素ではなく硫黄を副産物として放出する。硫黄酸化細菌などの化学合成細菌は硫化水素の酸化に伴い発生するエネルギーで炭酸同化を行う。後者は硫化水素の豊富な海底火山の熱水噴出孔付近で生産者の役割を担い、独自の生態系を形成している。滑走運動をする硫黄細菌の一種で化学合成独立栄養生物であるが、炭素固定に必要な酵素が不足しているため有機化合物を利用する混合栄養の株が多い。電子供与体として硫化水素のほか、チオ硫酸イオンも利用できる。
酸素非発生型光合成を行う生物としては、紅色細菌・緑色硫黄細菌・緑色非硫黄細菌・ヘリオバクテリアなどの光合成細菌などが知られている。同じバクテリアでも、シアノバクテリアは、酸素発生型の光合成生物である。
(硫黄酸化細菌が、硫化水素を酸化する反応式は
H2S+1/2O2=H2O+S+176kJ
このとき発生するエネルギーで炭酸同化すなわち化学合成を行う。上記のような化学合成生物である硫黄細菌のほかに、光合成細菌である緑色硫黄細菌・紅色硫黄細菌などを含めて硫黄細菌とよぶ場合もある。 )
地球史前半、先カンブリア紀原生代の約20億年前までは、太陽は現在より20~30%も暗かったことから、地球の大気組成が現在と同じであると仮定すれば、約20億年前より以前の地球は全球凍結状態であったと考えられる。しかし、実際にはそのような地質学的根拠が見当たらず、むしろ太古代の地球は現在よりも温暖だったようで、それと矛盾する。むしろ、過去の大気二酸化炭素濃度が現在よりもずっと高く、その強い温室効果によって温暖環境が維持されてきたと考えられる。その一方、地質学的調査によれば、約22億年前以前の大気二酸化炭素濃度は理論予測よりも低く、二酸化炭素だけでは温室効果が足りない可能性が指摘されている。二酸化炭素の温室効果を補うものとして、メタンが有力視されている。それ以外にも様々.な温室効果気体種やメカニズムが提案されているものの、未だに完全な回答は得られていない。しかも、メタンについても、温室効果として必要な濃度の推定はなされているものの、その濃度が本当に実現できるのかどうかは分かっていない。
極限性微生物
生命の最も古い痕跡は約38億年前のグリーンランドの堆積岩のものだが、どのような生物が作ったのかわかっていないが、東京工業大学地球惑星科学部教授 上野雄一郎は「昔の地球がどのような気候だったのか考える手がかりになる」と話している。
オーストラリアのピルバラ地方の地層から、2002年ごろから、生物の化石を発見したという報告があがる。しかし、一部の専門家から岩石が鉱化した跡にすぎないのではないかとの意見が出され、10年近く議論が続いていた。
その太古代中期の約34億6,500万年前の地球最古の化石は微生物マット(固体や液体の表面に付着した微生物が形成する生物膜)の形をしていた。その微化石は、石英砂粒から発見された。ピルバラ地方には、地球がまだ水の惑星で、海水が風呂のお湯ほどの温かさだったと考えられている。その34億6,500万年前の先カンブリア紀太古代の地層の岩石には、かつて微生物が含まれていたという証拠があり、これが地球上の生命の最初の直接的な証拠と言われている。
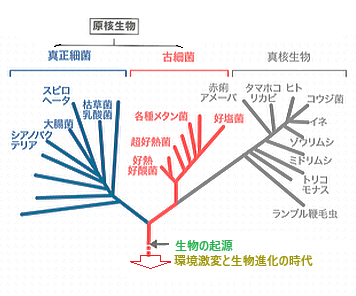 |
| 生物を基本的な遺伝の仕組みや生化学的性質を元に分類する『3ドメイン説』では、生物全体を3つのドメインに分け、これらドメインよりも下位の分類階級として「界」と言う用語を使う。 生物全体を「真正細菌ドメイン」「古細菌ドメイン」「真核生物ドメイン」に3分する。 「真正細菌ドメイン」と「古細菌ドメイン」に属する生物は「原核生物」、「真核生物ドメイン」に属する生物は「真核生物」で、「原核生物」とは真核、つまり明確な境界を示す核膜を持たない細胞からなる生物であり、すべて単細胞生物である。 真核生物は、核をもつ細胞で、DNAが核膜に包まれている細胞(真核細胞)をもつ生物を指す。 『3ドメイン説と5界説』によれば、動物界・植物界・菌界・原生生物界・原核生物界((モネラ界)に分けられる。原生生物界は、原核生物ではなく真核生物で、「菌界」「植物界」「動物界」にも属さない例外的な存在で、アメーバ・ゾウリムシ・キイロタマホコリカビなどが含まれる。 原核生物界は、真核生物と対をなす分類で、性質の異なるバクテリア(真正細菌)とアーキア(古細菌)の2つに分けられるが、真正細菌より古細菌の方が真核生物に近いことが判明している。 真正細菌は、単細胞生物の中でもより複雑な領域で、土壌や水、大きな生物の内部や外部など、地球上のほとんどの生息環境に適応している種であつ。 真正細菌は膜に結合した小器官を持たないため、ほとんどすべての代謝反応は細胞質内で行われる。 一部の真正細菌は窒素循環にも関与している。 また、宿主生物に寄生したり、病原性を示したりすることもある。 古細菌は極限環境に生息する単細胞の微生物で、地球上に最初の生命が誕生した直後に進化したと考えられている。 そのため、古代細菌と呼ばれる。 古細菌は、温泉・塩湖・海・湿地・土壌などに生息し、人間の皮膚や口腔、大腸にも生息している。 古細菌は、炭素循環や窒素循環に重要な役割を果たしている。 しかし、その病原性や寄生性はいまだに確認されていない。 古細菌には、メタン菌や好塩菌、好熱菌の3種類がある。古細菌と真正細菌の大きな違いは、この環境中での生息場所にある。 メタノゲンは、沼地や湖の堆積物、動物の消化管などの無酸素環境に生息し、メタンガスを発生させる。 好塩菌は高濃度の塩分を含む水に生息している。 好熱菌は、酸性硫黄泉などの熱水環境に生息する。 |
この太古の微生物は硫黄化合物を食べて生きていたとみられる。化石のサイズは10㎛程度だがその化石は微生物の細胞の形状をしており、さらに鉄と硫黄の化合物、黄鉄鉱の微細な結晶も発見された。これは、生物が硫黄などを代謝したことによる副産物であることは明らかだと研究チームは言う。硫黄を食べる微生物は、現在でも悪臭を放つ水路や土壌などに生息しており、海底熱水鉱床に生息するものは「極限性微生物」と呼ばれている。
約34億6,500万年前にピルバラ地域から産出した細胞の最古の化石は、無機的な化学反応で岩石に生じた粒を誤認したとかつては批判されたが、3Dイメージング技術の改良を経て内部構造が可視化され、確実な化石証拠となった。
マーティン・デイビッド・ブレイジャーMartin David Brasier(1947年4月12日~2014年12月16日)は、先カンブリア紀とカンブリア紀の微化石と進化の概念分析で知られるイギリスの古生物学者・宇宙生物学者であるが、教授は「ついに34億年前の生命の動かぬ証拠を手に入れた。当時のバクテリアは酸素なしで生きていたことを示している」と声明を発表した(2011年08月22日 AFPnews)。
海洋植物が枯れた後、バクテリアがその植物を分解するプロセスは非常に重要である。海洋植物が枯れると、表面に生息するバクテリアや菌類が活動を開始する。これらの微生物は植物の細胞壁を分解し、原形質を放出させる。次の中間分解では、より大きな微生物が活動を開始する。これらの微生物は、初期分解によって放出された栄養素を取り込み成長し、さらに植物の分解を速める。最終分解の段階では、大型の分解者、例えば、一部の昆虫や甲殻類が活動を開始する。これらの生物は、微生物が分解した残り物を消費し、さらに分解を促進する。海洋植物はバクテリアや他の微生物によって徐々に分解され、海洋環境に栄養を補給する。
太古代初期の約38億年前の原始の海において、非常に単純な単細胞の原核生物が初めて誕生した。その海底に出現した最初の生物は、きわめて単純な構造の、核膜を持たない単細胞の微細な原核生物だ。核膜に囲まれた細胞核を備えないが、細胞内にDNAが散在しているわけではなく、「核様体Nuclear body」と呼ばれるDNAの凝集体を構成している。原核生物には、ミトコンドリアやゴルジ体などの細胞内小器官を持たない。原核生物の多くは細胞壁を持ち、細胞の形状と保護を担う。細胞壁の成分は糖鎖(糖が複雑に連なって形成する一群の化合物)とペプチド鎖(アミノ酸がペプチド結合により短い鎖状につながった分子の総称)の2つの主要な成分から構成されているペプチドグリカンである。球菌(ブドウ球菌)・桿菌(形状が細長い棒状または円筒状を示す細菌や古細菌【大腸菌】)・らせん菌(桿菌の中にはらせん状にねじれたものがある【カンピロバクター】)など、さまざまな形状のものが存在し、その形状は生活環境に適応していて多様である。
海底火山からの熱を取り込み、海水に溶けている有機物を体内に取り込んで生育していた好熱酸性古細菌Thermoacidophilesは、高温かつ酸性の環境に適応し、温泉や火山地帯、深海熱水噴出口などに生息する。この単細胞の微細な生物のDNAは核膜がなく細胞内に浮遊している「核様体」のため、DNAが損傷する危険性が高い。
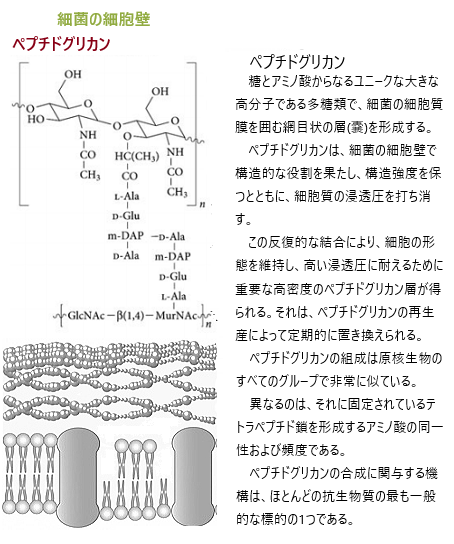 シアノバクテリア
シアノバクテリア 原核生物には性がないが、数分ごとに分裂して自己を複製するた。数秒でクローンを作るものもあった。この時代では、現在の地球とは異なり、地表付近には、酸素O2分子は存在せず、もしくは、存在しても極めて微量であった。しかも、生物に有害な紫外線が太陽から地表に大量に放射していた。しかし水深の深い海では太陽からの有害な紫外線は届きにくい。
その後、太古代終期の約27億年前頃から地球表層に酸素が存在するようになる。この酸素を作り出したのは海水中に生息する各種シアノバクテリアは、光合成を行う細菌で、太陽からの光エネルギーを使って、二酸化炭素と水から有機物と酸素を合成した。
シアノバクテリアは、この酸素光合成によって生物学的エネルギーを得ることができるシアノバクテリオタ門の独立栄養グラム陰性菌のグループにある。シアノバクテリアの名前は、青みがかった緑色(シアン)を指しており、シアノバクテリアの非公式の一般名である藍藻類の基礎を形成しているが、原核生物として、科学的には藻類には分類されていない。
シアノバクテリアは、最も多数の分類群であり、酸素を生成した最初の生物である。太古代中期に出現し、太陽光の赤と青のスペクトル周波数を吸収し、緑がかった色を反射する。水分子を水素イオンと酸素に分割し、その水素イオンは二酸化炭素と反応して炭水化物などの複雑な有機化合物を生成する(炭素固定)。その副産物として酸素を放出する。
シアノバクテリアは、何十億年にもわたって酸素を絶えず生成し放出することにより、酸化性の大気に変換した。原生代には、地球上の生命体の組成を劇的に変えた。原生代には、地球上の生命体の組成を劇的に変えた。その後、初期の単細胞生物が酸素環境で生き残るための適応が、シアノバクテリアを細胞内へ取り込むことでであった。嫌気性菌と好気性菌の間の内部共生であり、それが原生代の真核生物の進化に繋がった。
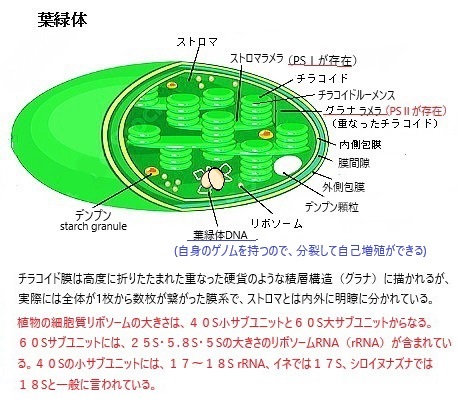 シアノバクテリアは、様々な形態のクロロフィル・カロテノイド・フィコビリンなどの光合成色素を使用して、太陽光の光エネルギーを化学エネルギーに変換した。寄生型の従属栄養原核生物とは異なり、シアノバクテリアは内膜を持っている。それが、光合成が行われるチラコイドと呼ばれる平らな嚢である。紅藻類・緑藻類・植物などの光独立栄養性真核生物は、いずれもシアノバクテリアを祖先とするクロロフィル細胞小器官で光合成を行っている。その後、真核生物のこれらの内部共生シアノバクテリアが進化を.重ね、葉緑体・色素体・エチオププラスト・ロイコプラストなどの特殊なオルガネラに分化した。
シアノバクテリアは、様々な形態のクロロフィル・カロテノイド・フィコビリンなどの光合成色素を使用して、太陽光の光エネルギーを化学エネルギーに変換した。寄生型の従属栄養原核生物とは異なり、シアノバクテリアは内膜を持っている。それが、光合成が行われるチラコイドと呼ばれる平らな嚢である。紅藻類・緑藻類・植物などの光独立栄養性真核生物は、いずれもシアノバクテリアを祖先とするクロロフィル細胞小器官で光合成を行っている。その後、真核生物のこれらの内部共生シアノバクテリアが進化を.重ね、葉緑体・色素体・エチオププラスト・ロイコプラストなどの特殊なオルガネラに分化した。(初期の地球の大気中の遊離ガス状酸素は、様々な表面還元剤によってすぐに除去されていた。)
光合成を行う微生物の中には、大きなコロニー(生物集団)を形成したものもある。それが化石化したストロマトライトstromatolite(微生物の死骸と泥粒などでつくられる堆積構造の岩石)は、大きいものは高さが1mを超す。西オーストラリア州のシャーク湾にあるこのコロニーの化石は、およそ30億年前に形成されたものだ。ストロマトライトは、古代ギリシャ語 のστρῶμα(strôma)は「層、地層」、およびλίθος(líthos)「岩」が語源でとなる。
地球上で最初に誕生した生命体のひとつであるシアノバクテリアや硫酸塩還元細菌、シュードモナドタseudomonadota(粘液細菌、土壌細菌のひとつで、多細胞的な行動を起こすグラム陰性のバクテリア)などの光合成微生物が主として作り出す微生物堆積層で、これらの微生物は、浅い水域の砂や泥の表面に定着して光合成を行い、副産物として酸素や粘液を分泌し、その粘液は砂や泥の粒子を捕捉して固定し鉱物の「微生物マット」を形成した。日中に光合成を行い、夜間に休止するというサイクルを繰り返すことで、シアノバクテリアは上部へと成長し、新たな層を形成する。この過程が繰り返されることで、炭酸カルシウムなどの鉱物が沈殿し、ストロマトライトの層内に堆積した。これにより、ストロマトライトの特徴的な積層を生成した。この層状構造は、数千年にわたる微生物の成長と進化の環境過程を反映している。しかも、肉眼では見えない小さな微生物が作り出した層状の岩のような構造物の一部には、そのフィラメントの中に解析するに値する堆積物が閉じ込めている。
本質的に、酸素は強い酸化力を持つ毒性の高い気体であるが、この時代以降、シアノバクテリアを含む一部の生物は、この環境に適応し酸素を利用した代謝機能を持つように進化した。酸素呼吸は、グルコースなどの有機分子を完全に分解し、二酸化炭素と水にすることにより、エネルギーを最大限に利用できる。これにより、一分子のグルコースから約38分子のATP(アデノシン三リン酸)が生成される。これは嫌気呼吸に比べて大幅に生産性が高い。シアノバクテリアが作った酸素は海水に溶け込むとともに、その一部は原始の海から大気に拡散した。そして、原生代初頭の約23 億年前以後、原始大気中にも酸素が多量に存在するようになると、生物に有害な太陽からの紫外線の多くは、酸素と紫外線エネルギーにより生成されたオゾンO3によって遮断され、地表に届かなくなり始めた。
 一般に大気中での紫外線照射によりオゾンは発生する。
一般に大気中での紫外線照射によりオゾンは発生する。3O2 ⇒ 2O3
またオゾンは不安定な分子であるため、放置しておくと以下の化学式で酸素に変化する。
2O3 ⇒ 3O2
この反応は温度や圧力が上昇するほど速くなる。
さらに大気中の酸素やオゾンの濃度が増加した原生代終期の約4億年前になると、海洋生物は、陸上に進出し始め、急速に陸上生物として目覚ましい進化を遂げた。現在、海に生息する植物プランクトンが光合成で作り出す正味の有機物の量は 地球規模で年間約50ペタグラムPg(ペタpetaは1015乗、即ち千兆)と報告されている。正味の有機物の量という意味は、植物プランクトンが増殖する際、呼吸により光合成で作り出した有機物と酸素を使 って活動エネルギーに変えることから、光合成で作られた有機物から呼吸でエネルギーとして消費された有機物を差し引いた値と言う意味である。
地球上で全陸上植物が光合成で作り出す正味の有機物の量は1年間に約 60 Pg(ペタグラムpetagram )と言われている。つまり、植物プランクトンが作り出す正味の有機物の量は陸上植物にほぼ近い。このため、海に生息する植物プランクトンは、陸上植物と同様、地球温暖化の原因である二酸化炭素を有機物に固定する重要な役割を果たす。
海洋表層で植物プランクトンによって作られた有機物の多くは、植物プランクトンを捕食する動物プランクトンや海水中の有機物を餌として増殖するバクテリアなどの呼吸により、海洋表層で分解され、二酸化炭素に戻されている。しかし、その一部は、分解されずに水深 200 m以上の中深層とよばれる深い水深に輸送される。一度、有機物が中深層に輸送されると、千年スケールで海洋表層に二 酸化炭素の形で留まることが知られている。この海洋表層(有光層)から海洋内部へ生物学的に炭素を輸送する経路を「生物ポンプBiological pump」と生物海洋学では呼んでいる。
海洋表層の植物プランクトンが死滅して、生物ポンプが働かなくなったとしたら、現在の大気中の二酸化炭素濃度は約3倍増加すると見積もられている。近年、植物プランクトンの増殖に必要な主要栄養物質の窒素やリンなどは豊富に存在するが、鉄が不足していると考えられている。そのため東部赤道太平洋域や北太平洋亜寒帯域、そして南極海域の表層に人為的に鉄を散布して、生物ポンプを促進するための中規模ながら鉄散布の実験が、世界各国の多数の海洋学研究者により実施されている。鉄は地表付近に存在する全元素の重量比で約5%を占め、4 番目に多い元素であるが、陸地から遠く離れた外洋域の表層海水中には極めて微量にしか存在していない。これは、約27億年前にシアノバクテリアが酸素を作り出した結果、海水中に酸素が多く溶け込み、そのほとんどの鉄粒子が酸化鉄として沈殿し、また数多くの塩類を含む海水が弱アルカリ性になった結果、海水には鉄がほとんど溶けることができなくなった。
物質がpH7である場合、これは中性であることを意味する。海洋の塩水やその他の自然環境は、様々な要因によって変化する。現在の海洋の表面付近の平均pHは約8.1である。これは、海洋がアルカリ性であることを示す。しかしpHは、通常、微妙なバランスで保たれる。例えば、ヒト血液は、7.35〜7.45のpH範囲にある。それは、わずかに変化しても、体内に損傷を生じさせる可能性がある。海洋における塩水の酸性化の主な効果の1つは、サンゴ礁にある。サンゴは、骨格を強く保つために炭酸カルシウムを吸収する必要がある。海はいま、しだいに酸性化している。海はもともと弱いアルカリ性だが、大気中に増えた二酸化炭素が海面を通して余計に溶け込むと、そのアルカリ性が弱まる。これが海の酸性化だ。海洋が酸性に変わると、サンゴ礁は溶解し死に絶える。ハマグリや.ウニなども炭酸カルシウムが不可欠であれば貝類も同様である。より多くの酸性海洋がこのような変化に耐えることのできない多くの海洋生物を死滅させ、地球の海洋全体の生態系を大きく変化させる。
海水のpHは一般的に弱アルカリ性を示し、表面海水では約8.1、深くなるにつれてpHは下がり、北西太平洋亜熱帯域では水深1,000m付近で約7.4と最も低くなる。 これは、深くなるにつれて有機物の分解により海水中の酸素が消費され、全炭酸濃度が増加することによる。これは、深くなるにつれて有機物の分解により海水中の酸素が消費され、全炭酸濃度が増加することによる。人為起源二酸化炭素の蓄積だけでなく有機物分解の影響を指摘されている。2009年には、既にハワイ近海では水深約250mでのpHの低下速度がもっとも速いことが報告されている。その後、大西洋においても海洋内部でのpHの低下が報告されており(2012年・2010年・2009年)、海洋酸性化は表面海水中だけでなく海洋内部においても世界的に進行している。
二酸化炭素が多く溶け込むとpHが下がり、海水のアルカリ性が弱まる。海洋酸性化の指標として用いられるpHは、水素イオン濃度の逆数の対数で定義される値であり、水素イオン濃度が増えるとpHは下がる。
太平洋のハワイ近海では、表面海水中の二酸化炭素濃度の長期的な上昇と、それに伴うpHの低下が報告されている。南大洋・地中海などでも、表面海水中のpHの低下傾向が報告されている。このように海洋のpHが長期にわたって低下する現象を「海洋酸性化」と呼んでいる。
有機物の海水による分解
近年増加する海岸浸食や永久凍土の融解、海洋生物由来の有機物の沈降により、海底に沈積する有機物量が増えている。また、海洋や沿岸地域では、有機物、例えば動植物の遺骸や排出物が、細菌や微生物によって大量に分解されいる。これらの分解プロセスでは、「炭素C」を含んでいる有機物が無機化される際に二酸化炭素CO2などの気体が発生する。このプロセスは自然界の炭素循環の一部であるが、これが過剰に起こると海洋環境に影響を及ぼす。
また森林土壌などでは複雑な組成を持つ有機酸(腐植酸)を含むので土壌は酸性化する。その有機物は酸性を示す有機化合物であれば、その供給が増えれば分解される低酸素化により更に酸性化が進行する。
二酸化炭素は海水に溶けるとすぐにそのほとんどが炭酸水素イオンHCO3-や 炭酸イオンCO32-といった陰イオンの状態になる。二酸化炭素そのままの状態で存在するものと、これらのイオンの総濃度とを合わせて全炭酸濃度と呼ぶ。この全炭酸濃度の観測により大気から吸収した二酸化炭素がどの海域にどの程度蓄積しているかを知ることができる。海水中の炭酸水素イオンや炭酸イオンは、海水を強酸性にするが、二酸化炭素は気体のままに留まる。
大気のCO2濃度(分圧)が海洋のCO2分圧より高ければ大気のCO2は海水H2Oに吸収され、低ければ海洋から大気にCO2が放出される。また海水もCO2を吸収すると、水中で反応して炭酸H2CO3を生成する。この炭酸はさらに解離して炭酸水素イオンHCO3-や炭酸イオンCO32-を生成して水素イオンH⁺を生じさせる。この水素イオン濃度の増加がpH値を低下させ、海水を酸性化させる。
その化学式は、
① CO2 + H2O ⇄ H2CO3 (炭酸)
② H2CO3 (炭酸) ⇄ H⁺ + HCO3- (炭酸水素イオン)
③ H⁺ + HCO3-(炭酸水素イオン) ⇄ 2H⁺ + CO32-(炭酸イオン)
海水中には、炭酸カルシウムCaCO3のように、過飽和の成分も含まれている。長期間硝瓶に入れて置くと、必ず瓶の内壁に.炭酸カルシウムが析出する。産業革命以前、人間活動の影響がまだ少なかった海では、カルシウムイオンCa²⁺や炭酸イオンCO32-が十分に存在し、サンゴや貝が自分の殻を作るのに必要な炭酸カルシウムを簡単に作ることができた。ところが、人間が二酸化炭素を大気中に排出し、海水に二酸化炭素 CO2が溶け込むと、海洋酸性化という現象が起こる。
上記①の式のように、二酸化炭素が水に溶けると炭酸を生成し、これはさらに上記③の式のように、炭酸水素イオンHCO3-や水素イオンH⁺に分解される。この水素イオンが炭酸イオンCO32-と結びついてHCO3-(炭酸水素イオン)となるため、炭酸カルシウムCaCO3の生産を減少させ、貝殻やサンゴのような炭酸カルシウムを基盤にした生物に影響を与える。この影響により、海洋の生態系全体が変わり、例えば、サンゴ礁の崩壊や、海洋の生物多様性の減少に繋がりかねない。
これを「海の酸性化ocean acidification」と呼ぶ。海の酸性化が進むと、カルシウムイオンCa²⁺と結合できる炭酸イオンCO32-の濃度が減少し、生き物たちにとっては成長に必要な炭酸カルシウムCaCO3を作りにくくなる。これが海の酸性化が生物に影響を及ぼすメカニズムである。
炭酸カルシウム CaCO3は水に不溶であるが、二酸化炭素と水に還元され炭酸水素カルシウムCa(HCO3)2が生成され、これにより水に溶けるようになる。
CaCO3 + CO2 + H2O → Ca(HCO3)2 (炭酸水素カルシウムCalcium hydrogencarbonate)
有機物が海水による分解でCO2の放出が海水中に蓄積されると、そのCO2が酸性化を引き起こす。酸性化された海水は、サンゴや貝殻生物にとって有害であり、これらの生物が炭酸カルシウムを形成する能力を低下させる。
雨水H2Oに溶解した二酸化炭素CO2が石灰石の炭酸カルシウムCaCO3と反応して可溶性の炭酸水素カルシウムCa(HCO3)2を形成する。この可溶性化合物は、雨水で洗い流される。この形態の風化を、炭酸化と呼ぶ。
海の中のバクテリア
海は、二酸化炭素の巨大な貯蔵庫だといわれている。海水中の二酸化炭素を活用して、植物プランクトンは光合成を行い、糖などを含む多様な有機物を生産している。その植物プランクトンは、動物プランクトンや魚などに食べられる。その生物の死骸や排泄物は、バクテリアなどによって分解され、二酸化炭素に戻される。その一方で、分解されなかった死骸は、マリンスノーとなって沈下し、海底に降り積もっていく。
海水中の有機物としては、上記の光合成産物のほかに、溶存態有機物Dissolved organic matter(DOM)と呼ばれるものがある。溶存態有機物がもつ炭素量は、陸上植物の炭素量に匹敵し、地球上で重要な炭素の貯蔵庫としての役割をもっている。その溶存態有機物も植物プランクトンの光合成を出発点としている。光合成によって作られた溶存態有機物の多くは、バクテリアに取り込まれて分解される。ところが、海水中に存在している溶存態有機物の大半は、バクテリアにも利用できない。この利用されない有機物が、数百年から数千年もの間、海水中に大量に滞留している。しかも、これを作っているのはどの生物なのかさえ分かっていなかった。現在ようやく、これもバクテリアの代謝産物らしいことがわかってきた。
溶存態有機物(DOM)は、天然水や廃水中に存在するさまざまな有機物の総称であり、フミン物質はその中の主要な成分の一つである。
その起源は、両者とも自然環境の中で生成されるが、フミン物質は、特に、動植物やや微生物の残存物が脱水や分解など物理的化学的な変化によって生成される多種多様な有機物の混合物であれば明確な化合物として識別が困難な物質の総称と言える。その分解産物から生成される高分子量の有機化合物は、溶存態のフミン酸・フルボ酸、および腐植質などから生成されている。そのため、高い分子量と複雑な化学構造を持ち、これにより、DOM全体の化学的および物理的特性に影響を与える。
従って代表的な難分解性溶存有機物とされているフミン物質は、長年研究が行われているが、分かったことはそれほど多くはない。フミン物質は微生物によっても分解されにくく、構造的に非常に安定し極めて長い期間、水中に残存し続ける。フミン物質の性質を決めている核の部分の構造は変わらないが、外側の殻を形づくる組織は少しずつ構造を変化させている。分解して栄養素として取り込もうという微生物がとりついていても、利用するのは困難とまで言われている。
難分解性溶存有機物の典型とされ、天然水中の溶存有機物の30〜80%を占めるといわれるフミン物質は、疎水性の有機酸で、分子量の大きいフミン酸と低分子のフルボ酸からなる。植物などが微生物により分解されて生成し、水田には主に外から入る。水と水田から出る水の量をチェックしながら難分解性溶存有機物を測定すると、水田からフミン物質は出ていないことがわかった。田植えから水がなくなるまで1年間の物質収支を算定すると、水田から出る有機物にフミン物質は少ない。逆にフミン物質は水田に吸収され蓄積され、出水されるのは親水性の酸であった。
湖水や下水処理水はフミン物質が思っていたより少なく、親水性酸が卓越していた。また流入する河川水については、河口付近の水は親水性酸とフミン物質が同程度であったが、上流地点ではフミン物質の割合が高かったと言う。さらに、サンプルごとの類似性を調べて見ると、湖水と下水処理水が似ていたようだ。組成として主なものは双方ともに親水性酸であった。これまで調べた琵琶湖・手賀沼・諏訪湖・十和田湖など、.どこでも同じであった。東京湾でも同様であった。フミン物質が高濃度で存在する方が、逆に非常に希で、しかもpHが低い湿原や泥炭地の水などに限られる。
難分解性有機物は、浄水処理過程で生成される発がん性物質であるトリハロメタンなどの原因物質である。トリハロメタンは、クロロホルムをはじめとする4種類の化合物の総称で、発がん性が指摘されている。そのため湖沼が水道水源になっている場合、水道水についての健康リスクが懸念される。生成されるその量は有機物量および塩素量にしたがって増加する。水道の原水に含まれるフミン物質などの有機物と、浄水処理過程で用いられる塩素が反応して生成されることから、水道水の水質基準が設定されている。フミン物質の研究は、20世紀から今日まで行われてきたが、その構造や機能などの全体像は未だ解明されていない。
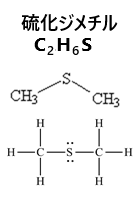 |
| 「磯の香り」の本体である硫化ジメチルDMSは、海洋から大気に放出され、大気中で硫酸エアロゾルとなり雲核として働く。 その主な発生源は、海藻や植物プランクトンが生成するジメチルスルホニオプロピオネートDMSPで、海洋細菌の分解作用によってDMSに変換される。 |
硫化ジメチルは海のプランクトンにより作られ、磯の匂いの元となる物質でもあるが、海水に溶けにくいため、溶けきれなかった大量の硫化ジメチルが大気中に放出され、酸化されて、やがて硫酸塩(硫酸イオン SO42-を含む無機化合物の総称)に変化する。この硫酸塩が雲の元となる粒子になり、雲の形成につながるといわれている。海洋の小さい微生物によって作られる有機物が、地球の気象に影響を及ぼしていた。
(硫化ジメチルDMSは、大気中で酸化されて硫酸エアロゾルとなり雲の凝結核を形成する。海洋で植物プランクトンやバクテリアが増えると、海洋から大気へのDMSの放出量が増えて、雲の形成を促進する。)
鉄は地殻中の4.7%を占め、酸素・ケイ素・アルミニウムに次いで4番目に多い元素であり、金属元素の中ではアルミニウムに次いで2番目に多い。鉄は現代生活に欠かせない金属の代表でありながら、鉱石中では、そのほとんどが酸化されたかたちの化合物や結晶として存在している。そのため土壌には鉄が豊富に含まれていても、鉄分の供給方法としては、自然界では広葉樹の森から川を通じて海に運ばれる「フルボ酸鉄」や「フミン酸鉄」に依存するが、ダムや河川の改修工事などでこのシステムが機能しなくなり、生態系の鉄欠乏が常態化している。水溶性のフルボ酸やフミン酸は、鉄を錯体化しこれを「フミン酸鉄」や「フミン酸鉄」として鉄イオン化(Fe3+やFe2+)して海藻や植物プランクトンが吸収しやすい形に変換する。鉄イオン吸収や光合成、そして呼吸を促進し、食物連鎖の底部にある海藻や植物プランクトンの成育に大きく貢献して、磯焼け回復にも役立っている。
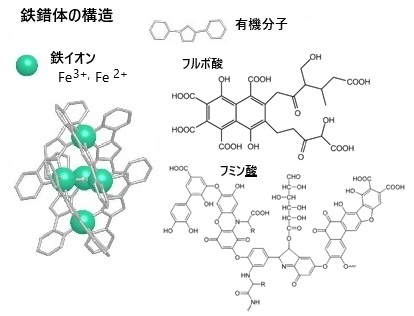 なお、フルボ酸やフミン酸は鉄だけではなく、様々ミネラルを錯体化する。.こうしたミネラルが海へと流れつくよう、河川の上流では植林活動を促進し、海の生態系を復活する取り組みも行われている。
なお、フルボ酸やフミン酸は鉄だけではなく、様々ミネラルを錯体化する。.こうしたミネラルが海へと流れつくよう、河川の上流では植林活動を促進し、海の生態系を復活する取り組みも行われている。藻類にとって、鉄は必須微量金属元素の一つ であり、植物プランクトンの光合成系や呼吸系 における電子伝達、クロロフィルの生合成、硝酸及び亜硝酸の還元などに深く関与しており、 藻類増殖に不可欠な元素である。葉に光があたると、はじき出された電子が次々に受け渡されていき、最終的にATP(エネ ルギー通貨)や NADPH(相手に電子を与える還元剤)が作られる。 この電子の受け渡しを行うのが鉄である。鉄が不足すると植物は光化学系の特に光化学系Iが作れない。またクロ ロフィルの生合成経路には鉄を必要とする酵素があるため、葉は黄色くなり、光合成能力 は大きく低下する。
特に、南極海や北太平洋亜寒帯域などの高栄養塩海域では、表層に多量 栄養塩 (硝酸塩・リン酸塩・ケイ酸塩)が高濃度で存在するにもかかわらず、鉄の不足が植物プランクトンの増殖を制限する要因となっている。鉄が不足すると、植物プランクトンの光合成が抑制され、海洋の一次生産能力が低下する。これにより、海洋生態系全体に影響が及び、食物連鎖の上位にある魚類や海洋哺乳類にも影響が出る。
キレートchelateの中でも、金属イオンを取り囲むようにして結合した構造を「錯体complex」と呼ぶ。フミン物質が光吸収や化学反応において重要な役割を果たし、水中での光化学反応や微生物の活動に影響を与える。 鉄は、ほぼ全ての生物にとって必要であるが、特に植物や植物性プランクトンといった一次生産者にとっては、必須栄養素である。鉄はイオン化(電子を帯びた状態)し、土壌や水に溶けた状態でないと吸収されない。しかも、酸素に触れるとすぐに酸化して結晶化し、沈殿する。 鉄は普遍的に存在してはいるのですが、ほとんどが結晶化していて、吸収しやすい形に変換させる機能が備わっている一部の植物を除き、簡単には吸収することが出来ない。鉄は大切でありながらも吸収しにくいミネラルの1つで、欠乏すると、植物は葉が黄色くなり、やがて枯れてしまう。 こうした問題を解決するのがフルボ酸鉄(orフミン酸鉄)である。森で作られるフルボ酸(orフミン酸)が土の中に含まれる鉄をキレート(掴む)すると錯体化し、フルボ酸鉄(orフミン酸鉄)となって地下水や河川に流れ出して沈殿することなく移動できる。やがてその水を利用する農作物、海中の植物プランクトンや海藻に鉄として吸収されることとなる。鉄単体では難しかった吸収が、フルボ酸やフミン酸の力によって可能になる。この働きは海の生物にとって大きな意味を持つ。 海底には鉄がたくさんあるにもかかわらず、水に溶けない状態で沈殿しているため、海水中の鉄不足は深刻化してる。このことは、海藻が著しく減少する「磯焼け」の原因にもなっている。
| 特徴 | 錯体 | キレート |
| 配位子の種類 | 金属イオンに複数の配位子(分子またはイオン)が結合した化学種を指す。 | 多座配位子が必須 |
| 環構造の有無 | 環構造は必須ではない。配位子が金属イオンの周囲に配位結合を形成。通常、配位子は1つの原子で結合している。 | 環状構造を形成 |
| 安定性 | 錯体には単純な配位結合も含まれ、特に環構造を必要としない。配位子や条件に依存する。 | 一般的に錯体よりも高い |
| どちらも化学において金属イオンと他の分子(配位子)が結合した構造。 「錯体」は広い概念で、「キレート」はその中でも特に多座配位子によって環状構造を形成する特別な場合を指す。 |
||
石灰藻はその名の通り、炭酸カルシウム成分を藻体に含む藻類の一種、北極や南極から熱帯、さらに潮間帯下部から水深200mにも及ぶ海底にまで生育し、現在までに世界で約830種が報告されている。 海水とわずかな光さえあれば、どこにでも生育できる海藻と言える。
このまま温暖化により高い水温で推移すれば、多くの藻食性動物の活性が高まり食害の進行が速まり、秋から春先にかけて、より多くの海藻が食べ尽くされる。)
海洋に鉄分を投入して植物プランクトンの増殖を促し、二酸化炭素CO2を吸収させるという温暖化対策が提唱されている。いくつかの研究結果から、海洋生物に悪影響を及ぼす酸素欠乏状態を引き起こしたり、一部の生物に有害なタイプのプランクトンが増殖する危険性があると言われたが、今回、このような問題点には根拠がないと示された。また、鉄分が投入された海域で増殖したプランクトンは、大部分が海底に沈み、堆積物に埋もれることが分かった。事実ならCO2の長期間にわたる封印を実現できる。 中規模であるが現場鉄散布実験を実施した結果、現在までの計12回の全ての実験において、現場鉄散布により植物プランクトンの二酸化炭素を消費する光合成能力の増加が確認されている。しかし、表層の従属栄養細菌や動物プランクトンの増殖も促進されることから、顕著な生物ポンプ能の増加による、地球温暖化の抑制効果はあまり期待できないことが実験結果により判明している。
また、鉄散布により海洋生態系を人為的に変化させてしまう可能性があるため、大規模な海洋鉄散布は行うべきではないという意見が多くの海洋学研究者か ら出された。このため、現在、地球温暖化抑制策としての現場鉄散布実験は、あま り現実的でないと考えられている。しかし、現場鉄散布実験を通して、これ ら外洋域の植物プランクトンの増殖が海水中の鉄の減少により著しく制限されていることを実証すされた。また、現場鉄散布実験で得られた結果は、海洋における炭素の動態を解明する上で、大変貴重なものとなっている。近年、地球温暖化による海洋生態系への影響も懸念されている。地球温暖化で海洋表層の水温が高くなることにより、海洋の成層化、つまり表層と中深層の水が混合しにくくなることが、より強化されることが予測されている。これにより、海洋中深層から表層への栄養物質の供給が弱まり、植物プランクトンの増殖力も弱化する可能性がある。植物プランクトンが減少すると、それを餌とする捕食者・動物プランクトンや、さらにそれを補食する魚類などにも影響を及ぼすことが予測されることから、ひいては海洋生態系全体を変化させる危険性が高まる。
しかしこの実験では、植物プランクトンの1種である珪藻diatomが繁茂し、粘着性が高い大規模なグループが形成された。しかも、発生したプランクトンの50%以上が海底に沈み込んでいた。
珪藻
珪藻は単細胞生物であるが、核膜を有する細胞核を持つ真核生物に分類され、原核生物の古細菌や真正細菌から分離される。珪藻は植物プランクトンの一種で、プランクトンの種類の中で最も一般的な種.である。しかも、周辺植物とのコミュニティにおいて不可欠な要素を構成している。珪藻は、海洋や淡水で主要な一次生産者として炭素循環に重要な役割を果たす。こうした変化が海洋炭素循環にも影響を与え、地球全体の気候システムに関連する可能性がある。例えば、藻類が増えることで炭素を多く吸収する一方で、過剰増殖により酸素不足や海洋の富栄養化のリスクが発生する。つまり、海氷藻類の放出は単なる「溶けて流れる」以上に複雑な相互作用を引き起こし、生態系や気候に波及する多層的な影響力を示す。
珪藻はミクロの生物であるが、その生物量をあなどれない。地球規模で年間360億tもの有機物を生産している。この量は陸上植物全体が作り出す量にほぼ匹敵する。さらに、珪藻が毎日生み出す酸素の量は、全地球上の光合成生物が放出する酸素の25%にも達する。珪藻は地球生態系の中で、大きな役割を演じている。
珪藻は微細藻類として分類されるが、化石の証拠は、珪藻が中生代ジュラ紀の約1億5000万年から2億年前、またはその前に発生したことを示唆している。珪藻類の最も古い化石の証拠は、ジュラ紀後期にタイから産出された琥珀の現存するHemiaulus属の標本である。生物進化の歴史の中では比較的新しい生物と言えるが、その生存戦略として、さまざまな最新メカを細胞に搭載した生物でもある。
珪藻細胞は他の単細胞性の藻類と異なり、球形や楕円球の形をとらずに、複数の面からなる、カドのある形をとっている。これにより、体積あたりの表面積を大きくする。しかも、珪藻はガラスのような硬い被殻をもっているため、その多様な形を保つことができる。実際、珪藻には細長い種類や平べったい種類がかなり多い。特に、付着珪藻ではこの傾向がより高い。体積に対する表面積の比が大きくなると、単位原形質あたりに供給する無機塩類などの栄養と、CO2値は高くなるので、その珪藻の複雑な形は生存戦略上とても有利に働く。加えて、珪藻はどの種類も大きな液胞を持つため、核のある中央部を除き、原形質は被殻の内側にへばりついたように配置している。つまり、被殻の内部は原形質で一杯に満たされてはいない。これにより、細胞における原形質の単位体積あたりに供給される栄養とガスの量を増加させる。これが、生存戦略上有利に働く。
植物プランクトンである珪藻のガラスのような殻が、薄暗い環境での光合成にどのように機能しているのかが明らかになった。珪藻は、外側の被殻という透明で硬い多孔質の殻で被われており、穴の大きさと間隔、その配置によって、光への応答が異なっていた。被殻と相互作用する波長が光合成で吸収する波長と一致することも明らかになった。それが、細胞全体で吸収できるように、被殻の様々な部分が光を再分配していたことも解明された。さらに、高照度から低照度への移行期には、光合成を助けるために十分な時間、被殻内を光が循環し、光合成を約9.8%促進することも発見された。
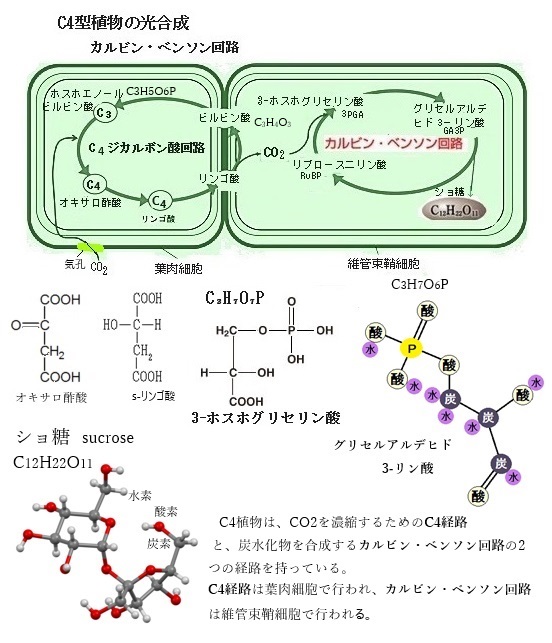 独立栄養生物の基本的な代謝としては光合成がまずあげられるが、それは光や温度、二酸化炭素や酸素などの溶存ガス分子、窒素や硫黄などの主無機栄養、および鉄や亜鉛などの微量栄養など様々な環境要因に晒されている。
独立栄養生物の基本的な代謝としては光合成がまずあげられるが、それは光や温度、二酸化炭素や酸素などの溶存ガス分子、窒素や硫黄などの主無機栄養、および鉄や亜鉛などの微量栄養など様々な環境要因に晒されている。水中ではCO2の溶存量は非常に低く、また溶存分子の拡散速度もかなり遅いことから、海洋の光合成生物はCO2獲得のための積極的なシステムを必要とする。
特に海洋のような高塩・高アルカリ環境下では、さらにCO2獲得が困難となるため、このような環境においても海洋性珪藻類は、CO2に対して高親和性光合成を行う高い一次生産力を保有している。その一つが、海洋性珪藻類のCO2濃縮機構 CO2-concentrating mechanism(CCM) である。そのCCMは恐らくほとんどの藻類が進化した後に、大気組成の変化を受けて獲得されたもので、その起源は3〜4億年前に遡ると見られている。CCMの獲得は、生態系の中での広範な収斂進化の賜物であり、従って、これにかかわる分子はかなり多様であると見られている。
CCMは本質的に大きく2つのステップから成ると考えられる。1つめは、細胞膜上に存在する無機炭素輸送体によって細胞外から能動的に無機炭素を取り込むステップで、2つめは、取り込んだ無機炭素の漏れ出しを防ぎながらCO2へと変換し、効率良く炭酸固定化酵素ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO)へと供給するステップである。珪藻類は2億5千万年前に現れ始めた最も新しいCO2濃縮機構生物と考えられ、その多様な細胞や葉緑体のサイズおよび形態から、ユニークなCO2濃縮機構を有しているものと考えられている。
海洋性珪藻は、地球全体の炭素固定量の約 20%を担う重要な一次生産者であるが、現在の大気圧環境下における溶存CO2濃度は珪藻にとって不十分である。そこで海洋性珪藻は、無機炭素不足を解消する ために、溶存無機炭素dissolved inorganic carbon (DIC) を能動的に細胞内に取り込み、RubisCO周辺のCO2 濃度を高める無機炭素濃縮機構 (CCM) を発達させた。このCCMの働きによ り、珪藻は無機炭素に対して高い親和性を示す。珪藻では、無機炭素輸送体と炭酸脱水酵素carbonic anhydrase (CA) を用いて無機炭素を濃縮する生物炭素濃縮メカニズム biological carbon fixation が機能している。無機炭素輸送体は、溶存無機炭素を 細胞内に取り込みを支える重要な因子である。
海洋性タラシオシラ・シュードナナPhaeodactylum tricornutum では、細胞膜及び 葉緑体4重膜上の無機炭素輸送体が 溶存無機炭素を取り込み、葉緑体内部の炭酸脱水酵素 (CA) によって重炭酸イオンHCO3- が CO2 に変換 され、 炭酸固定化酵素RubisCO に供給されると考えられている。
一方、海洋性中心目珪藻 Thalassiosira pseudonana (T. pseudonana は、珪藻生理学研究のモデル)では、 細胞外に局在する炭酸脱水酵素 (CA) がHCO3-を膜透過性を持つ CO2 に変換し、細胞膜を透過させる ことで無機炭素の取り込みを行う。取り込まれた CO2 は、細胞内の炭酸脱水酵素 (CA) によって HCO3- に変換され、 P. tricornutum と同様に葉緑体 4 重膜上の無機炭素輸送体が 溶存無機炭素を取り込み、葉緑体内部の 炭酸脱水酵素 (CA) によって HCO3- が CO2 に変換され、 炭酸固定化酵素RubisCOに供給されると考えられている。 T. pseudonana の無機炭素濃縮機構 (CCM) は、細胞 内外の炭酸脱水酵素 (CA) と葉緑体 4 重膜上の無機炭素輸送体によって支えられている。現時点で、T. pseudonana の 炭酸脱水酵素 (CA) の局在は明らかになって来ているが、無機炭素輸送体についてはC4光合成のような維管束鞘細胞と葉肉細胞の複雑な相互作用による有機酸の脱炭酸反応を介する濃縮経路とは異なり、無機炭素濃縮機構 (CCM) は細胞膜や葉緑体膜に局在する無機炭素輸送体を介した無機炭素の直接的な輸送によると考えられている。
(溶存無機炭素は溶液中の無機炭素の総称で、全無機炭素total carbonとも呼ばれる。その無機炭素には二酸化炭素 CO2、炭酸H2CO3 、炭酸イオンCO32−や重炭酸イオン HCO3- が含まれる。)
珪藻は二次共生によって成立した生物と見られている。一次細胞内共生はシアノバクテリア種の祖先生物が真核従属栄養細胞(生育に必要な有機炭素源を自分自身では生産できず、主に植物や動物に由来する有機化合物を摂取することで獲得する真核生物群)に細胞内共生によって取り込まれることによって、植物や緑藻、紅藻の起源となる始原植物細胞を保有した。細胞共生とはある生物の細胞が他の生物の細胞内に入り込んで(取り込まれて)一体となって生活することで、この現象が生物進化、特に真核光合成生物の進化と多様性の実現に大きな役割を果たしてきた。藻類にみられる多様性の本質は細胞共生、特に真核生物同士の共生(真核共生)にあるが、その事前段階として従属栄養の真核生物が、原核生物のシアノバクテリアを食作用によって取り込む一次共生によって誕生した、光合成真核生物の「一次植物」と呼ばれる進化過程が存在していた。
淡水域・汽水域・海水域に広く分布する珪藻は、重要な一次生産生物であり、更に紅藻を細胞内へ取り込んで進化した二次共生藻でもあると見られている。珪藻や紅藻などの水域に存在する光合成生物は、陸上植物と異なる進化を遂げており、それぞれの生存環境に応じて異なる集光性色素タンパク質を持つ。
水中を透過する太陽光エネルギーは、青色から緑色の光であるため、陸上植物よりも限られた範囲の光エネルギー資源を確保するために多様性が生まれた。生息域が広く分布する珪藻は、重要な一次生産生物であるが、紅藻を細胞内へ取り込んで進化した二次共生藻でもある。褐色を呈するその原因は、褐藻から単離された集光性色素タンパク質であるフコキサンチン-クロロフィルa/c結合タンパク質ucoxanthin-Chlorophyll-Protein Complexes (FCP) にある。FCPは太陽光エネルギーの中の青色から緑色の光を吸収する。これは植物が持つ集光性色素タンパク質の吸収領域である赤色と青紫色と大きく異なる。珪藻類は、こうして紅藻がさらに真核従属栄養細胞に共生することによって成立した。
二次共生の過程を経て成立した珪藻の葉緑体は、その構造も複雑化して4重膜構造を有している。外側2膜は葉緑体小胞体、内側2膜は葉緑体包膜と呼ばれる。外側の膜は核膜と融合しており、核葉緑体連合を形成している。さらに葉緑体内には、植物の光合成CO2固定反応を担う酵素、ピレノイドというRubisCOを多量に含むタンパク質顆粒が存在している。
また珪藻ゲノムの詳細な解析から、珪藻葉緑体で働くタンパク質のうち、核コードのものの75%以上が緑藻型である一方、葉緑体コードのものは紅藻型であることが指摘されている。つまり、珪藻の一次共生では緑藻の取り込みと葉緑体化に伴う遺伝情報の核移行の後、間もなく紅藻が取り込まれて緑藻型葉緑体に取って代わったことが示されている。この複雑な過程を経て成立した珪藻類の核ゲノムの炭素代謝系は、様々な系統由来の代謝系の細胞小器官organelle間で重複した経路を経て、これらが基質やタンパク質の輸送経路を通じて互いに連携し、既知の真核型の代謝には見られない精緻な代謝ネットワークを完成させている可能性が高い。
直接的な光合成基質となるCO2が極度に欠乏している海水中でも、海洋性珪藻類が、高い光合成活性を維持していることからも、CO2だけでなくHCO3-も取り込んでいることが示唆されてきたが、実際に、これまでの生理学的解析では、海洋性珪藻類は、CO2とHCO3-の両方を取り込む能力を有していることが明らかにされている。
珪藻のタラシオシラ・ワイスフロッギー Thalassiosira weissflogii がC4光合成を行っている証拠が報告された。C4光合成は陸上植物ではトウモロコシなどでフォスフォエノールピルビン酸カルボキシラーゼ (PEPカルボキシラーゼ)経由で、効率良く炭酸固定化酵素ribulose-1,5-bisphosphate carboxylase/oxygenase(RubisCO)が炭素を受け取り暗反応が進む光合成で、RubisCOだけで炭酸固定をおこなうC3光合成よりも効率の良いことが知られている。珪藻のRubisCOのCO2半飽和定数は 30~60㎛であるが、海水にとけ込んでいるCO2は10㎛程度であれば.、珪藻は常にCO2欠乏状体になっている。元来もっているRubisCOの能力を目一杯使えていない可能性があった。ところが中心珪藻のThalassiosila weissflogiiのPEPカルボキシラーゼの生化学的な活性特性と細胞の増殖試験から、この種がC4光合成を行っていることが分かった。
珪藻のサイズは一般に20〜200㎛(1㎛ = 0.001mm)で、いくつかは〜2,000㎛の範囲で大きな種もいる。珪藻は単細胞生物であり、孤立した細胞として、またはリボン・扇状・ジグザグ、または星の形をしたコロニーで発生する。しかも、珪藻はどこにでもいる生物で、海・川・湖沼・水槽・水田といった水中はもとより、水がうっすら滲み出す湿った岩の上、コケの表面、湿り気のある畑の土の表面などにも珪藻は生息している。水質的にも高山の雪解け水の流れから、異臭を放つドブ川のような非常に汚れた水にまで生存している。例えばpH1.2の群馬県草津温泉の「湯畑」からも出現する珪藻もいる。
草津温泉「湯畑」の微生物といえば、湯畑などで観察出来る紅藻イデユコゴメ(Cyanidium属) が有名であり、紅藻ガルディエリア(Galdieria属)や珪藻ハネケイソウの一種(Pinnularia属)などの真核藻類 も生育している。原核生物では好酸性好熱性細菌(Bacillus属)や好酸性好熱性の古細菌archaeaも確認されていた
アフリカのケニアにあるpH11と強アルカリ性を示すナクル湖にも珪藻はいる。数千から多いときで数百万のフラミンゴが飛来する。しかし、近年の異常増水によりフラミンゴの食糧である藍藻類の生育量が激減し、「湖の浅い部分はフラミンゴが多く水面が見えないところも多い」などと記述されていたが、今では、失われた過去の偉観となる。
アメリカのユタ州の北部にある、西半球最大の塩水湖グレートソルトレイクGreat Salt Lakeの塩分濃度は、海水の約3~5倍もあるが、そんな湖水にも珪藻はいる。
また、海産ミジンコのに付着して生きているような種類や、クジラの体表面に付着する種類も知られている。
海藻はすべて付着生活をしている。このため海藻は沿岸の浅い海域でしか生息できない。珪藻では付着種のほか浮遊性種もいる。大海のど真ん中や、水深数千mある海域でも浮遊珪藻は生きている。珪藻が様々な所で生存できるのは、珪藻の種類が多く、その生態的特性も様々な環境に適応して多様に分化しているからである。
海氷藻類 sea ice alga
海氷の微生物は非常に多種で、豊富な藻類・バクテリア・原生動物が生息している。特に藻類はシンパジック環境sympagic environmentを支配しており、北極圏の海氷と関連していることが判明した単細胞真核生物は1,000以上と推定されている。sympagicとは「共存」を意味し、この環境では、海氷内やその表面、または下部に多くの微生物や藻類が生息している。種の構成と多様性は、場所や氷の種類、および放射照度によって異なる。通常では、北極圏や南極圏などのペネート珪藻pennate diatomsが豊富で、氷の底に付着した長さ1mまでのフィラメントを形成する。
メロシラ科に属する珪藻・タルケイソウMelosira arctica(arctica=北極.)は、北極圏にも広く分布しており、海洋生物の重要な食料源になっている。タルケイソウMelosiraはバケツのような形の殻を持つ珪藻で、長い群体(糸状体)を作る。湖沼や河川の下流などで見つかる。細胞の中央部に被殻のつなぎ目が見える。糸状体が他の藻類に付着していることが多い。
海氷の柱全体に海氷藻類群集が見られるが、その存在量と群集の構成は時期によって異なる。海氷上および海氷内で藻類が利用できる多くの微小生息地があり、藻類のグループごとに異なる好みがある。例えば、冬の終わりから春先にかけて、N. frigididのような運動性の珪藻が、塩水チャネルが到達する限り、氷の最上層を支配していることが分かっている。その存在量は、初年度の氷よりも多年氷の方が多い。さらに、渦鞭毛藻類(うずべんもうそう)は、南極の海氷の南半球の春初期に優勢であることも知られている。
渦鞭毛藻類は2本の鞭毛を持つ単細胞藻類の一群である。細胞の表面に縦横の溝を持つ、独特の形をしている。渦鞭毛藻の細胞はテカtheca(包膜)と呼ばれる独特の細胞外被構造を持っている。植物は通常、細胞膜の外に細胞壁などの外被構造があり、細胞を包んでいる。一方、一部の藻類では細胞の形態を保持する外被構造が細胞膜の内側に発達する場合がある。渦鞭毛藻のテカthecaがその例で、細胞膜の内側に小胞があり、その中にはセルロースからなる板状の構造があり、それは「よろい」に似ていることから鎧板とも呼ばれる。このプレートの形と数、発達の程度および配列は種によって一定であるため.、この渦鞭毛藻の独特な外観が分類上の重要な特性と認識されている。
渦鞭毛藻の多くの種では、テカは上殻(じょうかくepitheca)と下殻(かかくhypotheca)とよばれる二つの部分からなる。上殻と下殻の間に横溝があり、また横溝と交差して細胞後方に縦溝が伸びている。典型的な渦鞭毛藻では上殻と下殻の大きさはほぼ同じであるが、種によって上殻が極度に退化した、または未発達なものや、下殻が退化した、または未発達なものなどがある。
形態は非常に多岐にわたるが、縦横2本の鞭毛を持つことが最大の特徴である。一本は鞭状のものを後ろにひき、もう一本は羽状のものを横向きの溝に沿って巻き付けるようにしている。推進力の異なる2系統の鞭毛を備えることによりに、細胞は急停止や急発進、方向転換など、多彩な遊泳が可能となっている。また、細胞の表面に鎧板と呼ばれる硬い板状の構造が並び、時には突起を有する角張った形態を持つ。ただし、無殻のものもある。
渦鞭毛藻の約半分は葉緑体を持ち、光合成をおこなう。残りの半分は従属栄養性で、他の原生動物を捕食して生活する。また光合成を行う渦鞭毛藻の中にも活発な捕食行動を行うものもいる。光合成能を持つものも、多くは溶存の、あるいは粒子状の有機物を取り込むことができる。このような栄養摂取を「混合栄養」と言い、他の藻類にも例は多いが、この群で圧倒的に数が多い。中には寄生性のものもある。
光合成能を持つ渦鞭毛藻は一次生産者として食物連鎖中の重要な位置を占めるが、中でも渦鞭毛藻類の仲間の褐虫藻は海洋動物や原生動物の細胞内共生体であり、特にサンゴ礁の生成に極めて重要な役割を担っている。
多くの造礁サンゴは褐虫藻を細胞内共生させている。褐虫藻は宿主内で光合成を行い、光合成代謝産物を宿主に提供している。
近年、海水温の上昇により褐虫藻の光合成装置、特に酸素発生部位である光化学系II(水H2Oを分解し酸素を放出し、その後の光合成に繋がるための電子を提供する)の光合成活性が低下する光阻害が見られている。高温による褐虫藻の光阻害に伴いサンゴの白化が起こる。他の光合成生物でも同様に高温ストレスにより光阻害が助長されるが、それらの温度感受性は褐虫藻ほど高くはない。
光化学系IIの反応中心から電子が放出されると、電子を失った反応中心に電子が供給される。この電子は水を分解することによって得られる。
水2分子(2H2O)を分解すると、二つの電子(e-)、四つのプロトン(H+)、一つの酸素分子(O2)が発生する。光合成で発生する酸素はこれに由来する。
(藻類は明確な分類群ではなく、緑藻・紅藻・褐藻など複数の系統群を含む。種類によって形態は様々で、細胞壁の成分や色素が異なる。
珪藻はストラメノパイルStramenopilesと呼ばれる分類群に属し、シリカ【二酸化ケイ素SiO2】でできた複雑な幾何学模様の殻を持つのが特徴。)
珪藻の細胞膜の外側を覆う構造は幾つかのものがあり、なんといっても目立つものが、珪藻の細胞壁に当たる部分の被殻frustuleで、そのユニークな特徴は、錐台(すいだい=Frustum)と呼ばれる水和二酸化ケイ素(SiO2・nH2O)でできた細胞壁に囲まれていることにある。これらの錐台は構造的な色を生み、「海の宝石」や「生きたオパール」と呼ばれる。
水和二酸化ケイ素の化学式はSiO2・nH2Oで、n は水分子の数を示し、シリカゲルの水和状態に応じて変化する。構造体の所々にケイ酸【[SiOx(OH)4−2x]n 】)が残っている。 脱水によって多孔質になっていることに加え、ケイ酸が持つ-OHと水分子が水素結合を形成する ため、多量の水分が吸収される。
これは無色透明の珪酸質(SiO2・H2O)でできている。被殻は幾つもの珪酸質構造体の要素が組合わさってできているもので、それぞれに形態学上の呼び名がある。そして、その外側には薄い粘液質の層がある。これは有機被覆 organic casingと報告されている。また被殻を裏打ちするディアトテプムdiatotepum と呼ばれる有機質の膜状の層の存在も透過型電顕の観察により一部の種で知られている。珪藻は、その小殻の構造に応じて 2 つの大きなグループに分けられる。中央珪藻は、放射状の対称性を持ち、海洋環境では優勢である。羽状珪藻は、左右対称性を持ち、主に淡水に生息している。
珪藻の増殖速度は種によって様々で、大型の付着性種では遅いものでは2週間で1回しか分裂しないようなものもあるが、浮遊性種は一般的に増殖速度が速いものが多い。これは珪藻が鞭毛などの運動器官を持たず、静置していればすべて光合成のできない暗い水深へ沈んでしまう運命を持っていることに関係がある。実際は水の中の流れのために、上方へ巻上がってくるものもあり、全部が全部下方へと沈んでいくわけではない。増殖速度を高めれば、暗い水中へと沈む細胞数以上の細胞を新しく生成.できる。つまり、沈む以上に細胞数が増えれば、種の生存戦略上支障はない。1日に2回分裂をする種もあるが、培養では1日に3回分裂した例もある。
核分裂後、細胞質が分裂し、その後分裂面に向かい合って新しい娘殻が形成される。これらの娘殻は、親被殻の「蓋」と「身」に関しては、どちらも新しい「娘殻」が常に親の被殻の内部で形成されるため、親の被殻よりサイズの大きい「蓋」を作ることが不可能なためである。このため、細胞分裂で作られる新しい殻は、常に「いれこ」細工のようになり、細胞分裂を行うたびに、そのサイズはどんどん小さくなっていく。イチモンジケイソウEunotia formicaでは、.分裂するたびに被殻は長さが1μmずつ減少する。こうして無性的な2分裂を何回と無く繰り返すと、ついには被殻の長さが最初のものと比べ1/2~1/4にまで減少する。
分裂によりサイズが元の大きさの1/2~1/4まで減少した珪藻は、減数分裂を行い配偶子を形成する。そして、その後有性生殖をする事によって再び大きさを回復する。この過程では接合子が2倍~4倍くらい大きくなるが、この増大する接合子が増大胞子 auxosporeと呼ばれる。接合後、増大を始めたものを便宜上「若い増大胞子 young auxospore」と呼び、増大が終わった完成品のみを「増大胞子auxospore」と呼ぶ場合もある。
中心類珪藻と羽状類珪藻とでは有性生殖が異なる。前者は卵と精子による卵生殖であるが、後者では鞭毛を持った精子は形成されず、珪酸被殻を持たない裸の同型配偶子がアメーバ的に移動し接合する。
(減数分裂
① 遺伝子は、細胞内の核の中にある染色体に含まれている。
② ある個体の体細胞の染色体は、2組からなり、それぞれ母親から受け継いだ1組と、父親から受け継いだ1組。これらは、体細胞分裂を通しても同じものが複製される。
③ 卵や精子といった配偶子が生成される際の分裂は減数分裂で、配偶子に含まれる染色体は1組のみ。体細胞の染色体と比べて半減していることになる。
1組ずつ染色体をもった配偶子が2つ集まる受精により、体細胞と同じ量の染色体を得ることになる。受精してできた受精卵は、発生を経て、親と姿かたちの同じ成体になる。)
珪藻は、水生環境で春と秋に毎年大量に咲く原生生物であり、世界の海洋における光合成の約半分に関与していると推定されている。
十分な栄養素と日光の存在下では、生きている珪藻の集合体は、無性多重分裂によって約24時間ごとに倍増する。個々の細胞の最大寿命は約6日である。
目次へ
グリーンランドは厚い氷河におおわれた島である。その氷河の周りを縁取るように岩盤が露出している。1960年代~ 70年代のグリーンランドにおける鉱物資源探査の過程で、イスア地域には世界最古の地層が分布していることが知られた。イスアには、グリーンランド語で「地が消え果てる場所」という意味があると言う。そこから先は大地が大陸氷河に覆われ見えなくなる地域のことを指す。今では、大地の果ての向こうに初期地球の風景が見えてくる。
 |
| タービダイトは、大陸棚の端の急な斜面を滑り落ちる水中雪崩によって、大陸棚の下の深海トラフや海盆に堆積する。 物質が海の谷に眠るとき、最初に沈殿するのは砂やその他の粗い物質であり、次に泥が沈殿し、最終的には非常に細かい粒子状物質が沈殿する。 タービダイトは海底での土石流である乱泥流による堆積物であり、未固結の堆積物が水中で攪拌され、流れ下る現象により積層された。 |
イスア地方の礫岩堆積層の存在は、38億年前に海であったことを示している。その後の調査により、堆積岩と思われた地層は広域的な 変形作用で生じた片理の発達した火山岩であり、イスア地域での堆積岩の占める比率は極めて低いことが解かった。それは.礫岩堆積層を走向方向に追跡すると「礫」が完全に消え、珪長質の火成岩に変わってしまっている様子からも観察されている。
現在の地球には、約14億km3もの海水が存在する。加えて地中には、この2倍くらいの水が存在すると言われている。その多くはH2Oではなく、水酸化物イオンOH-として鉱物中に含まれている。その大量の水は、海洋プレートが「沈み込み帯」から、地球深部へ持ち込まれた。その海水がマグマによって熱せられて、活発な熱水循環が起こる。その後の過程で、プレート内部にOH-を含む粘土鉱物などが大量に作られる。やがてマグマオーシャンの中に含まれていたOH-が、冷え固まるときに、鉱物中に取り込まれる。
地球型惑星の起源となる原材料物質は、岩石を主成分とする石質隕石の中でも、特にケイ酸塩の球粒組織を多く含み、しかも溶融しないまま岩石質と金属質が分かれないまま、水や揮発性元素を多く含むことのが特徴である。 1969年にオーストラリアのビクトリア州マーチソン村に落下した「マーチソン隕石」は、地球上には存在しないアミノ酸を含むことで知られる炭素質隕石であったが、15%もの水が含まれていた。原始地球型惑星を作った微惑星に、地球の起源を求めるのが自然な成り行きとなった。
原始地球の大気の気圧が現在よりもずっと高かった水が、気体ではなく液体として地球に存在するには、地表付近の温度が数百度以下に下がらなければならない。
 |
| 花崗岩の中のジリコン結晶 |
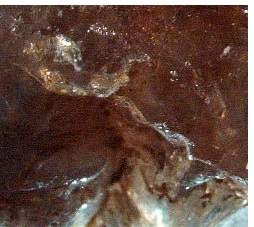 |
| ジルコンzirconとは、化学組成 ZrSiO4 ケイ酸塩鉱物、結晶系は正方晶系を取る。 岩石を構成する鉱物の中には、化学反応を起こしにくく安定であるために、信頼度の高い年代を得ることができるものがある。その代表格がジルコンである。 ジルコンには放射性元素であるウランやトリウムが多く含まれるために、精度の高い年代測定が可能。 現在最も古いとされるジルコンは、44億400万年前のもの、このジルコンの化学組成を解析すると、この鉱物はもともとは花崗岩に含まれていた。花崗岩は、プレートの沈み込みによって生じたマグマがゆっくり冷え固まった岩石である。 |
イスアに分布する縞状鉄鉱床の露頭です。縞状鉄鉱床とは,磁鉄鉱と石英粒子が繰り返した地層です。縞状鉄鉱床は微生物の作用で形成されたのではないかという説があり、微生物様の化石の探査も行われました。日本の研究グループは、縞状鉄鉱床を連続的にサンプリングして詳しい分析を行いました。詳しい分析の結果、縞状鉄鉱床には、厚さ数mmのリン酸カルシウムに富んだ層が2枚みつかりました。リンは生命に必須の元素なので,リンの濃集は生命の存在を期待させます。。有機物の分析も試みられました。米国の研究者たちは、イスア地域のリン酸カルシウムに含まれる炭素の分析を行って、生命活動の痕跡が認められると発表しています。縞状鉄鉱床には,厚さ数mmのクロムに富んだ層が発見されました。“クロムは小惑星の衝突でもたらされたのではないか” という説があり、小惑星に多いイリジウムの分析や、クロムの同位体比の測定が試みられました。クロムに富んだ層は、はたして、天体衝突事件があったことを物語っているのでしょうか?
イスア地域の岩石は38億年前の海底に噴出した溶岩と堆積岩からなる。しかしながら37億年前にはトーナル岩tonalite(無色鉱物である曹長石や石英が90%程度を占め、これに10%程度の有色鉱物を伴う深成岩)の貫入があったが、現在では変成されアミツオク片麻岩と呼ばれているので、周辺の岩石は熱変成を被っている可能性がある。イスア地域にかけて分布しているアミツオク片麻岩は、カナダのアカスタ片麻岩が発見されるまでは、世界最古の岩石として知られていた。その特徴的な縞模様の片麻状構造の発達は顕著である。
38 億年前の溶岩や泥は、その後のプレートテクトニクスによって押し潰されたり、500℃を超える高温にさらされたりして別の岩石に変成しているのがほとんどである。広大なイスア地域であるが、38億年前の海底の泥の痕跡をとどめる岩石を探すのは、困難を極める。更にその後の広域変成作用でほとんどの岩石は角閃岩相【斜長石(白色)の点紋が多い角閃石(黒色〜濃緑色)を主体、の変成岩】からグラ ニュライト相【最も高い温度,圧力でつくられる変成岩】の高変成を受けて、初生的組織を失っている。イスア地域では、グラ ニュライト相の高変成により結晶片岩化した溶岩が最も普遍的に見られ、それをガーベンシーファーという特殊な名前で呼ばれている。それでも、結晶片岩化を免れ、38億年前の岩石の痕跡を残すものもあり、その顕著な例として、露出する玄武岩質の枕状溶岩の組織を残した露頭が複数箇所で認められている。溶岩が水中で噴火し、枕のような形で積み重なってできるもので、 38億年前の海底火山活動を示す証拠である。岩質は変成作用により角閃岩質にかわり、形状は地殻変動により変形している。
地球最古の鉱物であるジルコンは、44億400万年前には地球表面の一部に海が存在した可能性を示している。 しかし、広くて立派な海の証拠が残っているのは、グリーンランドのイスアに分布する38億年前の地層であった。この地層は、粒子の大きい砂と細粒の泥が交互に繰り返す。つまり砂粒子は大きいので沈降しやすく、そのため浅い海に溜まる。一方泥は、深い海に堆積する。ところがイスアでは.、これらの堆積した深さが違う砂と泥が繰り返している。 それは、海溝の陸側の斜面だ。海溝の「沈み込み帯」では巨大地震が頻繁に起こる。この地震によって、比較的浅い斜面に溜まっていた砂層が崩れて、斜面を流れ下る「海底地滑り」が発生する。地滑りした砂は、深い場所の、普段は泥が溜まっている所まで流れ着いて海底扇状地を形成する。このようにしてできた砂と泥が繰り返す地層は「タービダイト」と呼ばれる。つまりこの地層は、海溝のように十分に深い部分を持つ、広い海が存在したことを示す。
しかも、イスアの地層群は、海洋プレートが沈み込む際に、その一部が陸のプレート側へ潜り寄せて形成される「付加体」の特徴が明らかになっている。
目次へ
有機物分解菌と原生動物は硝化菌と並ぶ重要な「ろ過バクテリア」で、「生物ろ過ができ上がる過程」は有機物分解菌が増殖するところからスタートする。そして「硝化菌」が増殖し最終的に原生動物が増える。
バクテリアと呼ばれるもののほとんどはこの有機物分解菌である。
原核生物(prokaryoteは、核や他の膜結合細胞小器官を持たない単細胞生物である。原核生物という用語は、古代ギリシア語の πρό (pró)「前」と κάρυον (káruon)「核」に由来する。原核生物は 細菌Bacteriaと古細菌Archaeaの2つのドメインに分けられる。細胞核を持つ生物は、第3のドメインである真核生物Eukaryotaに分類される。最初の「原核生物」は約8億年前に誕生したと考えられている。
原核生物は真核生物よりも先に進化し、真核細胞を特徴づける核、ミトコンドリア、その他の明確な細胞小器官のほとんどを持たない。かつては、細胞外膜を除いて、原核生物の細胞質内に細胞構成要素は存在しないと考えられていたが、タンパク質の殻、例えばエンカプスリン・タンパク質ケージに囲まれた準細胞小器官と考えられる細菌微小区画が、他の原核生物の細胞小器官とともに発見された。シアノバクテリアのような一部の原核生物は、単細胞である一方で、バイオフィルムによってまとまったコロニーを形成することがあり、大きなコロニーでは多層の微生物マットを形成することがある。粘液細菌のように、生活環に多細胞の段階を含むものもある。原核生物は無性生殖を行い、配偶子の融合を伴わない二分裂によって生殖するが、遺伝子の水平伝播は起こりうる。
その後、大きく遅れて約3億年前に誕生した「真核生物」は、核膜で囲まれた核と細胞小器官を持つ細胞からなり、動物・植物・菌類・原生生物(顕微鏡でしか見ることのできないアメーバ・ゾウリムシ・ミドリムシ・タイヨウチュウの仲間)などがこれに属する。真核生物のゲノムDNAはむき出しではなく染色体の中に収納され、細胞分裂によって新たにできた「娘細胞」(一つの親細胞から作られたまったく同じ二つの細胞)に、そして、有性生殖によって配偶子を介して次の世代に受け継がれていく。
自然界において、動物の排出物や、植物の枯れ枝・枯葉、そして、それらの死骸は、分解者とよばれる生物たちの働きで分解される。このとき、生物のからだに存在していた窒素もまた「土にかえる」。
生物生産によるNとPの取り込み比率(N/P = 16)が、不思議なことに、海水中の栄養塩濃度の比率(NO3- 【硝酸イオン】/ PO43- 【リン酸イオン】= 13~16)に近い。海水のN/P 比が16だから、生命進化の過程で、生物の元素組成比が海水に近づいたのか? それとも、海水中の栄養塩成分が、生物に取り込まれて分解再生を繰り返すうちに、余剰分が海洋から取り除かれて、海水の元素組成比が生物に近づいたのか? この謎は、誰にも解けない。
海水中の栄養塩の窒素とリンの平均比率(N/P)は16であり、生物体に含まれる平均比率も16である。海洋植物による栄養塩の取り込み比率は16に近いですが、生物体に取り込まれてからのNとPは、同じルートで全て海水に戻るわけではない。生物に取り込まれるルートも若干異なる。
① 窒素固定:大気由来の窒素分子N2を取り込んで体内でアンモニアNH3に同化する植物プランクトン(窒素固定生物)がいる。この作用により、海洋へN2が供給される。
湖沼に溶存している窒素は、窒素ガスN2を除いて、大きく無機態(DIN)と有機態(DON)のものに分けることができる。無機態窒素は、アンモニウムイオンNH4+、亜硝酸イオンN02-、硝酸イオンN03-の形態で存在している。硝酸イオンは、雨などの降下物の中に多く含まれ、雨が降った後、湖水中の硝酸イオン濃度が高くなることがある。これらの無機態窒素は栄養分として植物プランクトンに取り込まれ、体細胞に同化する。一方、有機態窒素は湖沼のプランクトン、その他の生物の遺遺骸、湖外から流入した落ち葉などに含まれる有機物がバクテリアや動物の捕食をとおして分解されて、アミノ酸や尿素として湖沼中に存在している。有機態窒素はさらに分解されるとアンモニアになる。多くの湖沼では、しばしば夏季の底層において底泥中の有機物の分解にともなうアンモニウムイオンの上昇が認められる。この他にも、生きている生物から、直接アンモニアや尿素が排出される。そして、これらは再び植物プランクトンの窒素源として取り込まれる。
植物プランクトンの中でも、窒素固定型シアノバクテリア(藍藻)の一種のアナベナAnabenaは栄養塩の窒素化合物が不足すると、水中に溶けている分子状の窒素ガスを直接利用し、窒素固定を行う。アナベナには空気中の窒素を同化する窒素固定能もあり、アカウキクサなど一部の植物と相利共生している。またアナベナは神経毒を生成する藍藻の一種で、野生動物をはじめ家畜やペットなどに害を与えることもある。夏期には大量発生してアオコを形成することもある。
シアノバクテリアcyanobacteria(藍色細菌)は、酸素発生を伴う光合成(酸素発生型光合成)を行う原核生物であり真正細菌の一群である。非公式の一般名である藍藻類の基礎を形成していが、それは科学的な分類ではない。シアノバクテリアは、世界的に広く分布する唯一の酸素性光合成原核生物であり、裸岩や砂漠の一時的に湿らせた岩、さらには南極の岩など、淡水・海・湿った土壌など多様な陸域および水域の環境に分布している。多様で極端な生息地で繁栄している。そのため地球規模の生物地球化学的サイクルの主要な原因となっている。彼らは地球上で最も古い生物の1つであり、化石記録は少なくとも21億年前に遡る。歴史的には「植物」に分類されていたが、原核生物である点で他の藻類や陸上植物(どちらも真核生物)とは系統的に大きく異なる。しかし、陸上植物のものも含めて全ての葉緑体は細胞内共生において取り込まれたシアノバクテリアに由来すると考えられており、植物の起源を考える上で重要な存在である。
シアノバクテリアは、様々な形態のクロロフィル・カロテノイド・フィコビリンなどの光合成色素を使用して、太陽光の光エネルギーを化学エネルギーに変換する。クロロフィルは、青い光と赤い光をよく吸収するため、残った緑色の光が散乱・反射されて植物の葉は緑色に見えることになる。カロテノイドは、葉緑体が吸収できない緑色の波長を含めて400nm~550nmにおける紫~青~緑色の光を吸収し、赤色や黄色は吸収されずに反射するため、人の目にはその色が見える。フィコビリンは、クロロフィルの吸収が少ない緑色の領域の光を吸収することができ、そのため見た目は青から紫色に見える。
従属栄養原核生物とは異なり、シアノバクテリアは内膜を持っている。これらは、光合成が行われるチラコイドと呼ばれる平らな嚢である。紅藻類・緑藻類・植物などの光独立栄養性真核生物は、シアノバクテリアに祖先を持つと考えられているクロロフィルのオルガネラで光合成を行う。これは、はるか昔に内部共生によって獲得され、その後、真核生物のこれらの内部共生シアノバクテリアは進化させ、葉緑体・色素体・エチオププラスト・ロイコプラストなどの特殊なオルガネラに分化させた。
② 植物は根から水に溶けたアンモニアイオンNH₄+や硝酸イオンNO₃-を、動物はタンパク質を消化してアミノ酸(CHNOなどで構成)の状態でしか窒素を吸収することができない。硝化細菌は、土壌窒素のうち最も還元された形のアンモニアNH 3を最も酸化された形の硝酸塩NO3−に変換する窒素循環の微生物で、無機窒素化合物の酸化からエネルギーを生成する化学石栄養生物でもある。硝化細菌には、アンモニア酸化細菌と亜硝酸酸化細菌の2種類が存在する。硝化細菌は、硝化の酵素反応に必要な複雑な内膜系を有している。その一般的な硝化経路は、アンモニアモノオキシゲナーゼと言う酵素でアンモニアNH 3を生合成的硝化の中間体であるヒドロキシルアミン硝酸塩NH2OHに酸化し、ヒドロキシルアミンオキシドレダクターゼという酵素でヒドロキシルアミン硝酸塩から硝酸HNO3に酸化し、 最終的には
2NO3 + 6H2 → N2 + 6H2O と段階を踏みN2を放出する。
③ 窒素固定菌「アゾトバクターAzotobacter」は、空気中の「窒素」を利用して窒素化合物である「アンモニア」を合成できる微生物の総称である。空気中の窒素からアンモニアなどの窒素化合物を合成することを「窒素固定」と言う。
窒素固定細菌は、植物との関係性によって大きく2種類に分けられる。特に大豆などのマメ科植物やヤマノイモ科植物を宿主として共生し根粒を作る「根粒菌(リゾビアrhizobia)」と「葉粒菌」などの「共生窒素固定菌」を言う。
共生窒素固定菌は、単独では空気中の窒素固定をしない。宿主植物と共生することで初めて窒素固定が行われる。
常緑低木マンリョウ(万両)は、 センリョウやヤブコウジなどとともに赤い果実が冬に実るので、正月の縁起物として玄関先に好んで植えられる。その常緑の葉は、枝の先端で互生し、質は厚く両面とも無毛で、表は濃緑色で光沢があり、長さ5-15cm、幅2-4cmの長楕円形、葉柄は5-10mm程度、先端は尖り、縁には波状の鋸歯が有り裏側に巻き込む。その波打つ緑の葉の鋸歯の間のへこみ部分にある小さな隆起には、内腺点(内腺体・葉粒とも呼ばれる)があり、共生細菌が共生している。共生細菌の葉粒菌は、茎の先端や芽の内部に巣食い、枝・葉・花・実・種子に感染し、次世代に受け継がれる(垂直感染)。窒素固定を行っている。親から子へ細菌を伝播するという意味で、葉粒菌は根粒菌よりも植物との結びつきが強いと言える。
実際に、人工的に無細菌状態(植物から細菌を取り除いた状態)にしたマンリョウの仲間の株を用いて種子から育ててみると、枝や茎が萎縮して正常に成長することができなかったという実験結果も報告されていた。
マンリョウの葉の縁には大きく波打つ膨らんだ鋸歯があり、その膨らみに共生菌が住む。この共生菌は植物の根に小さな瘤を作って共生する根粒菌に対して、葉に共生するため葉粒菌と呼ばれ、昔は根粒菌と同様に窒素を固定すると考えられていた。しかし、その葉粒菌がマンリョウの葉の縁で窒素固定に参加していないことが近年の研究で立証された。研究によれば、マンリョウがこの葉粒菌と共生することによって得られる二次代謝産物は、マンリョウを植食性の昆虫や動物から守る効果があるということが指摘されている。
マンリョウは園芸樹や果樹として、オーストラリアやアメリカ、太平洋諸島へ移植され、侵略的外来種となった。日陰で10年、日向ならわずから2年で成木となる。日向に植林された成木は、最高400の果実を産出でき、日陰でも多数の実をつける。種子には休眠期間がなくすぐに発芽し、若木は何年もの間、日陰のような悪条件のもとでも生存する。その後日が当たると急速に成長し実生繁殖力が高まり、果実が鳥に食べられて種子散布され、思いがけない場所にも生え広がる。このマンリョウの活力は、どこから生まれるのだろうか。
https://jspp.org/hiroba/q_and_a/detail.html?id=907
もう1つは「単生窒素固定菌」といい、アゾトバクターやクロストリジウム、シアノバクテリアなどが含まれる。これらは土壌や水などの中に広く生息し、単独で窒素固定を行う。窒素固定シアノバクテリアは、シアノバクテリアのうち大気中の分子状窒素N2をアンモニアに還元する窒素固定能力をもつものの総称である。窒素固定の能力は、単細胞・糸状体種の多くのシアノバクテリア分類群に散見されるが、普遍ではない。大部分は単生であるが、真核藻類・地衣類・シダ植物・裸子植物などと共生する種も知られている。環境に窒素化合物が不足する条件で主としてニトロゲナーゼ(nif)遺伝子が発現することで窒素固定活性が出現する。シアノバクテリアは、核膜に包まれた核をもたない原核生物で、真核生物の葉緑体のような細胞内小器官をもっていない。しかし、細胞内には真核生物の葉緑体の内膜構造に相当する「チラコイド」が存在し、細胞全体が一つの葉緑体のように機能して光合成を行う。
原核生物は真正細菌と古細菌に分けられるが、光合成を営む真正細菌を大別すると「シアノバクテリア」と「光合成細菌(光合成生物のうち、真核生物を除く)」に分けられる。両者の行う光合成には、有機物の合成に必要な電子の源として水H2Oが利用されるか、されないかの違いがある。シアノバクテリアの場合には水が利用され副産物として酸素が発生する(酸素発生型光合成)。光合成細菌の場合には水を利用しないため酸素が発生しない(酸素非発生型光合成)。なお、光合成器官として葉緑体を備えた真核光合成生物の営む光合成は、その進化の過程からもシアノバクテリアと同じ酸素発生型となる。
シアノバクテリアと光合成細菌の間には光合成色素にも違いがある。シアノバクテリアの場合にはクロロフィルaが中心色素となっているが、光合成細菌の場合にはバクテリオクロロフィルが光合成色素である。なお、クロロフィルとバクテリオクロロフィルは近縁の化合物である。ちなみに、真核光合成生物ではクロロフィルaが中心色素として機能しているが、クロロフィルbやクロロフィルcが補助色素として関与する場合がある。シアノバクテリアの場合には、クロロフィルaに加え、クロロフィルb・クロロフィルd・クロロフィルfなど別の種類のクロロフィル類が含まれる種があることが知られている。しかも、嫌気・微嫌気条件でのみ窒素固定活性を発現する種も存在する。ただし、単生窒素固定菌の中でもいくつかの種類は、植物の根圏の土壌や根の表面に集まって生息し、ゆるい共生関係を持つ真正細菌もある。
1980年代後半以降になって、根粒は作らないものの植物体内に生息して共生関係を担う、窒素固定細菌についての研究が促進されている。これらは「植物体内窒素固定細菌(エンドファイトendophyteの一種)」と呼ばれている。エンドファイトとは、内部共生生物であり、多くの場合、細菌または真菌であり、明らかな病気や障害を引き起こすことなく、そのライフサイクルの少なくとも一部で植物内に共生している。エンドファイトはいたるところに存在し(phyte)、これまでに研究されたすべての種類の植物で発見されている。しかし、エンドファイトと植物の関係の多くは未だ解明できていないが、例えば植物の内部に生息して窒素を固定する細菌として共生関係を築き、植物が窒素を吸収しやすい形に変換する。これにより、植物が窒素肥料に頼らずに成長することができる。
エンドファイトは、植物や微生物にコロニーを形成する微生物の内部共生グループであり、微生物や植物増殖培地から容易に分離できる。それらは、アルカロイド・フェノール酸・キノン・ステロイド・サポニン・タンニン・テルペノイドなどの新規の生理活性二次代謝産物の貯蔵庫として、抗菌・抗昆虫・抗癌、その他多くの潜在的特性を秘めながら機能している。一部のエンドファイトは、宿主の成長と栄養素の獲得を促進し、旱魃などの環境ストレスに耐える植物の能力を向上させ、昆虫・病原体・草食動物に対する植物の抵抗性を高めることにより生物的ストレスの減少に貢献しているようだ。エンドファイティックendophytic細菌や真菌は盛んに研究されているが、エンドファイティック古細菌は、特に植物のコアマイクロバイオームcore microbiomeに潜在的に直結するとして、人体や土壌などの特定の環境に存在する微生物群(細菌・ウイルス・真菌など)の遺伝情報の全体の中核になるとして研究され、植物の成長促進に果たす役割についての試験データが蓄積され活用されてきている。例えば、ヒトの体内や体表で生息する微生物とその遺伝情報の総称して「ヒトマイクロバイオームhuman microbiome」と呼ぶ。腸内細菌叢には、ヒトの細胞数に近い約40兆個の細菌が存在し、皮膚・口腔・鼻腔・膣などにも特に多くの微生物が存在している。
(「microbiome」は、特定の環境、特に人体や土壌などに存在する微生物群【細菌・ウイルス・真菌など】とその遺伝情報の全体を指す用語であり、微生物学や生物学、医学などの分野で頻繁に使われている。)
目次へ