顕生代Phanerozoic eon
TOP 車山お知らせ 歴史散歩 旅ネット信州
| 目次 |  |
顕生代 肉眼で見える生物が生息しし始めた地質時代 |
|
| 1)アフリカ大地溝帯とゾウ類 | |
| 2)漸新世にみられる風景 | |
| 3) 漸新世が語ること | |
| 4)漸新世プレートテクトニクス |
海洋の深層水で酸素濃度
海洋無酸素事変Oceanic Anoxic Event(OAE)とは、地質時代において海洋の広範囲が酸素欠乏状態になった現象を指す。この現象は数百万年にわたって続くことがあり、主に次のような原因が考えられている。全球的な温暖化により、海洋の水温が上昇し、酸素の溶解度が低下した。プランクトンなどの有機物が大量に生産され、これが海底に沈殿して分解される際に大量の酸素が消費された。また海洋の循環が変化し、深層水における酸素供給が減少した。
OAEは主に中生代のジュラ紀(約2億180万年前~約1億4500万年前)から白亜紀(約1億4,500万年前~6,600万年前)にかけて発生しており、この時期には多くの海洋生物が絶滅した。例えば、珊瑚や多くのプランクトンが影響を受け、海洋生態系に大きな変動が生じた。
海洋の深層水の酸素濃度は一般的に低い。これは主に深層水が地表から遠く、酸素の補給源から離れているためである。深層水は長期間にわたり酸素供給がないため、その濃度は減少する。酸素濃度が比較的高い地域も存在するが、全体的には浅層水に比べて酸素が少ない。
深層水の循環は、「大洋循環」と呼ばれる地球規模の運動によって行われる。大洋循環は、主に温度差と塩分濃度差による「熱塩循環」と呼ばれるプロセスで稼働される。冷たい水は一般に密度が高く、沈降する傾向がある。北大西洋や南極海などの極地方で冷やされ、沈み込んだ水が深層水として赤道付近へ流れ、また表層に戻るというサイクルを繰り返す。 この大規模な水の循環は、地球の気候や生態系に重大な影響を与えている。この全体のプロセスは数百から数千年をかけて完了すると見られている。これにより、地球全体の海洋がゆっくりと混ざり合い、栄養素や酸素の分配が行われている。
海洋の深層水で酸素濃度が比較的高い地域の一つは南極周辺の海域で、特に南極底水Antarctic Bottom Waterは、酸素濃度が高いことで知られている。これはいくつかの要因によって説明されるが、南極周辺の海水は非常に冷たいため、表層水が冷却されて密度が増し、深層へ沈降する。この過程で、冷たい水は酸素を多く含むことができる。また、表層水が冷たいと、その溶存酸素濃度が高くなる。冷たい海水は、大気中の酸素を取り込む能力が高いからである。この酸素が豊富な水が沈降することによって、深層水に酸素が供給される。南極から北へ流れる深層水は、海流の影響で比較的速い速度で移動し、長い間酸素を消費することなく高酸素濃度を保ったまま広がる。 これらの要因が組み合わさることで、南極周辺の深層水は他の地域と比べて酸素濃度が高く保たれている。
それでもこれでも深層水の酸素濃度は表層水より低い。それは相対的に見て高いという意味である。南極以外にも、北大西洋の一部も酸素濃度が高めの深層水を持つ地域として知られている。北大西洋深層水North Atlantic Deep Waterは、グリーンランド周辺で沈み込む際に酸素を多く含むためである。
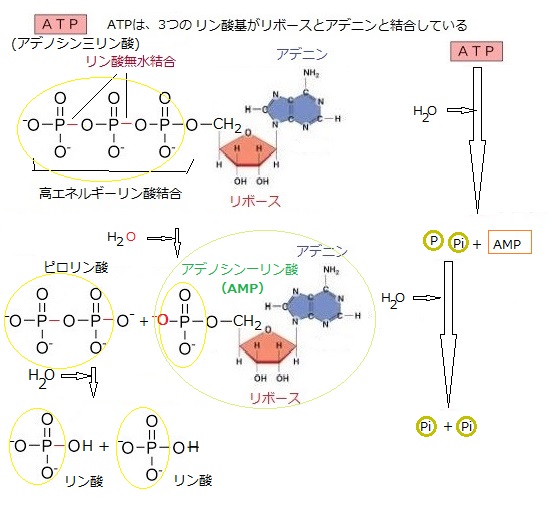 |
| ATP(アデノシン三リン酸)の分子構造のアデニンはプリン塩基(DNAやRNAを構成する核酸の一部で、アデニンAとグアニンGなどがある)の一種で、他の分子との相互作用において重要な役割を果たす。 植物がATPを生成するために必要なリン(リン酸)・アデニン(核酸を構成する5種類の主な塩基のうちの一つで、生体内に広く分布する有機化合物)・リボース(糖)は、次のような過程で取り込まれ、合成される。 リン酸は、植物が土壌から吸収する主要な栄養素の一つである。植物の根が土壌から水と共にリン酸塩を吸収し、これがリン酸として体内に取り込まれる。リン酸はATPのリン酸基として重要な役割を果たす。 アデニンは、窒素を含む塩基で、植物は窒素源からこれを合成する。窒素は、大気中の窒素を固定する窒素固定細菌や、肥料から供給される。植物は窒素を取り込み、複雑な生化学反応を経てアデニンを合成する。そのアデニンの構造が、酵素や受容体との結合において、特定の認識サイトに適合することで、ATPが正確に機能することができる。 リボースは、糖の一種で、植物の光合成過程で生成される。光合成のカルビン・ベンソン回路において、二酸化炭素が固定されてリボースなどの糖が生成されるが、これにより、リボースはATPの構造に組み込まれる。リボースは五炭糖(化学式はC5H10O5)でリボ核酸(RNA)の構成成分でもあり食品添加物や運動後の栄養補助食品としても利用されている。 そのリボースの五炭糖による五角形の構造がATP全体の安定性を保つための基盤となる。リボースの炭素原子は、アデニン(塩基)とリン酸基に結合して、ATPなどのヌクレオチドを構成する。この結合は分子全体の安定性を保つために重要である。また、これにより、アデニンとリン酸基が結びつき、ATP分子全体の一体性が保たれる。ATPのエネルギーは主にリン酸基間の高エネルギー結合に由来するが、リボースがなければこれらのリン酸基を安定的に配置することができない。リボースは、ATP分子が効率的にエネルギーを供給できる形状を維持するために不可欠である。 ATPは、リン酸基が結合した物質、そのヌクレオチドが鎖のように連なることによりアデノシン三リン酸はリン酸基供与体としても機能する。 リボースはATPの構造と機能において中心的な役割を果たしており、細胞内でのエネルギー代謝に重要な役割を果たしている。 具体的には、アデニンがリボースと結合してアデノシンを形成し、そのアデノシンに3つのリン酸基が結合している(アデノシン三リン酸)。この構造により、ATPはエネルギーを貯蔵し、必要に応じて放出することができる。 ATPの3つのリン酸基同士の結合は「高エネルギーリン酸結合」と呼ばれ、多量のエネルギーが蓄えられている。この結合が切断されると、エネルギーが放出され、さまざまな生体反応に利用される。 またADPにリン酸基が再結合することで、ATPが再生成される。このプロセスは、呼吸や光合成などの代謝反応によって行われる。 ATPは生体内でエネルギー通貨として機能し、様々な生化学反応において重要な役割を果たしながら、物質の能動輸送なども担う。 |
海底では、有機物(例えば、プランクトンの死骸など)が沈殿し、微生物によって分解される。好気性菌は酸素に基づく代謝機構を備えた生物である。細胞の呼吸で知られた過程の中で、好気性菌は、たとえば糖や脂質のような基質を酸化してエネルギーを得るために、酸素を利用する。好気的呼吸における、ブドウ糖(単糖)の酸化は良い例である。この際、好気性微生物(酸素を利用する微生物)がグルコースC6H12O6を酸素O2によって酸化し、エネルギーを得る。このエネルギーは微生物の生命活動を支えるために使用される。グルコースC6H12O6が酸素O2によって酸化される反応は、化学種が電子を失うプロセスを指す。具体的には、グルコースの炭素原子が電子を失い、二酸化炭素CO2として放出される(酸化反応)。同時に、酸素O2が還元されて水H2Oを生成する。還元とは、化学種が電子を得るプロセスを指す。酸素分子は電子を受け取り、水分子に変わる(還元反応)。
C6H12O6 + 6O2 ⇒
6CO2 + 6H2O + エネルギー(ATP)
(グルコースと酸素から二酸化炭素、水、そしてエネルギーATP【アデノシン三リン酸】が生成される反応により獲得したエネルギーを、植物はさまざまな生命活動を行うために使用する。)
ATPは、細胞内でエネルギーのやり取りに使われる重要な分子で、そのATPの化学式は C10H16N5O13P3 。
ATP分子は、アデニン、リボース(糖)、および3つのリン酸基から構成されている。これらの構成要素が結びつき、エネルギーを貯蔵および供給するための高エネルギー結合を形成する。植物において、ATPは光合成および細胞呼吸の過程で生成され、さまざまな代謝活動に必要なエネルギーを提供する。例えば、光合成では、光エネルギーを化学エネルギーに変換してATPを生成し、それを使ってグルコースを合成する。また、細胞呼吸では、グルコースの分解によってATPが生成され、細胞内のエネルギー源となる。
この化学式において放出されるエネルギーはブドウ糖1モルあたり約2,880 kJであり、このエネルギーは、ブドウ糖1分子につき38分子のアデノシン二リン酸から38分子のアデノシン三リン酸(ATP)を再生することで保存される。これは糖1分子当たりから嫌気的反応によって産み出されるエネルギーの19倍も多いエネルギーである。真核生物(細菌をのぞくすべて)は、この好気的呼吸によって、膜透過のための能動輸送に必要となる分のエネルギーを除いて、アデノシン二リン酸からアデノシン三リン酸を正味36分子を得ることができる。この反応において、ブドウ糖の酸化には酸素が使われ、水が作られることがわかる。好気性生物が必ずしもこの反応系をすべて利用するものではないが、嫌気性生物よりもATP生産量が多く、生存に有利に働くことが示される。
好気性生物は酸素を利用してエネルギーを生産する生物であり、そのエネルギー生産性は非常に高い。好気性生物は酸素を利用して、グルコースを完全に二酸化炭素と水に分解する。この過程で、効率的で大量のエネルギー変換が可能となる。好気性呼吸は主に細胞内のミトコンドリアで行われる。ミトコンドリアは「細胞の発電所」とも呼ばれ、その構造と機能がATPの大量生産を支えている。好気性呼吸では、1分子のグルコースから最大約38分子のATPを生成する。これに対して、酸素を使用しない嫌気性呼吸では、1分子のグルコースからわずか2分子のATPしか生成できない。この大きな差が、好気性生物のエネルギー生産性の高さを示す。高効率で大量なエネルギー生産能力は、好気性生物が複雑な多細胞生物へと進化するための決定的な基盤となった。これにより、高度な生物機能や行動が可能となった。このように、好気性生物のエネルギー生産性の高さは、ミトコンドリアの機能、酸素の利用、ATPの高効率な生成に依存し、それが生物の進化に大きく貢献した。しかしながら、大量に好気性生物が出現し活動すれば、海洋無酸素事変が再発する。
 暁新世-始新世温暖化極大
暁新世-始新世温暖化極大5,500万年前に『暁新世-始新世温暖化極大Paleocene/Eocene Thermal Maximum(PETM)』が発生した。この急速な全球的温暖化現象は、新生代に発生した温暖化現象では史上最大規模のもので、全球の平均気温が5〜 9℃上昇したほか、海洋の炭酸塩(主に炭酸カルシウムCaCO3)の溶解による海洋酸性化も発生した。湿度も高かった。その原因として北大西洋での海底火山活動やそれに伴う海底に大量に埋蔵されているメタンハイドレートmethane hydrateの融解などの温暖化ガスの大量放出があり、元の二酸化炭素濃度に戻るのに3万年を要したとされる。
この出来事は霊長類の進化史にも大きく影響したと見られている暁新世末期にやや低下した気温は始新世では再び温暖化に転じ、新生代では最も高温の時代になった。
極地付近にも氷床はなく、5300万年前のクジラによく似た半水生哺乳類が進化を遂げた古代クジラが南極の海域を泳いでいたのは、約4900万年前の「始新世」だった。南極周辺に生息していた世界最古のクジラが発見された。それも多数の化石が出土している南極半島の堆積層からは、長さ60cmのクジラのアゴ骨から、そのクジラの推定全長は最大6mで、口いっぱいに歯が並び、おそらく巨大なペンギンやサメ、大きな硬骨魚を食べていたと考えられている。そのような動物の骨もアゴ骨と一緒に出土している。
広範な火山活動が見られた。火山の噴火によって大量の二酸化炭素CO2が大気中に放出され、その結果、地球全体の温暖化が進んだ。特に、約5,800万年前の暁新世後期に形成された大規模な火山活動の地域「北大西洋火成地域North Atlantic Igneous Province」の大規模な火山活動がCO2の主要な供給源と見られている。この北大西洋火成地域は、地中海プレートと北アメリカプレートの衝突によって引き起こされたと考えられている。この衝突により、地中海プレートが北アメリカプレートの下に沈み込み、マグマが地表に噴出した。この火山活動は、約5,800万年前から約5,600万年前まで続き、この期間中、大量の玄武岩質の溶岩が噴出し、広大な範囲にわたって火山岩を堆積させた。特に、アイスランドやグリーンランドなどの地域で顕著な火山活動が痕跡が見られる。このアイスランドを中心とする北大西洋の大規模な火成地域で、古第三紀には、州は大きな玄武岩質溶岩平原であるテュレアン高原を形成した。面積は少なくとも130万km2、体積は660万km3に広がっていた。
地質学的な研究により、この地域の火山活動が当時の気候変動にも影響を与えたことが示されている。テュレアン高原は北大西洋の開通時に分断され、北アイルランド・西スコットランド・フェロー諸島・アイスランド北西部・東グリーンランド・ノルウェー西部、および北大西洋の北東部に位置する多くの島々に現在も痕跡を遺す。
暁新世から始新世への温暖化極大化(EOT)の時期には、大規模な火山活動が発生した。この火山活動の規模は非常に大きく、地球規模の気候変動を誘発した。この時期には、地殻の移動やプレートの沈み込みが活発化し、特に、インドプレートとユーラシアプレートの衝突によって、ヒマラヤ山脈の形成が進行していた。ヒマラヤ山脈の大規模な隆起は、大陸同士の衝突による。現在のインドプレートは、かつては独立した大陸で、インド洋の南半球に位置していた。それがプレートの動きとともに北上を続け、始新世初期の約5,000万年前頃にユーラシア大陸と衝突し始めた。こうして両大陸の間にあった海は消滅し、海に堆積していた堆積岩は大陸に挟み込まれて陸上へと隆起した。
 全捄規模のプレートテクトニクスによるマントルの上昇と冷却は、地下深くからのマグマの上昇を促進し、火山活動を頻発させた。これにより、大量の二酸化炭素が大気中に放出され、温室効果が強化された。海流の変化も火山活動の活発化に寄与した。例えば、南極海のゲートウェイの開放により、海水の循環が変化し、気候に影響を与えた。このような大規模の環境変化は、大気中の二酸化炭素濃度の増加とともに、地球全体の気温上昇を引き起こし、地球の気候システムに大きな影響を与え、多くの生態系を混乱させた。
全捄規模のプレートテクトニクスによるマントルの上昇と冷却は、地下深くからのマグマの上昇を促進し、火山活動を頻発させた。これにより、大量の二酸化炭素が大気中に放出され、温室効果が強化された。海流の変化も火山活動の活発化に寄与した。例えば、南極海のゲートウェイの開放により、海水の循環が変化し、気候に影響を与えた。このような大規模の環境変化は、大気中の二酸化炭素濃度の増加とともに、地球全体の気温上昇を引き起こし、地球の気候システムに大きな影響を与え、多くの生態系を混乱させた。
暁新世から始新世への温暖化極大化(EOT)の時代には、いくつかの重要な海洋ゲートウェイの変化があった。これらの変化は、地球の気候システムに大きな影響を与えた。南極周極流Antarctic Circumpolar Current(ACC)は、南極大陸と南アメリカ大陸の間に形成された海峡を通る海流であるが、この海峡が開いたことで、南極大陸の冷たい海水が北上し、地球全体の海水温を引き下げた。
北極海のゲートウェイ、例えば、グリーンランド・スコットランド海嶺やプロト・フレーム海峡の変化により、北極海と北大西洋の間の海水の流れが変わり、北大西洋の海流が弱まり、北半球の気候に影響を与えた。特に、北大西洋の海洋表面温度が低下し、北大西洋循環North Atlantic circulationの始動に影響を及ぼした。
大西洋子午面循環
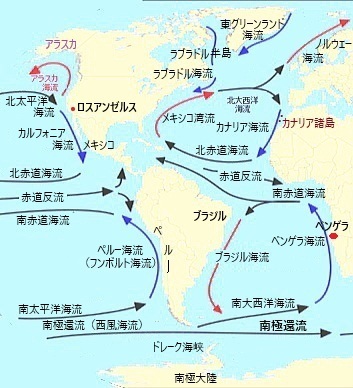 南極周極海流(南極還流)Antarctic Circumpolar Current(ACC)は、地球上で最も強力かつ重要な海洋循環の一つで、南極大陸を取り囲むように流れており、南大洋全体にわたって影響を及ぼしている。 海流は周極であり、これにより暖かい海水が南極大陸から遠ざかり、南極大陸がその巨大な氷床を維持することを可能にしている。 ACCは南極周辺の南大洋を一周する海流で、約24,000kmにわたる非常に広大な領域をカバーしている。ACCは、南緯40度から60度の間の南極前線地帯Antarctic Convergence Zoneに位置している。 |
大西洋子午面循環と北大西洋循環の違い 大西洋子午面循環Atlantic Meridional Overturning Circulation(AMOC)と北大西洋循環North Atlantic Circulationは、いくつかの重要な要素で異なる。 AMOCを直訳すれば「大西洋南北逆転循環」、つまり大西洋の主要な海流システムで、地球の海洋循環システムの構成要素であり、気候システムに重要な役割を果たしている。 AMOCには、天候・気温・塩分の変化によって駆動される地表および深海流の大西洋海流が含まれている。これらの海流は、主要な海流の流れを含む全球の熱塩循環の半分を占めており、残りの半分は南極海逆転循環である。 スケールと範囲 AMOCは大西洋全体を含む広範囲な循環で、赤道からグリーンランド、さらには南極まで及ぶ。 北大西洋循環は北大西洋の一部、特にヨーロッパや北アメリカの沿岸を中心とした循環である。 構造と動き AMOCは、主に南北方向の深層循環を含み、暖かく軽い表層水が北に向かい、冷たく重い深層水が南に向かう大規模な循環である。 北大西洋循環は主に表層水の動きを含み、風や地形による影響が大きい。 子午面とは 子午面meridional planeは、地球上で北極から南極に至る面を意味する(meridional ;「南北」の意味)。 具体的には、経度0度(本初子午線)やそれに平行な任意の経度の面planeを指す。この面を基に、北半球と南半球の循環パターンを研究する。 子午面循環は、これらの縦の面を通る海水や大気の大規模な動きを説明する。特に、AMOCのように南北方向の水の移動に焦点を当てた循環を記述する際に重要となる。 AMOCと北大西洋循環は、異なる規模とメカニズムを持ちながらも、地球規模の気候に大きな影響を及ぼしている。 |
このプロセスは非常に複雑で、他にもさまざまな要因が関与しているが、海洋表面温度と塩分濃度の変化が北大西洋の冷却とAMOCの変化に大きく関与している。 このような海洋ゲートウェイの変化は、北半球の気候に対して大きな影響を与え、高緯度地域での冷却と気温の勾配の増加を引き起こした。これにより、北半球の気候はより寒冷なものとなり、最終的には北半球の氷河期が始まる契機となる。
北大西洋の海洋表面温度の低下と大西洋子午面循環(AMOC)の始動に影響を及ぼした主な原因は、気候変動であった。地球温暖化により、海水の温度が上昇し北極付近の氷が溶け、北大西洋の海洋表面温度が低下した。これにより、AMOCの始動に影響を及ぼした。海洋循環の変化が、熱を効率的に地球全体に分散させる能力が低下させた。この温暖化は、気候変動や海水環境に大きな影響を及ぼした。温暖化により、気温が急激に上昇し、極地や高山地域で氷河が溶けた。これにより、海面上昇が引き起こす。
北大西洋の表面塩分濃度の低下の原因は、降雨量の増加と氷河の融解であった。降雨量が増えると、海水が希釈され、塩分濃度が低下する。また、氷河が融解すると、海水に溶け込んだ水が塩分濃度を下げる。 これらによる海洋ゲートウェイの変化は、地球の気候システムに大きな影響を与え、特に南極大陸の氷床の拡大や北半球の気候変動にも関与した。
海洋に溶解している炭酸塩は主に炭酸カルシウムCaCO3として存在している。炭酸塩が海洋に溶解する過程は、主に風化と溶解、陸上での風化作用により、石灰岩やその他のカルシウム含有鉱物が化学的に分解され、炭酸カルシウムが水に溶ける。これにより、カルシウムイオン Ca2+と炭酸イオンCO32-が生成される。風化によって生成されたカルシウムイオンと炭酸イオンは河川によって海洋に運ばれる。海洋に運ばれたカルシウムイオンと炭酸イオンは、海水中で再び反応して炭酸カルシウムを形成することがある。
この炭酸カルシウムは、一部は溶解し、一部は沈殿する。海洋生物、例えば、貝やサンゴなどは、炭酸カルシウムを利用してその殻や骨格を形成する。これにより、炭酸カルシウムがさらに海洋中に増える。炭酸カルシウムを含む海洋生物が死亡したり、その殻が破壊されたりすると、炭酸カルシウムは海底に沈積する。これが堆積岩としての石灰岩などを形成する。 このようなプロセスを通じて、炭酸塩は海洋中に溶解し、さまざまな化学反応と生物活動を通じて循環する。
海洋に溶解している炭酸カルシウムCaCO3は、海洋の炭素循環において重要な役割を果たしている。この溶解した海洋中の炭酸カルシウムは、炭酸塩システムの一部として存在し、海水のpHを安定化させる役割を果たす。このシステムには炭酸イオンCO32-、重炭酸イオンHCO3-、および二酸化炭素CO2も含まれる。多くの海洋生物、例えば、サンゴ・貝・プランクトンなどは、炭酸カルシウムを利用してその骨格や殻を形成する。これにより、炭酸カルシウムは生物の成長と維持に不可欠な物質となる。 炭酸カルシウムを含む生物が死んだり、その殻が破壊されたりすると、これらの物質は海底に沈降し、堆積物として積み重なる。長い時間の中で、これが石灰岩などの堆積岩を形成する。
海洋中での炭酸カルシウムの溶解と再沈殿は、地球規模の炭素循環の一部で、これにより、大気中の二酸化炭素が海洋に吸収され、長期的な炭素の貯蔵庫となる。 海洋に溶解している炭酸カルシウムは、生物学的、化学的なプロセスに多大な影響を与え、地球全体の生態系にも重要な役割を果たしている。
炭酸カルシウムの溶解と海水の酸性化には密接な関係があるが、少し異なるプロセスを経る。一般的に、温度が上がると炭酸カルシウムの溶解度は増加するが、炭酸カルシウムは海水中では比較的溶解しにくい物質であり、温度の影響はそれほど大きくはない。 海水温が上昇すると、二酸化炭素の溶解度が減少する。これは、温かい水の分子の方が活発に運動するため、冷たい水よりも二酸化炭素を少なく保持する傾向がある。
大気中に放出された二酸化炭素が増えると、海洋により多くの二酸化炭素が吸収される。これが海水中で炭酸H2CO3を形成し、炭酸水素イオンHCO3-と水素イオンH+に解離する。さらに炭酸水素イオンHCO3-がさらに解離して炭酸イオンCO32-と水素イオン H+にイオン化する。
反応式は H2CO3 ⇌ HCO3- + H+
HCO3- ⇌ CO32- + H+ (炭酸は段階的にイオンに分離する)
これが、海水のpHを下げて酸性度を高める。また水素イオンの増加は炭酸カルシウムの溶解を促進する。したがって、海水温の上昇そのものが直接炭酸カルシウムのイオン化を速めるというよりも、二酸化炭素の影響が海洋酸性化を引き起こし、結果として炭酸カルシウムの溶解が進む。
二酸化炭素CO2が海洋H2Oに溶け込むと、水と反応して炭酸H2CO3を生成する。この炭酸はさらに解離して水素イオンH+と炭酸イオンHCO3-を生成する。この過程によって海水のpHが低下し、海洋酸性化が進行する。
固体の炭酸カルシウムCaCO3の水素イオンH+による溶解により、カルシウムイオンCa2+と炭酸イオンHCO3-が生成される。これが海洋生態系において貝殻やサンゴなど、炭酸カルシウムを主成分とする生物にとって致命的な脅威となる。海水の温度上昇と酸性度の増加が海洋生態系に大きな影響を与える。特に、カルシウムを多く含む生物、例えば、サンゴや二枚貝は、酸性度の上昇によって成長が妨げられる。
北大西洋火成岩地域
「暁新世から始新世にかけての温暖化極大化」は、暁新世末期の約5,300万年前に始まった。この時代の温暖化は、大規模な火山活動が大量の二酸化炭素CO2を大気中に放出し、温室効果を強化したことが主な要因と見られる。暁新世から始新世への温暖化極大化の時代には、世界的に火山活動が一段と活発化した。この時代の温暖化は、主に北大西洋火成地域の火山活動によるものとされている。
北大西洋火成岩地域North Atlantic Igneous Province(NAIP)は、いくつかの大陸プレートにまたがっている。これは、地質学的な用語で、北大西洋周辺の火山活動が集中している地域を指す。主に北アメリカプレートとユーラシアプレートの境界に位置し、更に一部はグリーンランドや北大西洋の中央部を含む。北アメリカプレートでは、グリーンランド東部やカナダの一部、ユーラシアプレートでは、イギリス・ノルウェー・アイルランド、特にアイスランドでは、北アメリカプレートとユーラシアプレートの境界に位置し、2つのプレートが離れていく場所にあるため、独特の地質学的特性を示し、その火山活動は一段と活発である。
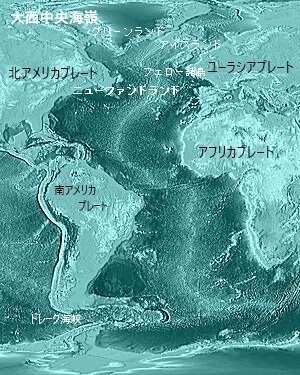 |
| 大西洋中央海嶺は、地球のプレートテクトニクスの重要な要素で、大西洋の中央を南北に貫く巨大な海底山脈である。 大西洋中央海嶺は、リソスフェアプレート(地殻と上部マントル)が拡大する場所で、この拡大により、新しい海洋地殻(海嶺)が形成される。 この海嶺では、マグマが上昇し、冷えて固まることで新しい地殻が形成される。この過程は「海底拡大」と呼ばれ、大西洋中央海嶺の特徴は地震活動が活発な地域となり、特に新しい地殻が形成される過程で頻繁に地震が発生する。 海嶺には多くの海底火山が存在し、これらの火山活動でも新しい地殻が形成される。大西洋中央海嶺の拡張速度は年間約2〜5cm程度とされているが、この速度は地質学的時間スケールでみると非常に速い。 地球のプレートは常に動いている。拡張するプレートがあれば、収束するプレートもある。大西洋中央海嶺でプレートが東西に拡大する一方で、太平洋プレートは西向きに移動している。 太平洋プレートはその西側の境界、例えば、日本海溝で北アメリカプレートの下に沈み、マリアナ海溝でフィリピン海プレートの下で収束するなど、太平洋プレートの西側で他のプレートの下に沈み込む。これは、地球規模で見ると太平洋が徐々に狭まる要因の一つとなる。 このように、プレートテクトニクスの多岐にわたる動きによって、大西洋中央海嶺の拡張が間接的に太平洋の広さに影響を与えるようにも見られる。 大西洋中央海嶺は、プレートテクトニクス理論の重要な証拠の一つで、海底拡大とプレート移動のメカニズムを示す場所として、貴重なな研究対象となる。 北大西洋火成岩地域は、暁新世末期の約5,600万年前〜約5,380万年前にかけて形成された広大な火山活動地域で、現在の北アメリカ大陸のカナダのラブラドール地方やニューファンドランドなどや、グリーンランドの特に東部の地域に広がる火山岩層や、現在でも活発な火山活動が見られアイスランド、フェロー諸島などアイスランドとスコットランドの間の地域に、火山岩が広がっている。 更に北ヨーロッパの一部を含む。 スコットランドの特に西部のハイランド地方やヘブリディーズ諸島で、北大西洋火成岩地域に関連する火山岩が見られる。 その生成は、大規模なマントルプルームの活動によるもの、このプルームは、地下のマントルが上昇し、地表近くで冷却・固化して火山岩を形成した。このプロセスは、大陸の分裂やプレートテクトニクスの影響を受けていた。 その大規模な火山噴火、地熱活動は、大量の玄武岩質溶岩の噴出を伴い、それにより、広大な玄武岩台地や火山岩の層が形成された。また、この地域は、地熱エネルギー資源としても注目されている。 この地域の研究は、地質学や地球物理学の分野で非常に重要であり、地球の歴史やプレートテクトニクスの理解には欠かせない地域である。 |
北大西洋火成岩地域 (NAIP) は、大西中央海嶺 Mid-Atlantic Ridge)の近くに位置しているが、厳密には大西中央海嶺そのものには属してはいない。大西中央海嶺は、大西洋の中央に位置する海底山脈であり、プレート境界のひとつである。ここでは新しい海洋地殻が生成され、大西洋が徐々に広がっていくプロセスが進行している。
一方、大西中央海嶺は、パンゲア大陸が分裂を始めたことに起因する。この分裂は、当時存在していた超大陸パンゲアの崩壊によって引き起こされた。パンゲアの崩壊により、大陸プレートが複数の方向に引き伸ばされ、新しい海洋地殻が形成された場所が生まれた。パンゲア大陸は地球上のすべての大陸が一つに結合した超大陸で、内部の熱と地殻の運動により引き裂かれた。
大陸が引き離される過程で、リフトゾーンrift zone(引き裂き帯)が形成され、マントルの溶岩が上昇して新しい海洋地殻が生成されました。このリフトゾーンが大西中央海嶺として進化し、大西洋が徐々に広がっていくことになった。このプロセスのきっかけは、地球内部の熱とプレートの運動にあ、地球内部の熱エネルギーがマントルの対流を引き起こし、プレートがゆっくりと移動することで、大陸が引き裂かれた。このような地質学的プロセスにより、現在の大西中央海嶺が形成され、大西洋が拡大していった。
一方、北大西洋火成岩地域は、パンゲア大陸の分裂によって形成された地質学的な地域であり、主に大陸プレートの一部に属している。この火成岩地域は、大規模なマグマ活動と火山活動によって特徴付けられ、大西中央海嶺の影響を受けているものの、その直接的な構造体とは異なる。パンゲア大陸の分裂とマントルとの関係は、地球内部の動力学と密接に関連している。
地球の内部にはマントルと呼ばれる層があり、これは固体であるにもかかわらず、地質学的スケールで、非常に長い時間を掛けて流動的に動くことができる。熱い物質が上昇し、冷えた物質が沈むという対流運動が行われている。この対流が、地表のプレートを引っ張り、押し合う力を生み出す。
マントル対流によって引き裂かれると、大陸地殻が薄くなり、リフトバレーrift valley(裂け目)が形成される。ここからマグマが上昇し、新しい海洋地殻を生成する。リフトバレーが拡大し続けることで、大陸が次第に分裂し、現在の大陸の配置に至った。マントルのエネルギーは膨大で、その熱エネルギー源のマントルは約1,200℃から4,000℃の温度に達することがあり、これが対流運動を引き起す。このマントル対流は数千kmに及ぶ巨大な対流セルを形成し、プレートを動かす力を提供した。その地殻変動は、火山活動や地震などはすべてマントルから伝わるエネルギーによって引き起こされる。マントルのエネルギーは、地球の表面変動の根本的な要因となり、地球全体の動的な特性を表現するための重要な要素となる。
この時代の火山活動は、地球全体に影響を与え、気候変動を加速させたと考えられている。海底のメタンハイドレートが温暖化によって融解し、大量のメタンガスが放出された。メタンCH4は温室効果ガスとして非常に強力で、二酸化炭素 CO2よりも猛威となる。
メタン分子は、二酸化炭素分子よりも効率的に赤外線を吸収し、地表から放出される熱を保持する能力も高い。これにより、メタンは単位あたりの温室効果はCO2よりも約25倍強力とされている。メタンは大気中に短期間(約10年)しか存続できないが、その間に強力な温室効果を発揮する。メタンは大気中で酸素分子O2や他の化合物と反応し、最終的には二酸化炭素CO2と水蒸気H2Oに変わる。
メタンCH4は反応性の高いヒドロキシルラジカル・OHと出会うと、酸化反応が起こる。最初にメチルラジカルCH3と水分子H2Oが生成される。
CH4+OH→CH3+H2O
その.メチルラジカルCH3がさらに酸素分子O2と反応し、メチルペルオキシラジカルCH3O2が生成される。
CH3+O2→CH3O2
その最終生成物のメチルペルオキシラジカルCH3O2が他の分子(例えばNOや他のラジカル)と反応し、
最終的にホルムアルデヒドHCHO、二酸化炭素CO2、および水蒸気H2Oが生成される。
この一連の反応により、メタンは大気中で酸化され、より持続性の低い物質に変換される。特に、ヒドロキシルラジカル•OHは「大気の洗浄剤」として重要な役割を果たしており、多くの有機化合物の分解を促進する。この反応メカニズムが大気中のメタン濃度を調節するのに寄与している。
その最終生成物のメチルラジカルCH3がさらに酸素分子O2と反応し、最終的に二酸化炭素CO2と水蒸気H2Oが形成される。
ヒドロキシルラジカル•OHは非常に反応性が高く、大気中で他の化合物と結合する前に短期間しか存在ない。そのため、単体の分子として浮遊することは非常に稀である。 •OHラジカルは、紫外線や太陽光のエネルギーが水蒸気H2Oを分解し、•OHラジカルを生成する。
H2O + hν → •OH + H
この•OHラジカルはすぐに周囲の化合物(特に有機化合物やメタン)と反応して分解を引き起こす。このようにして•OHラジカルは大気中の多くの汚染物質を分解し、「大気の洗浄剤」として機能している。大気中での短命な存在にもかかわらず、•OHラジカルは地球の化学バランスを保つ重要な役割を果たしている。 メタンは赤外線スペクトルの領域であれば強力に吸収するため、大気中の放射バランスにおいて大きな影響を与える。特に、水蒸気や他のガスが吸収しない波長域でメタンは効果的にエネルギーを吸収する。一方、二酸化炭素は大気中に数百年滞留するが、個々の分子の温室効果はそれほど高くない。
始新世Eocene(約5,380万年前~3,390万年前)の最初にはシベリアに大隕石が激突し、それが引き起こした気候変動により、新哺乳類に最初の危機が訪れた。始新世末、或いは次の漸新世初期には一時的に気温が急激に低下したが、この頃彗星が頻繁に地球に衝突したためだとする説もある(始新世終末事件)。
当時、大規模な海退が起こり、海の面積が減少したのは、気温低下が原因であるとも言われている。インド大陸がユーラシア大陸に接近し始めてテチス海が狭まっていき、南極大陸が南米大陸やオーストラリア大陸から分離するなど、始新世は海洋と大陸の配置が大きく変わりつつあった時代だが、それに伴って地球規模で循環する海流の動きも大きく変動していたと思われ、これもまた、海退と寒冷化の一因とされる。
気温が低下すると、海洋や陸上生態系が大気中の二酸化炭素CO2をより多く吸収するようになる。特に海洋では、気体の溶解度は温度に反比例するという原理が働き、温度が低いほど、水中に溶け込む二酸化炭素CO2の量が増える。これにより、冷たい海水はより多くのCO2を吸収するこれにより、大気中のCO2濃度が低下する。
CO2が海水H2Oに溶けると、以下のような一連の化学反応が進行する。
CO2 + H2O ⇌ H2CO3(炭酸)
H2CO3 ⇌ H+ (水素イオン) + HCO3-(重炭酸イオン)
HCO3- ⇌ H+ + CO3- (炭酸イオン)
これらの反応は、温度が低いと右側に進行しやすくなる。つまり、CO2が水中でより多くの炭酸や重炭酸イオンに変換されるため、海洋がCO2を吸収しやすくなる。
始新世-漸新世の移行期、動植物は多様化、特に植物の種類が増えた
プレートテクトニクスもまた、始新世の気候に影響を与えた。大陸プレートの移動や衝突、海洋プレートの沈み込みなどの地質活動は、地球の気候システムに大きな変化をもたらした。例えば、南極大陸が移動することで海流や風のパターンが変わり、それが気候に影響を与えた。
始新世から漸新世への移行期には、アルプス山脈が形成された。アフリカプレートがユーラシアプレートに押し付けられたことで、かつてのテチス海(古代の海洋)の海底地殻が褶曲し、高い山脈が形成された。既に約6,500万年前の新生代暁新世の初期、アフリカプレートとユーラシアプレートが衝突し始めていた。この衝突は、アルプス山脈の基盤となる岩石が形成された最初のステップであった。海底地殻の押し上げテチス海があったことを示す証拠として、ヒマラヤ山脈には海底の堆積物と思われる地層が激しく褶曲した露頭があり、その地層からは多数のアンモナイトの化石が出土している。
始新世は、地球の気候が非常に温暖であった時代であった。この時期、地球全体が温暖で、極地にも温暖な気候が広がっていた。この温暖な気候は、主に大気中の二酸化炭素CO₂濃度が高かったことによるもので、当時のCO₂濃度は現在の約3倍とされている。また、大陸の配置や海流のパターンも気候に影響を与えていた。
始新世の動植物は多様で、特に植物の種類が増えた。この時代には、現代の植物の多くの祖先が出現した。例えば、花と果実の被子植物が大いに発展し、現在の森林の基盤となる樹木が広がりった。また、初期の哺乳類もこの時代に出現し多様化した。
動物の繁殖状況については、温暖な気候が生態系に豊かな環境を提供し、多くの動物が繁殖しやすい環境を享受していた。特に、初期の哺乳類や爬虫類、鳥類が繁栄し、多様な種類が見られた。
具体的には、始新世の初期には気温が現在よりもはるかに高く、熱帯雨林が広範囲に広がっていた。この温暖な気候は、始新世の終わりにかけて徐々に冷却され、後の新生代の氷期に向かっていく過程にあった。
漸新世は、熱帯始新世の古風な世界と中新世のより近代的な生態系との間の重要な移行期と見なされることがよくあるが、漸新世の主な変化には、草原の世界的な拡大と、熱帯広葉樹林の赤道帯への回帰にある。漸新世の始まりは、始新世-漸新世の移行Eocene-Oligocene transition (EOT)と呼ばれている。約3,390万年前〜3,340万年に発生した絶滅イベント(動植物の入れ替わり)の時代でもあった。
始新世-漸新世の移行期は、始新世末期の約,3400万年前に起こった大規模な環境変化の時期を指す。この時期には、地球全体で急激な冷却が起こり、それに伴って多くの動植物が絶滅した。この現象は「始新世-漸新世の絶滅イベントEocene-Oligocene extinction event」として知られている。地球の気温が急激に下がり、概ね温暖な始新世の気候から寒冷な漸新世の気候へと移行した。これは南極大陸に氷床が形成され始めたことが大きな要因であった。しかし、既に始新世中期から後期にかけて大気中の二酸化炭素がゆっくりと減少していた。北アメリカ中部では、この40万年の間に年平均気温が約8.2±3.1℃も低下した。中央チベットでは氷点下に近い状況が存在していた。この厳しい地球の寒冷化は、南極大陸の恒久的な氷床被覆の始まりでもあった。
極端な気候の変化は、海洋の循環パターンも変化させた。これにより、海洋生物の生息環境が大きく変化した。特に温暖な気候に依存していた多くの植物や動物が絶滅した。一方で、寒冷な気候に適応した新しい種が出現した。この時期の変化は、地層や化石の記録からも 地質学的な証拠として明らかになっており、特に海洋の底に堆積した珪藻土や、有孔虫などの微化石の分布からも確認されている。この移行期は、かつての地球の生態系や気候環境に大きな影響を与え、結果的に現代の地球の姿を形成する上で非常に重要な役割を果たした。
(珪藻は植物性プランクトン、その化石を原料としているのが珪藻土で、その主成分は二酸化珪素SiO2である。海洋の底に堆積した珪藻土は、地質学や古気候学において非常に重要な情報を提供する。
珪藻土を採取し、顕微鏡で観察することで、微細な化石の形状や構造を詳細に調べ、これにより、過去の海洋生物の種類やその変遷を明らかにする【顕微鏡観察】。
化石や有機物中の酸素同位体や炭素同位体を分析することで、過去の海水温度や炭素循環についての情報を得ることができ、それらのデータは、古気候学研究には重要となる【同位体分析】。
化石や有機物に含まれる元素や化合物の組成を分析し、これにより、過去の海洋環境や生物の生存環境についての詳細な情報が得られる【化学分析】。
古代のDNA(aDNA)を抽出して分析することで、過去の生物の遺伝情報や進化の過程を理解することができる【DNA分析】。
放射性同位元素を用いて、化石や堆積物の年代を測定し、特定の地層や化石の正確な年代を特定し、過去の出来事のタイムラインを構築できる【年代測定】。
これらの方法を組み合わせることで、過去の海洋生態系の構造や変遷について詳細な理解が得られる。)
現存するほとんどの哺乳類の種は、漸新世の終わりまでに出現した。原始的な三本指の馬・サイ・ラクダ・鹿・ペッカリー(外見的はイノシシに近く、イノシシ科よりは小さい)などが活動していた。犬・クマ・イタチ・アライグマなどの肉食動物は、旧世界の暁新世を支配していた原始的な 肉食性哺乳類クレオドントに取って代わり始めていた。リスのようなナッツや果物を食べる果樹が生息地で減少したため、低木果樹や草本を食べるげっ歯類やウサギが増え、大幅な多様化を遂げた。
バラ科やグミ科、スグリ科・ヤマモモ科など果実が食べられる落葉低木は、古代から地球上に存在していたと考えられている。特に、これらの植物の多くは中生代(約2億5,217万年前〜6,600万年前)や新生代(6,600万年前〜現在)にかけて進化してきた。 果実をつける植物の出現は、昆虫や他の動物との共進化の過程を辿り、果実を介して種子を拡散する役割を果たしてきた。バラ科の果実は、中生代ジュラ紀(約2億130万年前〜約1億4,500万年前)には存在していたとされている。この時代には、現代のバラ科の植物の祖先が出現し、その後進化を遂げ、新生代にはイチゴ・ビワ・リンゴ・スモモなど多様化し、現在のように多くの種が存在するようになった。 地質時代、特に新生代にかけて、バラ科の果実は小型哺乳類や鳥類にとって重要な食物源であった。多くの動物がこれらの果実を食べることで、バラ科の植物は種子散布の恩恵を受けていた。植物の進化と分布については、化石記録や分子系統学的研究から多くの情報が得られている。これにより、特定の科や属がどのように進化し、広がっていったのかが明らかにされている。
かつてユーラシアに存在していた霊長類は、アフリカと南アメリカに生息範囲が縮小されていた。 初期のシカ・キリン・ブタ・ウシは、ユーラシアの漸新世中期には現れている。最初のネコ科動物であるプロアイルルスProailurusは、約3,080万〜2,500万年前の漸新世にヨーロッパとアジアに生息していた絶滅したネコ科の属で、化石はモンゴル・ドイツ・スペインで発見されている。Proailurusは、最初の「真の」猫であり、猫の家族全体の祖先であると考えられている。
パレオマストドンPalaeomastodon(「古代のマストドンmastodon」;paleoは「古・旧・原始」の意)は、ゾウ目テング内の絶滅した属で、その化石は、約3,390万〜2,303万年前の新生代古第三紀漸新世の地層から出土している。ゾウやマストドンの祖先または近祖先と考えられ、ゾウ目の一員として、現在のエジプト・エチオピア・リビア・サウジアラビアの沼地や河川デルタ環境に生息していた。2016年の研究では、成獣の肩の高さは約2.2m、重量は2.5t以上と現代のゾウに似た大型の草食動物と推定された。
漸新世の時代、大陸は現在の位置に向かって漂流し続けた。南極大陸とオーストラリアおよび南アメリカとの間の海底に存在する深海の溝や谷などの深海チャネルが確立されると、南極大陸はより孤立した。オーストラリアはジュラ紀以来、南極大陸から非常にゆっくりと離れていったが、2つの大陸の間に海路が確立された正確な時期は不明のままである。しかし、漸新世初期の終わりまでに、2つの大陸の間に深い水路が存在していたという推定もある。南アメリカと南極大陸の間のドレーク海峡の形成時期も不確かであり、推定値は始新世前期から中新世(約2,303万年前~約533万年前)の範囲であるが、ドレーク海峡を通る海洋循環も漸新世前期の終わりまでに存在していたようだが、漸新世中期から後期(約2,900万年前〜2,300万年前)から中新世中期(約1,500万年前)までのドレーク海峡の一時的な収縮によって中断された可能性もある。
(ドレーク海峡Drake Passageは、南アメリカ・ホーン岬と南極半島の北側に連なるサウス・シェトランド諸島との間の海峡。南極海であるが、ギネスブックで世界一幅の広い海峡として認定されており、最狭部でも約650kmもある。また世界でも最も荒れる海域の一つ。)
暁新世(約6,600万年前~約5,380万年前)に始まった北東太平洋の海洋構造プレートの再編成は、漸新世の北米沈み込み帯にマレー断裂帯とメンドシーノ断裂帯が到着したことで最高潮に達した。マレー断裂帯Mariana Trenchは、北アメリカ大陸の西岸南カリフォルニアの北部沖合いからハワイに向って北太平洋を東西方向に走る海底の大断裂帯で、太平洋プレートが北アメリカプレートの下に沈み込むことで形成された。
メンドシーノの亀裂帯Mendocino Fracture Zoneは、カリフォルニア州メンドシーノ岬のすぐ沖合から西に少なくとも4,000 kmにわたって北西ハワイ海嶺付近まで伸びる左ずれの断層帯で、その断層帯に沿って、海嶺やその麓には舟状海盆troughが連なる。北アメリカプレートと太平洋プレートの沈み込み帯に位置する太平洋の海底に形成された大規模な断層で、この亀裂帯は、地殻の破壊と再構築を引き起こす大規模なプレートの動きによって地震活動が活発な地域となっている。
舟状海盆(しゅうじょうかいぼん)とは、細長い海底盆地で、深さが6,000mより浅いもの。トラフtroughとも。細長くないものは単に海盆と呼び、深さ6,000mを超えるものは海溝trenchと言う。
メンドシーノ岬は、アメリカ合衆国西部カリフォルニア州北部のハンボルト郡に属している。サンフランシスコから北に300kmの北部海岸に位置する。溝の水深は西部で深く、6,000mを越す。海嶺からは玄武岩の礫が多産し、海面上にでていた時代もあった。メンドシーノ岬付近は、アメリカ合衆国の大陸部分の中でも、非常に地震活動が活発な地域として知られている。例えば岬の西の沖合いで発生した1992年の4月25日から26日にかけて起きたメンドシーノ岬地震のモーメントマグニチュードは7.2に達した 。メンドシーノの亀裂帯の形成過程は、プレートテクトニクスによるもので、具体的には、太平洋プレートが北アメリカプレートの下に沈み込む際に生じる圧力と引っ張りの力が、地殻を破壊し、亀裂帯を拡大する原因となる。この断層は、地殻の破壊と再構築を繰り返すことで成長し、地震の発生源となる。
太平洋プレートが北アメリカプレートの下に、ほぼ西の方向へ潜り込むところに日本海溝がある。マリアナ海溝は、フィリピン海プレートと太平洋プレートの境界に位置し、太平洋プレートの沈み込みによって形成されたもので、世界で最も深い海溝の一つであり、その最深部は、チャレンジャー深淵Challenger Deepと呼ばれ、最大深度は約11,000 mに達する。その北東304kmにグアム島がある。
 カスケード山脈とロッキー山脈の形成には、北アメリカプレートと太平洋プレートの相互作用が関与している。カスケード山脈は、北アメリカプレートと太平洋プレートが衝突することで形成された火山帯であり、この沈み込み帯で、太平洋プレートが高温のマントルに押し込まれ、その結果、マグマが上昇し火山を形成した。一方、ロッキー山脈は、北アメリカプレートとテイデスプレート(北東太平洋プレート)が衝突することで形成された造山帯であるが、ロッキー山脈は、太平洋プレートと北アメリカプレートの衝突や圧縮によって、北アメリカプレートが他の小規模なプレートと衝突することで地殻が押し上げられて形成された造山帯である。ロッキー山脈は、中生代白亜紀の8,000万年から新生代古第三紀暁新世の5,500万年前のララミド造山運動Laramide
orogenyの間に形成され、そこでは多くのプレートが北アメリカプレートの下に滑り込み始めた。その沈み込みの角度が浅かったため、北アメリカ西部に広がる山々の帯が広がっていった。それ以来、さらなる地殻変動と氷河による浸食により、褶曲山脈のロッキー山脈は急峻.な山と谷で形作られた。その全長は約4,800kmを超え、北はカナダのブリティッシュコロンビア州最北部から、南はアメリカ合衆国ニューメキシコ州の州都サンタフェ周辺に広がっている。山脈の最高峰はアメリカのエルバート山(4,401m)で、カナダのカナディアン・ロッキーではロブソン山(3,954m)が最も高く、豊かな自然と険しい地形が共存する。この山脈は褶曲運動によって形成されたため、火成岩と変成岩が主成分とされ、特に白亜紀の1億年前から6,600万年にかけて比較的若い山地が形作られ、一方で古生代初期から存在するバージェス頁岩など、先カンブリア時代にかかる6億年以上前に形成された層も確認されている。最後の氷河期の終わりに、人間は山脈に住み始めた。鉱物や毛皮などの天然資源が山岳地帯の初期の経済的開発を推進したが、山脈自体は人口密度が高かったわけではない。1859年、コロラド州クリップルクリークCripple
Creekの近くで金の鉱脈が見つかると、以後はゴールドラッシュに沸き、地域経済の姿を完全に変えた。
カスケード山脈とロッキー山脈の形成には、北アメリカプレートと太平洋プレートの相互作用が関与している。カスケード山脈は、北アメリカプレートと太平洋プレートが衝突することで形成された火山帯であり、この沈み込み帯で、太平洋プレートが高温のマントルに押し込まれ、その結果、マグマが上昇し火山を形成した。一方、ロッキー山脈は、北アメリカプレートとテイデスプレート(北東太平洋プレート)が衝突することで形成された造山帯であるが、ロッキー山脈は、太平洋プレートと北アメリカプレートの衝突や圧縮によって、北アメリカプレートが他の小規模なプレートと衝突することで地殻が押し上げられて形成された造山帯である。ロッキー山脈は、中生代白亜紀の8,000万年から新生代古第三紀暁新世の5,500万年前のララミド造山運動Laramide
orogenyの間に形成され、そこでは多くのプレートが北アメリカプレートの下に滑り込み始めた。その沈み込みの角度が浅かったため、北アメリカ西部に広がる山々の帯が広がっていった。それ以来、さらなる地殻変動と氷河による浸食により、褶曲山脈のロッキー山脈は急峻.な山と谷で形作られた。その全長は約4,800kmを超え、北はカナダのブリティッシュコロンビア州最北部から、南はアメリカ合衆国ニューメキシコ州の州都サンタフェ周辺に広がっている。山脈の最高峰はアメリカのエルバート山(4,401m)で、カナダのカナディアン・ロッキーではロブソン山(3,954m)が最も高く、豊かな自然と険しい地形が共存する。この山脈は褶曲運動によって形成されたため、火成岩と変成岩が主成分とされ、特に白亜紀の1億年前から6,600万年にかけて比較的若い山地が形作られ、一方で古生代初期から存在するバージェス頁岩など、先カンブリア時代にかかる6億年以上前に形成された層も確認されている。最後の氷河期の終わりに、人間は山脈に住み始めた。鉱物や毛皮などの天然資源が山岳地帯の初期の経済的開発を推進したが、山脈自体は人口密度が高かったわけではない。1859年、コロラド州クリップルクリークCripple
Creekの近くで金の鉱脈が見つかると、以後はゴールドラッシュに沸き、地域経済の姿を完全に変えた。ララミド造山運動とコロラド州鉱化帯との関係
ララミド造山運動Laramide Orogenyは、地質学的な出来事の一つで、特に北アメリカ西部の山脈の形成に大きな影響を与えた。この造山運動は、中生代白亜紀の後期から新生代暁新世(約80〜55百万年前)に発生した。 ララミド造山運動は、プレートテクトニクスに関連し、具体的には、ファラロンプレートが北アメリカプレートの下に沈み込む過程で、圧縮応力がプレート境界から遠く内陸部にまで伝わり、地殻変動を引き起こした。この結果、ロッキー山脈を含む現在の北アメリカ西部の褶曲山脈が形成された。
ララミド造山運動は、北アメリカプレートの一画であるクラプレートKula Plateが北方に移動する際に、ファラロンプレートFarallon Plateと衝突し、その圧力によりロッキー山脈の地殻が押し上げられ褶曲山脈の地層が形成された。このプロセスは長い時間をかけて進行し、現在のロッキー山脈の地形を形作った。ファラロンプレートFarallon Plateのスラブが北アメリカプレートの下に滑り込んでいた北アメリカの西海岸沖の地殻変動を起因にする。ファラロンプレートはかつて存在したプレートととして、現在はほとんどが北アメリカプレートの下に沈み込んでいる。そのため、現在の地図にはファラロンプレートとして表示されていない。しかし、ファラロンプレートの一部が現在のプレートの一部として残っており、これが太平洋プレートの西側に位置するゴルダプレートやフアン・デ・フカプレートJuan de Fuca Plate、ココスプレートなどである。
位置的には、ファラロンプレートはかつて北アメリカプレートの西側に広がっていた。現在、その残存部分はアメリカ西海岸から南下し、メキシコの西側沖に至る地域に分布している。その沈み込みに関連する火成活動は、海洋地殻が平らなスラブとなって沈み込み、つまり浅い角度で沈み込んでいたことを示している。その結果、その下にある海洋リソスフェアの浅い沈み込みの原因の1つが、このプレートの収束速度【海洋プレートが大陸プレートの下に沈み込む速さを指す。通常、年間数cm単位で測定される】が速まったことによる可能性が高い。そのため沈み込みに関連する火成活動は、プレートの端の近くではなく、はるか東のコロラド州鉱化帯に沿って発生している。ファラロンプレートが北アメリカプレートの下に沈み込むことで、高温高圧の条件が生じ、これが鉱物の生成を促進した。このようなプレートの動きは、コロラド州鉱化帯の鉱床形成に重要な役割を果たした。このように、ファラロンプレートとコロラド州鉱化帯の関係は、プレートテクトニクスと鉱床地質学の観点から非常に興味深い。
金は反応性の極めて低い元素で化合物をあまり作らないため、ほとんどは自然金として産する。電気伝導性が高く、腐食しにくいなど耐久性にも優れている。展延性と光沢を併せ持ち、金属加工の材料に適している。ほとんどの自然金は熱水の作用によって石英の脈の中に形成され、閃亜鉛鉱・方鉛鉱・黄鉄鉱などの硫化鉱物を伴う事が多い。
マグマ(溶岩)は大陸の地下にあるマントルの一部が高温によって溶け出したもので、これが地表の近くまでに上昇してくると火山を形成する。火山の地下に存在するマグマは高温であるため周囲の地層に含まれる地下水を温めていき、そこから熱水の対流(水の循環)が生まれる。そこにマグマから分離した金の粒子が紛れ込んでいく。
マグマの科学的組成としてはケイ素Siという石英SiO2の成分がもっとも多く含まれており、やがて熱水にはこのケイ素と金の粒子が含まれていく。この熱水の中に含まれるケイ素は冷えて固まると「石英」という岩石の成分に変化するため、この石英の部分に金が入り込んでいくことになる。これにより、金の粒子は石英のかたまり、すなわち石英脈に含まれることになる。それが金の鉱床を形成する。
金が石英に濃縮されていることが多いのは、地下深部でマグマが冷却されると、周囲の岩石に含まれる水が加熱されて熱水が形成される。その熱水は、周囲の岩石から金を溶解する。この熱水は高温高圧の状態で移動し、金を運搬する。
熱水が冷却されると、石英が結晶化し始める。このプロセスは主に低温高圧の環境で進行する。石英はSiO2の結晶であり、冷却によって固体の結晶を形成する。石英の結晶が成長する際に、その内部や周囲に微小な隙間が生じ、その隙間に金が捕捉される。結晶化が進むにつれて、金はさらに石英の結晶内部や隙間に取り込まれ、最終的に石英脈内に金が濃縮される形で鉱床が形成される。このプロセスにより、石英脈中に金が効果的に濃縮され、多くの金鉱脈が形成される。石英の結晶内部に取り込まれた金は、肉眼では確認できないほど微細な粒子として存在することが多い。
特に熱水の移動経路や、温度・圧力の条件が鉱床の形成に重要な役割を果たす。石英脈中に金が濃縮される形で鉱床が形成されるためには、水が金を運搬し、熱や圧力の変化によって金が沈殿するプロセスが必要となる。コロラド州の鉱化帯などでは、このようなプロセスが繰り返されることで、豊富な金や他の鉱物資源が集中する地域が形成されてきた。鉱床の形成には長い時間がかかる。このような金の鉱床を多く含む地層(鉱脈)がいわゆる「金山」とよばれる場所として開発されることになる。
ファラロンプレートは、古代には北アメリカプレートの一部であり、太平洋プレートとの境界付近に位置していた。一方、コロラド州鉱化帯は、アメリカ合衆国のコロラド州を含む地域に広がる鉱床帯である。ファラロンプレートが北アメリカプレートと衝突することで、地殻変動が引き起こされ、これが鉱床の形成に作用した。このプロセスは、地質学的な時間スケールで見ると非常に長い期間と広域に及ぶ。具体的には、ファラロンプレートが北アメリカプレートの下に沈み込むことで、高温高圧の条件が生じ、これが鉱物の生成を促進する。ファラロンプレートが北アメリカプレートの下に沈み込むことで、高温高圧の条件が生じ、金や銅などを産出するにはメカニズムが働く。ファラロンプレートが北アメリカプレートの下に沈み込むと、その深部の高温高圧の条件下でプレートが溶け始める。このメルトmelt(溶融物質)が上昇し、地殻内に侵入する。
生成されたマグマは、上昇して冷却される過程で、金属鉱物を濃縮させる。このプロセスで、銅や金などの鉱床が形成されることがある。沈み込み帯では、メルトの生成だけでなく、熱水活動も活発になる。熱水は、地殻中の金属成分を溶解し、これらが冷却される際に鉱床を形成する。長期間にわたる地質活動により、鉱床が集中する。この過程は数百万年から数億年にわたって進行している。これらのプロセスにより、ファラロンプレートの沈み込みは、銅や金などの貴重な鉱物資源を生成する場となる。この現象は、プレートテクトニクスの一部として地球の資源分布に大きく寄与している。
マグマが冷却されると、金属元素が結晶化し、鉱物として形成される。この過程で、金属元素が集まり、濃縮される。金属元素が濃縮されたマグマが地殻に貫入し、地殻内でのプレートの移動やマグマ活動により、金属元素が集中して濃縮され鉱床が形成される。鉱床が地熱や化学反応により変成されることで、金属元素がさらに濃縮される。コロラド州の鉱化帯には、他にも有力な鉱物資源がある。例えば、銀・鉛・亜鉛・銅などの鉱物は、コロラド州の鉱化帯で重要な役割を果たしている。
鉱物の金属元素が熱的変成や化学変成、移動と再濃縮などにより再結晶する事例として、鉄鉱石は高温で加熱されることで鉄を取り出し、その過程で再結晶化する。また、銅鉱石も同様に高温で加熱されることで銅を取り出し、再結晶化することがある。他にも、ニッケル鉱石や亜鉛鉱石なども同様のプロセスを経て再結晶化する。これらの金属元素は、熱的変成や化学変成を経て再結晶化し、より純粋な形で取り出される。コロラド州鉱化帯の鉱床は、このような複雑な地質プロセスの結果として形成された。これらは、数百万年から数億年にわたる長い地質学的時間スケールで進行した。
新生代古第三紀始新世初期の約5,000万年前、古生代の後期から中生代の前期にかけて超大陸パンゲアPangeaを取り囲んでいた、唯一の広大な大洋パンサラッサPanthalassaの東部に広く分布していたファラロンプレートFarallon Plateから、バンクーバープレートが分離する。そのバンクーバープレートは北アメリカプレートに沈み込みを続ける。古第三紀漸新世の約3,000万年前、太平洋プレートが北アメリカプレートに到達し、バンクーバープレートはファラロンプレートと地理的にも離れると、バンクーバープレートはファン・デ・フカプレートJuan de Fuca Plateと名前を変える。
新生代新第三紀中新世の約1,000万年前から太平洋プレートとファン・デ・フカプレートの間に存在する発散境界(海嶺)の向きが、およそ20°傾く。これが「沈み込み帯subduction zone」という状態となる。「沈み込み帯」とは、地球のプレートテクトニクスにおいて、1つのプレート(通常は海洋プレート)がもう1つのプレート(通常は大陸プレート)の下に沈み込む領域のこと指す。このプロセスは「沈み込み帯」とも呼ばれ、いくつかの特徴的な現象を引き起こす。これにより、ファン・デ・フカプレートがすべて北アメリカプレートの下に完全に沈み込み、ファン・デ・フカプレートは消滅した。長い地質学的な時間スケールでは、「沈み込み帯」は山脈や海溝などの地形を大規模に形成したり変えたりする。例えば、北アメリカのロッキー山脈や日本列島の南部フォッサマグナ一帯の富士山・箱根山・天城山などの火山帯がその好例となる。
太平洋プレートが北アメリカプレートの下に深く沈み込む際に、沈み込むプレートが溶けてマグマが生成され、火山活動を引き起こすが、大陸の中央西部では火成活動は発生しない事象を、地質学者は、沈み込み帯の近くでの火山活動の欠如をマグマギャップと呼んだ。この特定のギャップは、沈み込んだスラブが比較的冷たい大陸のリソスフェアと接触していたためであり、高温のアセノスフェアに接触していなかったためにマグマギャップ状態になった能性が高い。沈み込みの角度が浅く、それが引き起こした抗力の結果の一つが、ロッキー山脈の祖先である広い山岳地帯であった。原始ロッキー山脈の一部は、後に拡張によって大規模に改築され、盆地と山脈の多い州となった。
テイデスプレートも、太平洋プレートの一部であり、北米プレートと接している。このプレートは、太平洋プレートが北米プレートの下に沈み込む場所で、これにより火山活動や地震が発生する。北アメリカ西部の地形が時計回りに回転した主な原因も、プレートテクトニクスの力によるもので、特に、太平洋プレートと北アメリカプレートの相互作用が、この地域の地殻変動を引き起こし、地形の回転や変形に寄与している。北米大陸は、北米プレートの上にあり、テイデスプレートとの境界付近では、地殻変動が頻繁に起こりる。この地域は、アメリカ西海岸やカナダ西部に位置し、地震帯として知られている。
カスケード山脈の南の火山活動を終わらせ、北アメリカ西部の多くの地形を時計回りに回転させた。そのロッキー山脈は最盛期を迎えると、新しい火山弧は、海岸から遠く離れた北アメリカ西部で確立され、メキシコ中央部からモゴヨン・ダティル火山田を通ってサンファン火山田に至り、さらにユタ州とネバダ州を通ってノーザンカスケード山脈に至った。これらの火山からの巨大な火山灰の堆積物は、ハイプレーンズのホワイトリバーグループとアリカリーグループを作成し、その優れた化石層を遺存させる。
サンアンドレアス断層San Andreas Faultは、アメリカ合衆国太平洋岸のカリフォルニア州南部から西部にかけて約1,200kmにわたって続く、巨大な右横ずれ変形断層である。断層全体に沿った平均すべり率は、年間20〜35 mmの範囲で、その断層の活動によって周辺地域は地震の多発地帯となっている。断層の西にある太平洋プレートは北西方向に動いており、東の北アメリカプレートは南西に向かって動いているが、プレートテクトニクスの影響下で比較的南東に動いている。滑りの割合は、カリフォルニア全体で年間平均約33〜37mmと言う。
目次へ
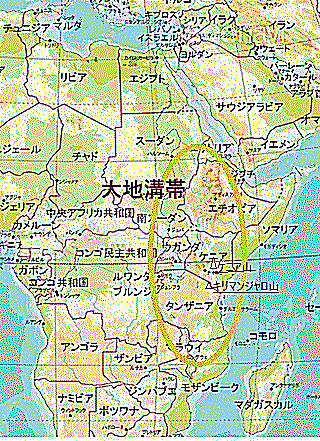
アフリカ大陸の地質は他の大陸の地質と異なることはない。5~6つの太古からの古い年代の岩石や地層の他に、原生代の岩石や地層も顕著にある。大地溝帯Great Rift Valleyは、アフリカ東部のアデン湾からモザンビークまで続く長大な谷で、中生代三畳紀の約2億5,000万年前から白亜紀の約1億年前にかけて形成され始めたと考えられている。この地溝帯はアフリカプレートとソマリアプレートの境界に位置し、現在も依然として東西に拡大している幅35〜100km、総延長は7,000kmにのぼる。正断層で地面が割れ、落差100mを超える急な崖や谷底にできた湖沼などが随所にある。
大地溝帯におけるプレートテクトニクスでは、アセノスフェア asthenosphere(岩流圏)とマグマが重要な役割を果たしている。アセノスフェアは、地殻の下にある硬い岩石の層で、固体ではあるものの、高温と高圧のために流動性を持っている。この層は、プレートが動く際の滑りやすい基盤となり、プレートの動きを可能にする。大地溝帯の形成には、このアセノスフェアの特性が大きく関与している。プレートの下のアセノスフェアには、岩石がすこし溶け滑りやすくなった薄い層がたくさん重なり、プレートが水平に動きやすくなっている。
これによりプレートが引き裂かれると、地殻が薄くなり、アセノスフェアが上昇する。この過程で、圧力が低下し、アセノスフェア内の部分的に溶けた岩石がマグマとして生成される。生成されたマグマは、密度が低いため上昇し、地殻内の割れ目や断層を通じて地表に向かう。この過程で火山活動が活発になり、溶岩流や火山が形成される。大地溝帯では、多くの火山が形成されており、これらの火山はマグマの供給によって活発に活動している。例えば、エチオピア高原やケニアの火山地帯がそれにあたる。アセノスフェアが柔らかく流動性を持っているため、プレートの動きが容易になる。プレートが引き裂かれる際には、アセノスフェアからのマグマが上昇し、火山活動を引き起こすという連鎖的なプロセスが見られまる。このため、大地溝帯のようなリフトゾーンでは、地殻の薄化とともに火山活動が顕著になる。
このように、アセノスフェアとマグマは大地溝帯の形成と進化において不可欠な要素であり、これにより、地質学的なダイナミクスが理解できる。
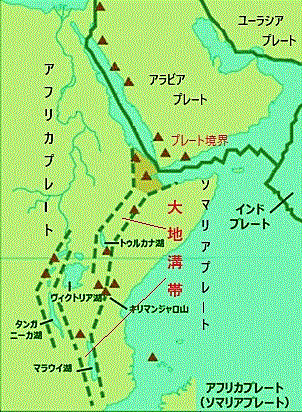 大地溝帯は、現在のケニア・エチオピア・タンザニア・ウガンダ・ルワンダ・ブルンジ・モザンビークなど、東アフリカの国々を含んでいる。この地域の地層は非常に複雑で、多様な環境を持っている。大地溝帯は断層活動によって形成された谷や湖があり、エチオピア高原、ケニア山、キリマンジャロ山などの火山も存在し、地層には、玄武岩や砂岩、石灰岩などの堆積岩が見られ、これらは数百万年にわたる地質学的な活動の結果がその表面を覆っている。
大地溝帯は、現在のケニア・エチオピア・タンザニア・ウガンダ・ルワンダ・ブルンジ・モザンビークなど、東アフリカの国々を含んでいる。この地域の地層は非常に複雑で、多様な環境を持っている。大地溝帯は断層活動によって形成された谷や湖があり、エチオピア高原、ケニア山、キリマンジャロ山などの火山も存在し、地層には、玄武岩や砂岩、石灰岩などの堆積岩が見られ、これらは数百万年にわたる地質学的な活動の結果がその表面を覆っている。この石灰岩は、大地溝帯が形成される前、その地域はかつて海に覆われていた時期があり、これらの水域に生息していた海洋生物の遺骸やカルシウム炭酸塩が沈殿して形成された。例えば、アフリカ大地溝帯の東部に位置するタンザニアのルクワ地域では、約2億5,000万年前の中生代三畳紀の魚竜(魚鰭類)「イクチオサウルス」などの化石が発見されている。石灰岩は堆積岩の一種であり、長い時間をかけて堆積物が圧縮されて固まることで形成される。プレートの動きや地殻変動により、これらの堆積岩が地表に現れることがある。現在の大地溝帯の環境は、サバンナや草原、森林そして湿地帯など多様な生態系が広がっている。これらの地域は、多くの動植物が生息するための豊かな生態系を保持し続けている。
(イクチオサウルスは、イルカのような流線型の魚竜で、数百にも及ぶ完全な骨格が発見されている事から、かなり研究も進んでいる。イクチオサウルスの体には大きな背ビレがあり、四肢はヒレ脚となり、前肢と後肢で大きさが異なり、前肢はかなり大きいが後肢は非常に短く小さい。尾ビレは垂直に立ち上がった半月系で、皮膚は滑らかで口の中には肉食の動物らしい小型の尖った歯が並んでいる。体長2m程度とあまり大きくはなく、同時代の海に生きたとされるリードシクティスやリオプレウロドンなどと比較するとかなり小さい。
イクチオサウルスは発達した大きな耳骨を持っていた事から、獲物となる小さな海生生物が起こす小さな振動を感知して狩りを行っていたと考えられている。また、発見された化石からイクチオサウルスは、卵を腹の中で孵化させる卵胎生である事なども分かっている。)
大地溝帯は、アフリカプレートとソマリアプレートの境界で形成された地形で、その境界の形成は、約3億年前の古生代最後の地質時代ペルム紀(約2億9900万年前~約2億5190万年前)に始まった。この境界は、アフリカプレートとソマリアプレートが分離し、それぞれ独自の移動を始めたことにより形成された。大地溝帯が活動して、アフリカプレートの本体(西側)とソマリアプレート(東側)が次第に分離して、プレートが引き離されることで、地表にひび割れが生じ、地溝帯が形成され、現在も盛んに陸地の拡大が起き現在も続いていることによる。このエネルギーの供給源は、地殻内の引張応力(テクトニックストレスplate tectonics)によるもので、 この過程で、地殻が薄くなり、断層や火山活動が活発化した。
ソマリアプレートは実は独立したプレートではなく、あくまでアフリカプレートの一部であり、大地溝帯が広がる境界の部分に亀裂が走り、1つだったアフリカプレートが2つに分裂しようとしている。 数十万~数百万年後には、アフリカプレートはこの亀裂を境界に2つのプレートに完全に分裂してしまうと考えられている。アフリカプレート内にはレユニオン島付近(マダガスカル島東方のインド洋上)、カナリア諸島付近(アフリカ大陸の北西沿岸に近い大西洋上)、カーボベルデ付近(北西アフリカの西沖合のマカロネシアに位置する)、カメルーンからサントメ・プリンシペなど多数のホットスポットが点在する。
ゾウの進化は非常に興味深いもので、数千万年に及ぶ進化の歴史があるが、それをどれだけ正確にたどれるかが人類史と同様極めて重要である。ゾウの祖先は、新生代古第三紀暁新世の約6,000万年前にアフリカで進化した。これらのゾウは、現代のゾウとは異なる特徴を持っていた。その約6,000万年前、ゾウの祖先にあたる種、例えばエリセリウムEritheriumは、ごく普通の短い鼻を持っていた。後にインド象やアフリカ象などのエリセリウム直系の子孫が、長い鼻を持つようになる。ゾウの遠い祖先は体もかなり小さかった(体重推定値3~8kg)。インド象やアフリカ象のよう立派な象牙を誇らしげに身に着けることもなかった。あの大きな幌のような耳を夏の暑い日にばたつかせることも、太古の昔にはできなかった。ゾウの化石は主にアフリカの東部と中央部で発見されている。特に、ケニア・タンザニア・ウガンダなどの国で多くの化石が見つかっている。ケニア・タンザニア・ウガンダはいずれもアフリカ大陸の東部に位置し、大地溝帯(グレート・リフト・バレーGreat Rift Valley)によって南北に貫かれ壮大な景観を形成している。この地溝帯は、アフリカプレートとソマリアプレートが分離する場所であるため、地球の内部活動が活発で、このプロセスは何百万年もかけて進行しており、地震や火山活動が頻繁に発生する。
この大地溝帯は地球の地殻が薄い場所の一つであり、地下のマグマが地表に近づくため、地熱エネルギー資源も豊富である。アフリカプレートとソマリアプレートが分かれる大地溝帯の活動が、アフリカ大陸東部に位置する東アフリカ大地溝帯を豊穣の台地にした。この地域は、アフリカプレートとソマリアプレートが分かれる場所であるため、地殻変動が活発である。この地域では、断層活動や火山活動が頻繁に発生しており、特にケニア山やソマリア山などの火山が活発である。また、大地溝帯は地下深くからのマグマの上昇によって形成されているため、地震も多発する。その一方、地殻変動は、地域の地形や気候に大きな影響を与える。例えば、大地溝帯の存在により、周辺地域に豊かな農業用地が広がる。大地溝帯には多くの火山があり、これらの火山が噴火すると火山灰が広範囲にわたって堆積する。火山灰が堆積した地域が肥沃な土壌を形成するのは、その化学組成と物理的特性による。火山灰は、カルシウム・マグネシウム・カリウム・リンなど生物にとって必須となるミネラルを豊富に含んでいる。これらの栄養素は植物の成長にも必須であり、火山灰が土壌に混ざることで、土壌の栄養バランスが改善される。
また、火山灰は細かい粒子であるが多孔質である。これが土壌に混ざると、土壌の水はけが良くなり、通気性が向上する。結果として、植物の根が酸素を吸収しやすくなり健全に成長できる。火山灰の多孔質性は、土壌の水を保持する能力を高め乾燥しにくくする。植物は安定して水供給を受けられる。火山灰には酸を中和する性質があり、酸性土壌を中和してpHバランスを改善し、植物にとってより良い成長環境が整えられる。これらの要素が組み合わさって、火山灰が堆積した地域はしばしば非常に肥沃な農地として知られている。火山活動が続く地域では、新たな火山灰が持続的に供給されるため、土壌の肥沃度が維持される。
酸性雨の原因は、化石燃料の燃焼や火山活動などにより放出される二酸化硫黄SO2や窒素酸化物NOxで、これらのガスは、大気中で光化学反応などの化学変化を起こし、硫酸や硝酸となって降水に溶け込み、酸性雨になる。また大規模な森林火災も酸性雨の原因になる。森林火災が発生すると、大量の煙やガスが大気中に放出される。その中には、二酸化硫黄SO2や窒素酸化物NOxなどが含まれており、これらが大気中で化学反応を起こすことで、酸性の物質が生成される。物質の酸性とアルカリ性の度合いの指標として一般に水素イオン濃度指数(pH)が用いられており、酸性度が強いほどpHは低くなる。純水(中性)のpHは7で、降水には大気中の二酸化炭素が溶け込むため、人為起源の大気汚染物質が無かったとしてもpHは7よりも低くなる。大気中の二酸化炭素が十分溶け込んだ場合のpHが5.6であるため、pH5.6が酸性雨の一つの目安となるが、火山やアルカリ土壌など周辺の状況によって本来の降水のpHは変わる。
自然界では、例えば酸性雨には硫酸H2SO4や硝酸HNO3などの酸が含まれており、これらは大気中の二酸化硫黄SO2や窒素酸化物NOxが水分と反応して生成される。また、火山活動自体も酸性のガスを放出する。火山から噴出されるガスには、二酸化炭素CO2や二酸化硫黄SO2などが含まれており、これらが水H2Oと反応して炭酸H2CO3や硫酸H2SO4を生成することがある。これらの酸が火山灰と反応して中和される。火山灰が酸を中和する性質は、その成分に含まれる鉱物による。特に、火山灰に含まれるカルシウムCa・マグネシウムMg・ナトリウムNa・カリウムKなどの成分が酸を中和する役割を果たす。これらの成分は、酸と反応して塩や水を生成し、酸性度を低下させる。
具体的には、カルシウム Caは、酸と反応して炭酸カルシウムCaCO3やカルシウム塩を形成、マグネシウム Mgは、酸と反応して炭酸マグネシウムMgCO3やマグネシウム塩を形成、ナトリウム Naと カリウム Kは、酸と反応して塩(ナトリウム塩やカリウム塩)を形成する。これらの中和反応によって、火山灰は酸を中和し、土壌の酸性度を調整する役割を果たす。
(カルシウムCaが炭酸H2CO3と反応して炭酸カルシウムCaCO3になる化学反応式は以下の通り
Ca + H2CO3 ⇒ CaCO3 + H2
これによって、土壌中でカルシウムCaが炭酸H2CO3と反応して安定な炭酸カルシウムCaCO3を生成する。実際の土壌環境ではこの反応が自然に進行することが多い。
マグネシウム Mgが酸と反応して炭酸マグネシウム MgCO3や他のマグネシウム塩を形成する際の化学反応式は
マグネシウムと塩酸の反応では : Mg + 2HCl ⇒ MgCl2 + H2
この反応では、マグネシウムは塩酸HClと反応して塩化マグネシウムMgCl2と水素ガスH2を生成する。
マグネシウムと炭酸の反応では : Mg + H2CO3 ⇒ MgCO3 + H2
この反応では、マグネシウムは炭酸H2CO3と反応して炭酸マグネシウムMgCO3と水素ガスH₂を生成する。
いずれの場合も、マグネシウムは酸と反応して特定のマグネシウム塩を形成する。 )
 |
| 漸新世初期には大規模な海退が起こった。北アメリカとヨーロッパは大西洋の拡大により完全に分断されたが、逆に北アメリカとアジアのシベリアとはベーリング海峡付近でしばしば接続し、動物の往来があった。 北アメリカと南アメリカは、白亜紀以降、大アンチル諸島が陸橋となってつながっていたが、やがて北東に移動していく。 インドがアジアに衝突し、テチス海は急速に消滅しつつあった。アフリカ・南アメリカ・オーストラリア・南極の各大陸は海で隔てられ、孤立している。各々の大陸が海退するほどの寒冷化による気候変動が、多くの動物の絶滅の一因となった。 アルプス山脈とヒマラヤ山脈の造山運動が開始された。北アメリカ西部の造山運動は続いている。 パラセラテリウムは草食動物で、主に乾燥した草原やステップ地帯に生息していた。大きな鼻先を持ち、長い鼻で低い植物を食べることができた。社会性は低く、単独で生活していたと考えられている。 漸新世の気候条件は、非常に低い気温が特徴であった。この間、南極大陸とグリーンランドは氷に覆われていた。 大陸の分離によって、動物相には地域ごとの違いが見られるようになった。また、前の始新世に栄えた動物の多くが、始新世と漸新世の境界付近で絶滅し、それに変わる新しい種の発展が見られる。 哺乳類の進化、特に大型化が進んだ。史上最大級の陸生哺乳類とされるアジア産奇蹄目(サイ類)のパラケラテリウムParaceratheriumはその代表と言える。 パラセラテリウムは、絶滅したサイの一種で、漸新世初期から後期(3,400万年前〜2,300万年前)に生息していた最大の陸生哺乳類の1つで、最初の化石は現在のパキスタンで発見され、中国とバルカン半島の間のユーラシア全体で遺物が発見されている。この巨大な動物は、現代の象に匹敵するほどの大きさを持ち、体長約7.4m、肩高約4.8m、体重約20tに達したとされている。 パラセラテリウムは草食動物で、主に乾燥した草原やステップ地帯で活動し、大きな鼻先を持ち、長い鼻で低い植物を食べることができた。社会性は低く、単独で生活していたと考えられている。 ゾウや現生のサイなどの現代の大型哺乳類の生活様式と類似していたようだ。その大きさのために、捕食者が少なく、妊娠期間が長かったとみられる。 ゾウの仲間の長鼻目(ちょうびもく)は、アフリカで進化し、大きな体躯を持ったが、まだ他の大陸には進出していない。 霊長目ではオナガザル上科(じょうか)と分かれる形で類人猿(ヒト上科)が出現し大きな発展を遂げていた。 |
農産物では、コーヒー が ケニアやウガンダなど、茶 が ケニアやタンザニアなど、砂糖がケニアやウガンダなど、コメ がウガンダやケニアなど、バナナがウガンダやケニアなどで生産されている。
鉱物資源では ダイヤモンド がアンゴラやコンゴなど、ゴールドがウガンダやケニアなど、ニッケルがコンゴやザンビアなど、鉄鉱石がケニアやウガンダなど、タングステンがコンゴやザンビアなどで採掘されている。
この地域の国々は、大地溝帯によって自然の境界線が引かれており、それぞれの国が異なる地形や気候を持っている。その歴史や文化も大地溝帯に影響を受けており、多くの先住民族がこの地域に住んでいた。ケニアのキリマンジャロ山の標高は約5,895m、実際にはタンザニアに位置している山であるが、ケニアの国境付近にあり、ケニアからもその雄大な姿を眺めることがでる。ケニア山の標高は約5,199m、ケニア中央部に位置し、首都ナイロビからもアクセスしやすい。タンザニアにはアフリカ最大の湖であるビクトリア湖がある。
ウガンダは、アフリカ大地溝帯の西側に位置しており、東アフリカ大地溝帯Eastern Rift Valleyと西アフリカ大地溝帯Western Rift Valleyが交差する地点で、豊かな自然環境と多様な生態系を持っている。人類の進化においても重要な役割を果たしている。東アフリカ大地溝帯は、アフリカの紅海から南に向かって広がる大地溝帯で、ケニアやタンザニアを通り、ウガンダに達する。ウガンダの東部は、この東アフリカ大地溝帯の一部が含まれている。ウガンダを含む東アフリカの大地溝帯は、人類の進化における重要な化石の発見地として知られている。特に、この地域では初期のホミニン(人類の祖先ヒト属)の化石が多数見つかっており、人類の進化の過程を理解する上で重要な手がかりを提供してくれる。ウガンダの地質構造は、他の地域と比較して安定しているため、化石や古代遺跡が良好に保存されている。このため、多くの重要な遺跡が発掘されており、人類の歴史を紐解く上で貴重な資料を提供してくれる。
西アフリカ大地溝帯はウガンダの西側を通り、アルバート湖やエドワード湖、キブ湖を含むエリアであり、コンゴ民主共和国との国境に近い。そのため古代の動物たちが生息していた場所として有名である。
ゾウの祖先に関する多くの情報も大地溝帯の化石に基づいている。特に、歯や骨の化石は彼らの形態や進化の過程を理解するための重要な手がかりとなっている。その化石の記録から、どのようにして現代のゾウが進化したのかを追跡できる。その祖先は、現代のゾウとは形態や生態ともにかなり異なっている。最初のゾウの祖先も、森林に生息していたと考えられて、木の葉や樹木の小枝を食べて生活していた。ただ、現代のゾウよりも小さく、体長は約1mで、また、鼻の形も現代のゾウとは異なり、鼻の穴は顔の側面に位置していた。また、彼らは群れを作らず、単独で生活していたとされている。
ゾウの進化の過程で、彼らは徐々に大きくなり、鼻の穴が顔の前方に移動し、現代のゾウのような形態に進化した。また、食性も草を食べるようになり、群れを作るようになった。それは生息環境と進化の過程によるもので、初期のゾウの祖先は、主に森林や密林に生息していたため、食物として利用できるものは木の葉や枝が中心となり、草地よりも森林が多い地域に住んでいたので、自然にこれらの資源に依存するようになった。その進化の初期段階での彼らの歯の構造も、柔らかい葉や小枝を噛み砕くのに適していたため、草を食べるよりも木の葉や小枝を食べるほうが効率的であった。進化の過程で、彼らの体格や歯の構造が変化し、草原地帯に適応していく中で、徐々に草を主食とするようになった。
パレオマストドンPalaeomastodon(「古代のマストドンmastodon」;paleoは「古・旧・原始」の意)は、ゾウ目テング内の絶滅した属で、その化石は、約3,390万〜2,303万年前の新生代古第三紀漸新世の地層から出土している。通常、ゾウやマストドンの祖先または近祖先と考えられ、ゾウ目の一員として、現在のエジプト・エチオピア・リビア・サウジアラビアの沼地や河川デルタ環境に生息していた。2016年の研究では、成獣の肩の高さは約2.2m、重量は2.5t以上と現代のゾウに似た大型の草食動物と推定された。
上顎と下顎の両方に牙を持っていた。下顎の牙は前方に突き出ており、一般的に平らでスコップのようで、おそらく木の樹皮をこすり落とし、さまざまな植物を根こそぎにするために使用されたようだ。対照的に、鋭い上顎の牙は主に防御兵器として機能していた。歯は垂直に生えていた。
アメリカマストドンmammut americanumは新生代新第三紀鮮新世の約350万年前から更新世約1万5,000年前まで北アメリカに生息していた原始的な長鼻類であった。約3,390万〜2,303万年前の漸新世のアフルカの地層から化石が出土しているパレオマストドンから進化した仲間の1つで、このアメリカマストドンは、更新世にアメリカ大陸全体に分布したと考えられている。体長は2.5~3.0m、大きく長い牙を持つが草食であった。寒冷地で生息していたアメリカマストドンは、褐色の体毛に覆われていた。
絶滅種のアメリカマストドン35頭のミトコンドリアゲノムの解析から、アメリカマストドンが、更新世(約258万年前~1万1,700年前)の間氷期の温暖化に対応して北米の北方緯度域に向かって繰り返し移動していたことを示した論文が、2020年9月2日、Nature Communications に掲載された。
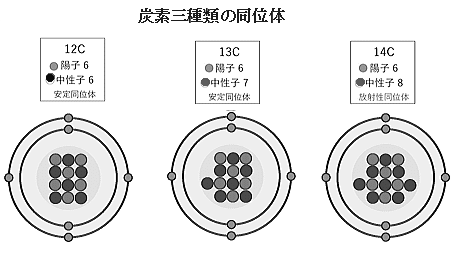 |
| 炭素には、3つの天然同位体がある。 炭素12は、 最も一般的な同位体で、天然の炭素の約99%を占めており、安定している。放射性はない。 炭素13 は、天然の炭素の約1%を占める安定同位体である。炭素12と同様に放射性はない。 炭素14 は、非常に微量で存在する放射性同位体で、半減期は約5,730年なので、放射年代測定に利用される。 炭素14は、大気中の二酸化炭素から植物に吸収され、動物は植物を食べることで体内に取り込む。炭素14は放射性物質であり、時間とともに放射線を放出して安定な炭素12に変わる。この放射線の放出速度は一定であり、その速度を利用して有機物を含む物体の年代を測定する。 測定方法は、年代を測定したい有機物のサンプルを採取する.。サンプルを化学処理して、炭素を取り出す。 取り出した炭素を加速器質量分析法(AMS)で分析し、炭素14の量を測定する.。測定された炭素14の量を基に、サンプルの年代を計算する。 放射性炭素年代測定法は、考古学・地質学・環境科学などさまざまな分野で使用されている。例えば、古代の木の標本や動物の骨、土壌サンプルなどの年代を測定する。 放射性炭素(14C)は、宇宙線と大気中の窒素との相互作用により、地球の大気中に生じる。その結果、14 C大気中の酸素と結合して放射性二酸化炭素を形成し、光合成によって植物に取り込まれる。 その後、動物は14 C植物を食べることによって取得する。動物や植物が死ぬと、環境との炭素交換が停止し、その後、14 Cは、含まれている14 C放射性崩壊を受け、歳月とともに減少し始める。その割合を測定する14 C木片や骨の破片など、死んだ植物や動物のサンプルから、動物や植物がいつ死んだかを計算するための情報を提供してくれる。 サンプルの年代が遡るほど、サンプルの数が少なくなり、また半減期があるため、炭素14では、5,730年ごとに、半分が失われる。 したがって、死んだ生物の炭素14の量を測定すると、それがどのくらい前にその大気との炭素の交換を停止したかを知ることができる。比較的手付かずの状況下では、放射性炭素研究所は5万年前まで、死んだ生物の放射性炭素の量を正確に測定することができる。 |
新生代更新世(氷河時代)ジェラシアン期の半ば過去80万年間の氷期と間氷期のサイクルのために、北米の居住可能な土地の約50%で氷床が周期的に拡大した。しかし、マストドンがこうした変動にどのように応答したのかは不明と言う。今回のEmil Karpinskiの研究チームは、北米の博物施設から入手したアメリカマストドンの骨と歯の化石の試料を調べて、33点の標本の完全なミトコンドリアゲノムの塩基配列を解読した。その結果、5つの異なる分類群のマストドンが特定され、そのうちの2種は、ベーリンジア(かつてロシアと米国の間に存在した地域)東部を起源としていた。Karpinskiの研究チームは、ベーリンジア東部に生息していたこれらの分類群の標本の年代に重複のないことを確認し、2つの分類群が別々の時期にベーリンジア東部に生息域を拡大させた可能性が高いと考えている。こうした拡大のあった時期は、温暖な気候条件によって森林や湿地が形成された間氷期と一致していた。
また、Karpinskiの研究チームは、この北方の分類群の遺伝的多様性のレベルが、大陸氷床の南側で生息していた分類群より低かったことを明らかにした上で、現代の気候変動によって、生物種の一部が、同じように生息域を北方に拡大させている可能性が高いと主張している。こうした生物種は、南方で生息する遺伝的多様性の高い集団と隔絶することで、脆弱な状態に陥ったようだ。アメリカマストドンは、更新世の終わりに、気候変動と狩猟の影響を受けて絶滅したと考えられている。
2021年9月24日
米ニューメキシコ州の米地質調査所United States Geological Survey(USGS)の研究チームはこのほど、北米大陸の内陸部で、2万3,000年前から2万1,000年前の人類の足跡を発見したと発表した。これまでは、人類の北米大陸への到達時期は1万6,000年前とされており、これより少なくとも7,000年も早くに到達していたこととなる。この研究成果は米科学誌「サイエンス」に掲載された。
研究チームによると、足跡はニューメキシコ州ホワイトサンズ国定公園のアルカリフラット(砂丘の上に広がる平坦な地形で、砂がアルカリ性であるため「アルカリフラット」と呼ばれている。)の一部となっている、浅い湖のほとりの柔らかい泥の中にあった。足跡が見つかった場所の上下の堆積層に含まれる種子の放射性炭素年代測定を実施し、足跡そのものができた年代を正確に特定することができた。足跡の大きさから、主に10代や幼い子供たちが往来した際にできたものと考えられている。科学者たちは、10代の子供たちが何をしていたのかははっきりわからないとしているが、ネイティブ・アメリカンの文化に見られる狩猟の一種のようなものを手伝っていた可能性がある。バッファロー・ジャンプと呼ばれる動物を崖に追い込む行動だったと考えている。オーストリア・ウィーン大学の放射性炭素年代測定の専門家トム・ハイアム教授は、「研究チームは足跡が見つかった場所の近くから採取した物質の年代を確認し、完全な陸生サンプル(木炭)が足跡の近くで採取された水生種の年代と同様の年代を示していることを確認した」と述べた。
 |
| 放射性炭素(14C)と窒素の関係は、放射性崩壊の過程に深く関連している。具体的には、炭素14が崩壊する際に窒素14に変わるという自然現象がある。大気中の窒素(窒素14)は、宇宙からの高エネルギー宇宙線の影響で、炭素14に変換される。 具体的には、宇宙線が窒素14原子に衝突すると中性子が窒素原子に取り込まれ、同時に陽子が放出される。これにより、窒素14が炭素14に変わる。炭素14は放射性同位体であり、時間とともに安定した窒素14に崩壊する。この崩壊過程をベータ崩壊と呼ぶ。 炭素14が崩壊する際、1つの中性子が陽子と電子に変わる。この結果、電子(ベータ粒子)が放出され、炭素14は窒素14に変わる。 放射性炭素年代測定法では、炭素14の崩壊率を測定することで有機物の年代を推定する。崩壊によって生成される窒素14は、最終的な生成物として自然界に存在する。再び宇宙線が窒素14原子に衝突し中性子が窒素原子に取り込まれ、同時に陽子が放出される。その大気中で生成された炭素14は、二酸化炭素として植物に取り込まれ、食物連鎖を通じて動物へと移行する。そして、時間が経過するとともに再び窒素14に戻る循環が繰り返される。 この関係性を利用して、放射性炭素年代測定法は、有機物の年代を測定する強力なツールとなる。考古学や地質学においては、炭素14の含有量を測定することで遺物や地層の年代を高精度で推定することができる。この方法は、約50,000年以内の年代を測定するのに適している。 宇宙線自体は中性子だけでなく、さまざまな高エネルギー粒子の流れから成り立っている。陽子(約90%)が宇宙線の主要成分で、アルファ粒子(約9%)、ヘリウムの原子核(2個の陽子と2個の中性子)、電子やガンマ線: 残りのわずかな割合を占め、また中性子もこれも含まれるが、全体の割合としては小さな部分を占める。 宇宙線が地球の大気に突入すると、大気中の原子核(主に窒素や酸素)と衝突して一連の二次宇宙線を生成する。これらの二次宇宙線には中性子が多く含まれている。これらの二次中性子が窒素14原子に衝突することで、窒素14が炭素14に変換される。具体的には、中性子が窒素14原子に取り込まれ、同時に陽子が放出されることで、窒素14が炭素14に変わりる。このプロセスは「n, p反応」と呼ばれる(nは中性子、pは陽子)。このようにして生成された炭素14は、地球の大気中に蓄積され、そこから植物や動物に取り込まれていく。このサイクルが放射性炭素年代測定法の基本原理となっている。 |
既に1980年代には、チリのモンテベルデで、1万4,500年前の人類の存在を示す確かな証拠が見つかっていた。2,000年代になると、米テキサス州中部の1万5,500年前のバターミルク・クリーク集落遺跡や、アイダホ州の1万6,000年前のクーパーズ・フェリー遺跡などでも広く認められるようになっていた。そして今回、ニューメキシコ州で発見された足跡は、最終氷期の最中に人類が北米大陸の内陸部に到達していたことを示している。英ケンブリッジ大学の遺伝学者、アンドレア・マニカ博士は、北米大陸の人口史に重要な意味を持つ発見だとした。専門外のため年代測定の信頼性についてはコメントできないとしつつ、「2万3,000年前に北米に人類がいたという確固たる証拠は、約1万5,000〜1万6,000年前にアジア人からネイティブ・アメリカンが分かれたことを示す遺伝学研究とは食い違っている」とBBCニュースで述べた。
「氷河期に陸続きになっていた際に押し寄せた新たな入植者が、北米大陸に先に定住していた最初の入植者に取って代わったことを示唆しているのだろう。どうやってそうなったのかは我々にはわからない」とも語った。
(放射性炭素Carbon-14は、大気中の窒素Nitrogen-14が宇宙線の影響を受けて生成される放射性同位体である。宇宙から地球に届く高エネルギーの粒子である宇宙線が、地球の大気に入る。 その宇宙線の粒子が大気中の窒素Nitrogen-14原子に衝突すると、高速の中性子が生成される。同時に陽子が放出される。窒素原子は炭素14(14C)に変わる。
この反応は
N-14 + n → C-14 + p nは中性子、pは陽子
このようにして生成された炭素-14は大気中に放出され、二酸化炭素CO2の形で植物や動物に取り込まれる。
この炭素-14は時間とともにベータ崩壊を起こし、窒素-14に戻る。
この崩壊過程を利用して、有機物の年代を測定するのが放射性炭素年代測定法である。
放射性炭素年代測定法Radiocarbon datingは、炭素14(14C)という放射性同位体の性質に基づいている。有機物の年代を測定するための方法で、放射性炭素Carbon-14の崩壊を利用して、物体がいつ作られたかを推定する。
具体的には、測定したい物体のサンプルを収集する。通常は、木材・動物の骨・土壌などが使われる。 そのサンプルから炭素14を抽出する。これには、化学的な処理が必要であり、その抽出された炭素から放射性炭素(Carbon-14)を検出する。このプロセスは、高感度の検出器を使用して行われる。 検出された放射性炭素の残量を基に、物体の年代を推定する。つまり放射性炭素は時間とともに崩壊するため、その割合を計算することで年代が求められまる。
炭素14 は、宇宙線と地球大気中の窒素との反応により常時生成される。生成された炭素14 は、大気中で酸素と結合して二酸化炭素となり、植物に吸収される。 動物は食物連鎖を通じてこの炭素14 を体内に取り込む。 それにより 、生物は生存中、大気中の炭素14 濃度とほぼ同じ割合の炭素14 を体内に保持す る。
それによる年代測定の原理 は、生物の死後、新たな炭素14 の取り込みが停止する。 以後、体内に存在する炭素14 は放射性崩壊により、時間とともに減少する。 炭素14 の半減期は約 5,730 年である。 この特性を利用し、試料中の炭素14 残存量を測定することで、生物の死亡時期を推定する。
放射性炭素年代測定法は、考古学や地質学などの分野で広く使用されている。)
放射性炭素年代測定の詳細
https://rarememory.com/carbonisotope/carbonisotope.html
目次へ
新生代新第三紀の中新世Mioceneの前の時代の新生代古第三紀最期の時代の漸新世Oligoceneは、約3,390万年前から約2,303万年前までの期間を指し、この古第三紀の最後の時代はルペリアン期Rupelian(約3,390万年前〜2,810万年前)とチャッティアン期(約2,810万年前〜2,303万年前)の2つに区分され、地球の環境と生物の進化においてそれぞれ特筆すべき時代であった。
 |
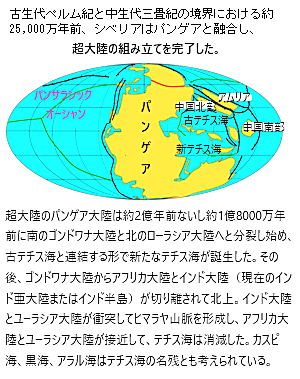
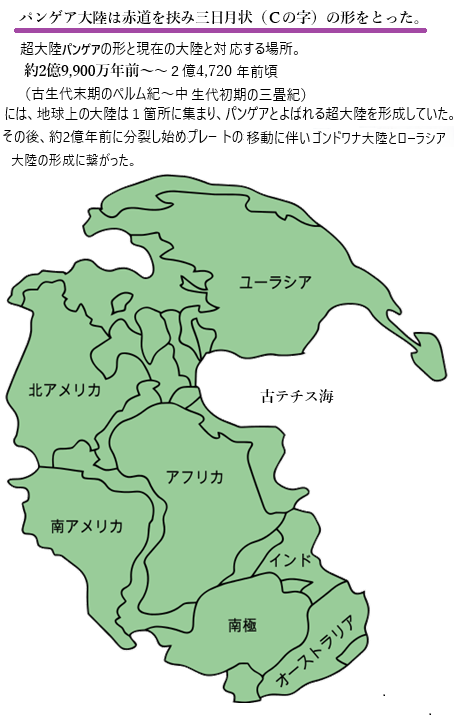
| ゴンドワナ大陸は、プロトゴンドワナと呼ばれるより古い超大陸の一部として誕生した。約5億4000万年前のカンブリア紀末期に、プロトゴンドワナが分裂し、ゴンドワナが形成された。この大陸は、地球のプレート運動によって形成され、その後の数億年にわたるプレート運動によって様々な地殻変動を経て成長した。 ゴンドワナ大陸は古生代カンブリオ紀の約5億4,000万年前から約1億8,000万年前まで存在した古代の超大陸で、現在の南半球の大陸,、南アメリカ・アフリカ・南極・オーストラリア・インド・アラビア半島などを含んでいた。 シルル紀中期の地球の古地理学、約4億3,000万年前。アバロニアとバルティカはローレンシアと融合してローシアを形成した。 石炭紀後期の約3億1,000万年前。ゴンドワナ大陸が移動し、ローシアと融合してパンゲアという超大陸を形成した。ゴンドワナは、パンゲアという巨大な超大陸の一部となり、パンゲアの形成によって、地球上のほとんどの陸地が一つに集まった。このような地質学的な動きは、地球の気候や生物の進化に大きな影響を与えた。 古生代ペルム紀と中生代三畳紀の境界における約25,000万年前、シベリアはパンゲアと融合し、超大陸の組み立てを完了した。 約1億8,000万年前、パンゲアは分裂を始め、ゴンドワナも次第に分裂した。この分裂過程でゴンドワナの一部が現在の南アメリカ・アフリカ・インド・オーストラリア・南極大陸として分かれ、現在の大陸配置が形成された。 ゴンドワナ大陸の歴史は地球の地質学的な動きと深く結びついており、その変遷は地球の気候や生物の進化にも大きな影響を与えた。 |
ルペリアン期は、約3,390万年前から約2,803万年前までの期間にあたる。これは漸新世Oligoceneの前期にあたる地質時代である。ルペリアン期には、大陸プレートが移動し続け、そのそれぞれの『エポックメイキング』ごとに、生物の棲息圏などの環境に常に大きな影響を与えて来た。特に、南極大陸の位置移動が気候変動に大きな影響を及ぼした。南極が現在の位置に南下するにつれて、周囲の海流パターンが変化し、これが乾燥と冷害の一因となった。
また、南極大陸の氷床が形成され始めると、これが地球の気温低下に直結した。氷床の形成は大気中の二酸化炭素濃度を低下させ、これがさらに乾燥した気候を促進し寒冷化を速めた。そのため、この時期には、地球の気候や生態系に多くの変化が見られた。大陸の移動に伴う気候変動、陸上と海洋相互の環境変化、生物の絶滅と進化など、様々な出来事が同時期に発生している。かつて繁栄を極めた多丘歯目も3,390万年前の漸新世前期ルペリアン期に絶滅した。しかも、北アメリカとヨーロッパが大西洋中央海嶺によって分断され拡大するプロセスが進行しており、気候も乾燥化していく傾向があった。また、海洋生態系でも大きな変化が起こり、新しい生物種が出現するなど、多様性が増していった時期でもあった。
南極大陸の氷床が形成され始め、これが地球の気温低下に繋がった。氷床の形成は大気中の二酸化炭素濃度を低下させ、これがさらに乾燥した気候を促進するという、このメカニズムは、地球の気候システムと炭素循環の相互作用によって説明される。南極大陸は過去には超大陸ゴンドワナ大陸の一部で、温暖な気候を呈していた時期もあった。
約2億年前にこの超大陸が分裂を始めた。具体的には南アメリカ・アフリカ・オーストラリア・インド亜大陸などのすべての大陸が南極から離れていった。南極が現在の位置に孤立したのは約4,000万~3,000万年前でほぼ同時期に南極大陸に氷床が形成され始めたと考えられている。また、南米と南極半島の間も約3,000万年前から開き始め、ドレーク海峡が形成され、南極大陸の周りがすべて大きな海に囲まれるようになる。これにより最終的に、南極大陸の周りをぐるぐる回る海流である南極環流Antarctic Circumpolar Currentが形成されることになる。その南極環流は、南極大陸の周りを取り囲むように、南半球の偏西風に駆動され東向きに流れる海流(環流)となった。地球上で最大の海流である。典型的な流速は20 cm/s、流幅が広く、厚さが3,000m以上であるため、流量としては100~150スベルドラップsverdrup(Sv, 毎秒 106 m3 ; 1スベルドラップは 106 m3/sと定義される。断面積 106 m2、水深 1,000 m、幅 1,000 mの海域を流速 1 m/s で通過する海流の流量は1スベルドラップとなる。)に達する。
およそ3,300万年前の新生代古第三紀漸新世、南極大陸と陸続きだった南アメリカ大陸とオーストラリア大陸が完全に南極大陸から分裂し、海が南極大陸周辺を取り囲み、地球の左回り自転の影響で生まれた南極環流は、赤道からの暖流を遮って南極大陸を急速に氷の大陸へと変えた。大まかな流れの中心は太平洋とインド洋では南緯50度付近、大西洋では南緯60度付近にある。南半球の偏西風によって主に駆動されており、南極環流の位置は偏西風の緯度と大体一致している。
海面から1,000m深程度までの海洋の流れを、長期間にわたって平均すると、海洋にはいくつかの大規模な水平方向の流れがある。地球規模でみると、南極大陸周囲に流量の大きい東向きの南極環流があり、各太平洋・大西洋・インド洋の亜熱帯域などに大規模な循環が存在する。例えば、北半球亜熱帯域では時計回りの北太平洋循環があり、南半球亜熱帯域では反時計回りの南太平洋海流South Pacific Currentと南インド洋海流South Indian Ocean Currentが、南極環流の一部として機能して循環している。このような海洋表層の循環は、主に海上を吹く風が海面に及ぼす力(風の応力)によって駆動されており、しかも地球の自転による効果によっても「西岸強化」と呼ばれる現象が見られる。
 北半球の亜熱帯循環を例にすると、日本列島付近に幅の狭い強い北向きの流れ西岸境界流があり、北太平洋では黒潮、その東側はゆっくりとした南向きの流れがあるカリフォルニア海流が流れる。北太平洋海域をより詳細に見ると、熱帯域には反時計回りの熱帯循環、南太平洋では亜熱帯域には時計回りの亜熱帯循環、とインド洋域の亜寒帯域に時計回りの亜寒帯循環が流れる。このような循環は、いくつかの海流により構成され、亜熱帯循環では、北赤道海流、黒潮および黒潮続流、カリフォルニア海流などの海流により、亜寒帯循環であれば親潮や亜寒帯海流などの海流により構成されている。
北半球の亜熱帯循環を例にすると、日本列島付近に幅の狭い強い北向きの流れ西岸境界流があり、北太平洋では黒潮、その東側はゆっくりとした南向きの流れがあるカリフォルニア海流が流れる。北太平洋海域をより詳細に見ると、熱帯域には反時計回りの熱帯循環、南太平洋では亜熱帯域には時計回りの亜熱帯循環、とインド洋域の亜寒帯域に時計回りの亜寒帯循環が流れる。このような循環は、いくつかの海流により構成され、亜熱帯循環では、北赤道海流、黒潮および黒潮続流、カリフォルニア海流などの海流により、亜寒帯循環であれば親潮や亜寒帯海流などの海流により構成されている。南極環流は、南極大陸を囲むように流れる世界最大の海流であり、南太平洋海流と南インド洋海流、そして南大西洋海流South Atlantic Currentは、南極環流の一部として機能している。 南極環流は、南極大陸の周りを一方向に流れることで、南太平洋海流と南インド洋海流を含む他の海流とも関連している。これにより、南極環流は世界中の海洋をつなげ、地球規模の海洋循環に寄与している。南極環流と南太平洋海流・南インド洋海流そして南大西洋海流との相互作用は、地球全体の海水の温度や塩分濃度の調整に寄与し、気候変動の影響を緩和する役割を果たしている。
(地球の海洋システムは、温度と塩分濃度の調整を通して気候変動の影響を緩和する複雑なメカニズムを持っている。このプロセスは主に「熱塩循環」と呼ばれる大規模な海洋循環によって支えられている。
海水の密度は温度と塩分濃度によって決まる。冷たい水や塩分濃度の高い水は密度が高く、温かい水や塩分濃度の低い水は密度が低い。例えば北大西洋や南極の周辺の高緯度地域では、海水は冷たく、塩分濃度も高くなるため、密度が高くなる。これにより、この冷たく重い水が沈み込み深層を流れていく。一方、赤道近くや他の暖かい海域では、海水は温かく、塩分濃度が低いため、密度が低くなり上昇する。
この沈み込みと上昇のプロセスが、地球規模での大規模な海洋循環を駆動する。これにより、海洋全体を通じて熱と塩分が運ばれ、地球全体の海水の温度と塩分濃度が調整される。熱塩循環によって、熱が赤道から極地へと運ばれる。これにより、赤道付近の過剰な熱が極地へと移動し、地球の温度バランスが保たれる。
海洋は大気中の二酸化炭素を吸収する能力があり、その二酸化炭素の溶解度は温度に反比例する。つまり、温度が低いほど二酸化炭素は海水に溶けやすくなる。冷たい海水はCO2をより多く吸収し、保持することができる。熱塩循環によってこの二酸化炭素が深海に運ばれ、長期間にわたり貯蔵される。これにより、大気中の二酸化炭素濃度が抑えられ、温暖化の進行が緩和される。南極環流は、この大規模な熱塩循環の一部として機能している。南極環流は、南極周辺の冷たい水を他の海洋へと運ぶことで、南半球全体の熱と塩分の分布に影響を与える。これにより、地球規模での気候システムにおいて重要な役割を果たしている。このメカニズムにより、海洋は地球の気候変動に対して重要な緩和効果を発揮している。
高緯度地域では冬季に海水が凍結し、氷が形成される。この過程で塩分は氷に取り込まれずに周囲の海水に残るため、海水の塩分濃度が上昇する。春や夏に氷が融解すると、その水は比較的淡水に近いため、塩分濃度は一時的に低下するが、全体としては冬季の塩分濃度上昇が上回る。高緯度地域では蒸発量が少ないものの、特定の条件下では蒸発が海水の塩分濃度を上げることがある。蒸発によって水分が失われると、残った海水の塩分濃度が上昇する。高緯度地域での降水は淡水を供給し、一時的に塩分濃度を低下させることがあるが、降水の影響は比較的小さい。
高緯度地域では冷たい海水が沈み込み、深層の暖かい海水が表層に上がってくることがある。このプロセスによって、塩分の高い水が表層に供給されることがある。潮流や風の影響によっても、塩分濃度が異なる地域からの水が混合され、高緯度地域の塩分濃度に影響を与えることがある。
これらの要因が組み合わさることで、高緯度地域の海水の塩分濃度が変動し、全体として上昇することが多い。このようなプロセスは、海洋循環や気候変動にも重要な影響を与えている。)
南極大陸周辺の南極環流が形成されて、これが南極大陸の冷却と氷床の拡大を促進した。南極環流は、南極大陸周囲の冷たい海水を循環させることで、氷床の成長を速めた。
アルベド効果は、地球の表面や大気が太陽光を反射する能力を示す。アルベド(反射率)が高い表面ほど多くの太陽光を反射し、アルベドが低い表面ほど多くの太陽光を吸収する。この効果は、地球全体の気候や温度に大きな影響を与える。氷床形成とアルベド効果の関係では、その氷床が形成されると、その白い地表面が太陽光を強く反射するため、地球の全体的なアルベドが増加し、この反射率の増加効果により、地表に吸収される太陽エネルギーが減少して気温が低下する。南極大陸や北極海の氷床、雪に覆われた山々などが好例となるが、砂漠のような明るい表面は高いアルベドを持ち、太陽光の大部分を反射するため、これらの表面は以外にも比較的涼しさが保たれる。
意外なのが、熱帯雨林や深い海洋、都市部のような暗い表面は低いアルベドを持ち、太陽光の大部分を吸収する。これにより、アスファルトの道路の表面は温度が上昇する。アルベド効果は、地球全体のエネルギーバランスに影響を与える。高アルベドの氷床や雪が広がると、より多くの太陽光が反射され、地球全体の温度が低下する。
逆に、これらの現象が減少すると、太陽光の吸収が増え、温暖化が進行する。この地球温暖化により、南極やグリーンランドの氷床が融解していく。これにより、アルベドが低下し、さらなる温暖化を引き起こす「フィードバックループ」が発生する。
一般的に、水面のアルベドは低く、約0.06から0.1程度である。これが、深い海洋や湖沼なると、水面は約90%から94%の太陽光を吸収し、残りの6%から10%を反射する。深い海洋の水面は多くの太陽光を吸収するため、海洋は地球の主要な熱エネルギーの貯蔵庫となる。この熱エネルギーは、海流や風を通じて、台風など地球的規模の猛威となる。現代の地球史レベルで考えれば、海洋の温暖化が極端に高まるため、水面のアルベドがさらに低下し、より多くの太陽光が吸収される。これにより、海洋温度が上昇し、やがて地球的な規模で気温が上昇する「フィードバックループ」が発生する。
気温が上昇すると、氷や雪が溶け、地表のアルベドが低下し、溶けて露出した海水や地面は太陽光をより吸収しやすくなる。その吸収された太陽光がさらに地表を温め、気温上昇を加速させる(アイス・アルベド・フィードバックIce-albedo feedback)。
北極圏の永久凍土が溶けると、その中に閉じ込められていたメタンガスが大気中に放出される。メタンは強力な温室効果ガスであり、二酸化炭素よりもはるかに影響は拡大する(メタン放出フィードバックmethane-emissions feedback)。
気温が上昇すると、大気中の水蒸気量が増加する。水蒸気は温室効果ガスの一種であり、二酸化炭素が2倍になると、二酸化炭素そのものの影響は1.2℃の上昇と考えられているが、水蒸気のフィードバックは1.9℃の上昇になると考えられ、地球の温度を一層上昇させる効果がある(水蒸気フィードバックsteam feedback)。
気温が上昇すると、乾燥が進み、森林火災の発生頻度が増加する。大気中のさらなる二酸化炭素濃度の上昇は森林火災を頻発させる。また、森林が減少することで二酸化炭素の吸収能力も低下する(森林火災フィードバックforest fires feedback)。
水面のアルベドは、地域的な気候に極端な影響を与える。例えば、現代の海洋近くの多くの大都市は、海洋の熱を吸収する効果により温暖な気候を保つことが多いが、北極海の氷が融解すると、氷の高いアルベド(約0.6から0.9)が失われ、低アルベドの水面が露出する。これにより、より多くの太陽光が吸収され、それが熱エネルギーに転換し猛威を振るう。
(水面のアルベドは、太陽光の入射角度によって変わる。太陽が高い位置にある直射光は、水面により多くの光が吸収される。一方、太陽が低い位置にあれば、その斜め光は、反射される光の割合が増える。)
目次へ
気温が低下すると、海洋や陸上生態系が大気中の二酸化炭素CO2をより多く吸収するようになる。特に海洋では、気体の溶解度は温度に反比例するという原理が働き、温度が低いほど、水中に溶け込む二酸化炭素CO2の量が増える。これにより、冷たい海水はより多くのCO2を吸収するこれにより、大気中のCO2濃度が低下する。
CO2が海水H2Oに溶けると、以下のような一連の化学反応が進行する。
CO2 + H2O ⇌ H2CO3(炭酸)
H2CO3 ⇌ H+ + HCO3-(重炭酸イオン)
HCO3- ⇌ H+ + CO32-(炭酸イオン)
これらの反応は、温度が低いと右側に進行しやすくなる。つまり、CO2が水中でより多くの炭酸や重炭酸イオンに変換されるため、海洋がCO2を吸収しやすくなる。
(海洋での気体の溶解度が温度に反比例する理由は、基本的にはガス分子の運動エネルギーと溶媒との相互作用に関係している。これには次のような原理が働く。
運動エネルギーの増加: 温度が上昇すると、ガス分子の運動エネルギーも増加する。これにより、ガス分子はより活発に運動し、液体から飛び出しやすくなる。そのため、溶解度が低下する。
溶媒-ガス相互作用の減少: 温度が上がると、溶媒分子もより活発に動く。これにより、ガス分子と溶媒分子との間の相互作用が弱まり、ガス分子が溶媒中に留まる確率が低くなる。
エンタルピー enthalpy(H)変化: 溶解過程において、ガスが溶媒に溶ける際にはエネルギーの変化が伴う。一般に、ガスが溶けるときのエンタルピー変化は吸熱的であり、高温ではこの吸熱過程が抑制されるため、溶解度が低下する。『エンタルピー』とは、定圧下で加えた熱のことで、その熱は仕事と内部エネルギーの上昇に使われるが、その両方を合わせた、言わば『定圧下で加えた熱量』 とここでは簡単に解す。
エンタルピーとは、化学系の内部エネルギーに圧力と体積の積を加えたもので、エネルギーの総量を示す。溶解過程では、エンタルピーの変化が重要な役割を果たす。 ガスが液体に溶けると、ガス分子が液体中の分子と相互作用を始める。この相互作用に伴うエネルギー変化がエンタルピー変化である。 ガスが液体中に溶ける際、ガス分子が液体の分子間空間に入り込むために、液体の分子間の結合が一部破壊される。この過程にはエネルギーが必要であり、その分エンタルピーが増加する。つまり、吸熱反応である。
同時に、ガス分子と液体分子が新たに相互作用を形成することで、エネルギーが放出される。これが放熱反応であり、これによりエンタルピーが減少する。 その実際の溶解過程では、吸熱と放熱の両方の過程が同時に起こる。ガスの溶解に伴う総エンタルピー変化(ΔH)は、これら二つのエネルギー変化の合計である。
Δ 𝐻 = 𝐻 吸熱 − 𝐻 放熱
多くの場合、ガスの溶解過程は全体として吸熱的であり、これは溶解度が温度に依存する理由の一つである。温度が上がると吸熱的な反応が進みやすくなり、結果として溶解度が低下する。 このエンタルピー変化の理解は、化学工学や環境科学、気象学など多くの分野で重要な役割を果たす。
これらの原理が組み合わさることで、海洋における気体の溶解度が温度に反比例する傾向が生じる。この現象は特に二酸化炭素や酸素の溶解度に顕著に見られ、海洋の生態系や気候変動に大きな影響を与えている。)
大気中のCO2濃度が低下すると、その低い二酸化炭素条件下で気孔が開くが、気孔が開けば蒸散が盛んになり水分が多く失われる。植物の光合成活動が一時的に減少するが、長期的には根系を発達させ、土壌からの水分と栄養の吸収効率が向上させる。この植物がより水分効率の良い方法で成長するようになると、この植物の生態系の変化が、乾燥した環境に対する適応能力を高める。
CAMはベンケイソウ型有機酸代謝Crassulacean Acid Metabolism (CAM;ベンケイソウ型有機酸代謝)の略で、CAM型光合成を行う植物をCAM植物と呼ぶ。ベンケイソウ型有機酸代謝(CAM)は、ベンケイソウやサボテン、アロエなどの多肉植物や乾燥地帯に生息する植物が採用する光合成の一形態である。CAM植物は、昼間は気孔を閉じて水分の蒸散を防ぎ、夜間に気孔を開いて二酸化炭素CO2を取り込み、その夜間に取り込んだCO2を、昼間なると光合成を行うための材料として使う。
CAMはベンケイソウ科植物に限らず多くの植物にみられる代謝であるが、ベンケイソウ科植物の葉の(Acid)酸含量が夜間に増し、昼間に消失する現象が、この科の植物に特有な代謝として認識・研究されたことに由来して慣用的に用いられている。
『アロエの味は夜に酸っぱく、昼は甘くなる』、 Grew( 1682)やde Saussure (1804)はこの酸蓄積にCO2が必要と述べている.。20世紀中頃までには、CAM植物は生物時計による日周変化の典型として興味がもたれていた(Bunning 1958)。
CAM植物はシダ植物3科5属、裸子植物1科1属(イチョウ科のイチョウ属)、単子葉植物8科98属、双子葉植物33科239属に分布し、ラン科での推定種数7,000種を含め約16,000種にみられる。この数は地球上の維管束植物の約6%に相当する。このように多数の植物にみられるCAMの進化は、環境条件によって誘発された適応によるものと考えられている。CAM植物はCAM型光合成と、葉や茎に水分を蓄える能力を備え、乾燥した環境に適応する多汁性という形態的特性を併せもつこで、砂漠などの水分ストレスが多発する環境下で生育する植物として進化した。従来、CAM植物のバイオマス生産力(動植物などの生物資源から得られる有機物)はきわめて低いとされてきたが、適切な農業的管理下ではC3植物をしのぎ、C4植物に次ぐ高い能力を示す種もある。
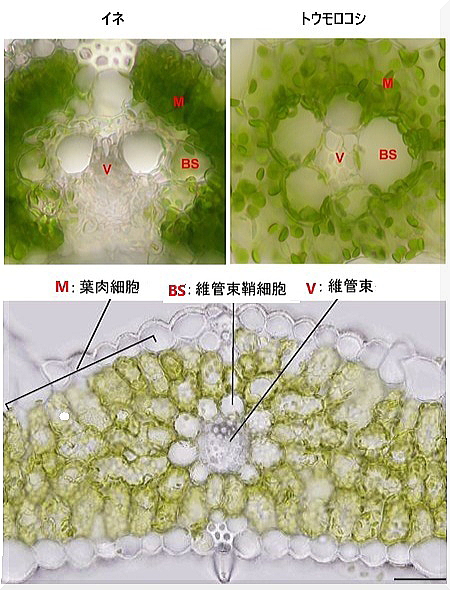 |
| ソルガムの原産地はアフリカとされている。数千年前にアフリカ大陸で最初に栽培され、その後世界中に広がった。 主な栽培国 アメリカ合衆国: 主に動物飼料やバイオエタノールの原料として栽培されている。 インド: 主に食用として栽培され、主食の一部となっている。 ナイジェリア: ソルガムの主要生産国で、食用や飼料として広く利用されている。 スーダン: ここでも重要な食料作物として栽培されている。 メキシコ: 食用および飼料作物として栽培されている。 ソルガムはC4植物で、カルビン回路に加えて、ハッチ・スラック経路Hatch-Slack Pathwayと呼ばれる特殊な経路を使用する。 ハッチ・スラック経路は、二段階の炭素固定プロセスを含み、特に高温や乾燥環境での光合成効率を高めるメカニズムである。 初期炭素固定では、光合成が行われる葉の葉肉細胞で、二酸化炭素CO2がホスホエノールピルビン酸(PEP)と結合してオキサロ酢酸(OAA)が生成される。 この反応は、PEPカルボキシラーゼという酵素によって触媒される。 :生成されたオキサロ酢酸は、リンゴ酸Malate、またはアスパラギン酸Aspartateに変換される。 これらの有機酸は、葉肉細胞から維管束鞘細胞へ移動する。 カルビン回路への供給は、維管束鞘細胞に到達したリンゴ酸やアスパラギン酸は、ピルビン酸PyruvateとCO2に分解される。 この分解反応により放出されたCO2が、カルビン回路で再利用されて炭素固定が行われる。 ピルビン酸は再び葉肉細胞に戻り、ATP(エネルギー)を使ってPEPに再生成される。 この循環が繰り返されることで、効率的なCO2固定が維持される。 |
その一方、葉組織には発達した 2 種の光合成細胞、葉肉細胞と維管束鞘細胞が存在し、両細胞にまたがるC4光合成回路を機能させている。その光合成を行う束鞘細胞は、葉の維管束組織を覆っている。つまりC4植物の光合成回路が機能しているは、葉緑体細胞と束鞘細胞の両方に存在している。しかも両細胞の葉緑体が環境ストレスに対して異なる応答を示す。C4 植物であるシコクビエ(米食が普及する以前には、日本各地で栽培されていた。日本では粉に挽いておねりや団子として食べられたことが多かった。)やトウモロコシが極強光下、もしくは乾燥・塩などの浸透圧ストレス条件下にあると、葉肉葉緑体が維管束鞘細胞側へ集合する。ま た、真夏に直射日光を受けている C4植物においても葉肉葉緑体の集合運動が起こる。それは過酷な環境下でC4 植物が生き抜くための耐性機構の一つではないかと考えられている。
ガス交換をする気孔は、乾燥や暑さによる水分の過剰な損失を抑えるため、一日のほとんどの時間帯で閉じている。これは、光呼吸に必要な酸素ではなく、RuBisCO酵素がCO2と反応する確率を高めるためである。
そのため熱帯や亜熱帯の気候に適応した植物が多い。 これらの特性により、C4植物はトウモロコシ・サトウキビ・ソルガムSorghum(イネ科の1年草)などの重要な農作物として広く栽培されている。トウモロコシの原産地はアメリカ大陸であるが、その栽培起源地域については諸説あり、米国南西部、メキシコ及び中央アメリカの複数地域説、メキシコと南米の複数地域説、メキ シコ南部単独説など複数がある。考古学的検証に基づくと、最初にトウモ ロコシの利用が始まったのは新生代第四紀完新世の約9,000~7,000 年前頃であり、約 5,400 年前頃には栽培が始まったと考えられている。アフリカ原産の「ソルガムきび」は、高温や乾燥に強く、約10,000年前頃から栽培化され始め、インド、中国を経てアジア各地、米国中南部など広範囲に広がっていた。
(『C3・CA・CAM植物 』の詳細 ⇒ https://rarememory.com/c3c4cam/c3c4cam.html)
氷床形成と大気循環の関係では、氷床の形成は、地球規模の大気循環パターンに影響を与えている。大気の冷却と乾燥化により、極地からの寒冷な風が増加し、乾燥した気候が広がる原因となる。これらのメカニズムが連鎖的に作用することで、氷床の形成が気温低下と乾燥した気候を促進する。地球の気候システムは非常に複雑で、多くの要因が相互に影響を与え合っている。
海洋酸性化は主に大気中の二酸化炭素CO2濃度の増加により引き起こされる。CO2が海水に溶けると炭酸H2CO3が形成され、これがさらに解離してH⁺が増加する。
海洋のH+(水素イオン)濃度が高まることを「海洋酸性化」と呼ぶ。貝類やサンゴを減少させる。カルシウムを利用して殻や骨を作る貝類・サンゴ・エビなどの生物は酸性下では生存できない。特に、生態系の基礎となるプランクトンの減少は食物連鎖全体に重大な影響をもたらす。小さなプランクトンから大型の海洋生物まで、酸性化はすべてのレベルで影響を及ぼす。この生物多様性の低下は、それに依存する他の種も影響を受け、生態系全体のバランスを崩す。
この海洋酸性化は海の化学組成を変え、生態系全体に長期的な変化をもたらす。
殻や骨は主に炭酸カルシウムCaCO3で構成されている。海洋の酸性化が進むと、水素イオンH+が増加し、炭酸カルシウムとの間で次のような化学反応を起こす。
CaCO3 + 2H+ → Ca2+ + CO2 + H2O
この反応で見られるように、炭酸カルシウムが溶解してカルシウムイオンCa2+と二酸化炭素CO2、水H2Oに分解される。
CO2が海水H2Oに溶けると、以下のような一連の化学反応が進行する。
CO2 + H2O ⇌ H2CO3 (炭酸)
H2CO3 ⇌ H+ + HCO3⁻ (重炭酸イオン)
HCO3⁻ ⇌ H+ + CO32- (炭酸イオン)
漸新世の地質時代のルペリアン期の後のチャッティアン期Chattian(約2,803万年前から約2,303万年前)では、いくつかの重要な変化が見られた。その名称はドイツ中部のカッセル周辺を指す古いラテン語名「Chatti」に由来する。ルペリアン期が全体的に温暖な気候であったが、チャッティアン期は地球規模の冷却が進み、より乾燥した気候が広がった。これにより、氷河の拡大が進み、地球の気温が下がった。そのため生物多様性が進み、新しい種類の海洋生物が出現した。特に、海洋のプランクトンや魚類の多様性が増した。また、Miogypsinoidesという新しい種類の有孔虫(石灰質の殻testと網状仮足を持つアメーバ様原生生物の一群である。仮足は真核細胞にみられる細胞質の一時的な突出である。これを備えた細胞は一般に「アメーバ様」「アメーバ状」と形容される。有孔虫は普通1mm以下の大きさ)が登場した。これらの生物は、浅海の堆積物に見られ、地域ごとの分布や進化の過程が研究されている。
陸上生物では、ハイエナに似た肉食性の哺乳類ハイエノドン類Hyaenodontaが、この時期に特に多様化した。絶滅した科に属し、ヨーロッパや北アメリカ、アジアに広く分布していた。例えば、Hyaenodon leptorhynchusという種がヨーロッパで発見されている大型の肉食動物で、非常に大きな頭蓋骨と長い吻部、体は長く、頑丈で、長い尾を持ち、小型の有蹄類を狩るために進化している。その歯は、他の肉食動物と異なり、骨を砕くよりも肉を切断するのに適していた。中新世の初期から中期にかけて、その時代の主要な捕食者であり、多様な生態系においても突出した存在であった。
チャッティアン期には、鳥類の進化も進み、これにより、生態系の複雑さが増した。鳥類の起源は約1億5000万年前のジュラ紀(約2億130万年前~約1億4500万年前)後期にまで遡り、恐竜の一群から進化した。直接の祖先は「獣脚類」と呼ばれる小型で二足歩行の肉食恐竜で、鋭い爪と歯、優れた運動能力を持ち、その中の一部の種が羽毛を持つようになった。近年、羽毛恐竜の化石が発見されたことによって、鳥類が恐竜の時代から現代に至るまで、進化をしながら姿を変えて生き延びてきたことが化石研究でわかってきた。これは生物が環境に適応し、生き残り、繁栄するためにどのように変化していったのかという貴重な事例となっている。
チャッティアン期には、古代の恐竜に似た鳥類から現代の鳥類へ進化する過渡期にあたる新しい種類の鳥が登場した。特に注目されるのは、エナンティオルニス類Enantiornithesというグループである。エナンティオルニス類は、チャッティアン期には世界中で優勢な鳥類グループに属していた。彼らは、古代の鳥類と現代の鳥類の間に位置し、その進化過程を理解する上で重要な役割を果たしている。
ネイバーノビスNavaornis hestiaeという新種の鳥が発見され、この発見は、古代の鳥類と現代の鳥類の進化の過渡期を理解するための「ロゼッタ・ストーン」とも呼ばれている。この鳥の頭蓋骨と脳の詳細な再構築が行われた。ネイバーノビスの脳は、アーケオプテリクスArchaeopteryx(始祖鳥)と現代の鳥類の間にあたり、その脳構造は古代的な特徴と現代的な特徴を合わせ持っていた。例えば、脳の一部はアーケオプテリクスに似ているが、他の部分は現代の鳥類に見られる特徴を持っていた。
最も有名な中間種が始祖鳥Archaeopteryxで、鳥類と恐竜の特徴を合わせ持つ化石として発見されており、鳥類と恐竜の中間的存在にあたる。そのため恐竜独特の特徴である長い尾骨と、鳥類には無い歯を持つ。鳥類の進化の過程を理解する上で重要な一歩となったネイバーノビスの頭蓋骨と脳の詳細な再構築により、鳥類の脳構造と行動の進化について新たな知見が得られた。
羽毛は鳥類の進化において最も顕著な特徴の一つであるが、初期の羽毛恐竜では飛行とは直接関係がなく、体温調節のために使用されていた可能性が高い。その後、時間が経つにつれて羽毛はより複雑な構造を持ち、やがて飛行に適した形に進化していった。現代の鳥類では、飛行・保温・水の防御、さらには異性を惹きつけるためにも使われている。
鳥類の骨格も、飛行能力に関連して独自の進化を遂げた。軽量で空気を入れられる骨を持ち、飛行の効率を高めるため構造を進化させた。尾骨を縮め、飛行中の機動性を高めるために適応している。これらの変化は、鳥類が優れた滑空力と持続力を獲得するためのものだった。
目次へ
 |
| パンゲア大陸に生息していた動物たちの中で、特に注目すべきグループは、爬虫類や初期の哺乳類、そして海生生物であった。初期の恐竜が現れた時期でもあり、彼らは植物や他の動物を食べることで生態系の一部を形成していた。テラトサウルス
Teratosaurusは三畳紀後期に生息したポポサウルス科に属する主竜類の化石爬虫類、このクルロタルシ類Crurotarsiは、主竜形類に属する爬虫類の一群、現生のワニに繋がる祖先群を含む。トカゲの様な姿の獣弓類や小型の草食恐竜を狩っていた。 シノグナトゥスCynognathusは四肢動物の一種で、哺乳類と爬虫類の特徴を併せ持っていた。全長1.5m程のオオカミ大の大型肉食動物、ただ脚が蹠行性(霊長類・クマ・パンダ・ウサギ・モグラなど踵を含む足の裏全体を使って歩行する)のため、体高はオオカミよりも低い。肉食に適化した鋭い歯を持つ、獰猛な捕食者であったと思われる。小さな切歯と発達した長い犬歯、断ち切ることに適した鋭い頬歯を持つ。 初期の魚類もこの時期に多様化し、さまざまなニッチを占めていた。硬い殻を持つ海生軟体動物アンモナイトが、プランクトンや小魚を捕食していた。 パンゲア大陸の分裂に伴い、気候が劇的に変化し、多くの動物が適応できず、地球史上最初の『大量絶滅時代』となった。大規模な火山活動が頻発し、気候変動や大気中の灰やガスの増加が多くの生物にとって致命的な環境を作り出した。特に海洋の酸素濃度が低下し、海洋生物の多くが絶滅した。 プランクトンなどの有機物が大量に生産され、これが海底に沈殿して分解される際に大量の酸素が消費され、深層水の酸素供給が減少した。 グルコースC6H12O6が酸素O2によって酸化される反応は、化学種が電子を失うプロセスを指す。具体的には、グルコースの炭素原子が電子を失い、二酸化炭素CO2と水H2Oとして放出される。これにより海洋無酸素事変Oceanic Anoxic Eventが勃発する。 |
大西洋中央海嶺は主に水中にあるが、その一部は、たとえばアイスランドのように、海抜に広がる程に十分な標高を持っている。その尾根の年間平均拡散率は約2.5cmと言う。大西洋での最深地点は、プエルトリコ海溝に位置し、8,605mである。
北大西洋の下の海嶺は、1853年にUSS Dolphin(ドルフィンはアメリカ海軍の潜水艦)の測深に基づいて、マシュー・フォンテーン・モーリーによって最初に推測された。尾根の存在と南大西洋へのその拡張は、1872年のイギリスの軍艦(海洋調査船)チャレンジャー号HMS Challengerの遠征中に確認された(HMSは国王【女王】陛下の船【His (Her) Majesty's Ship】の略)。隊長チャールズ・ワイヴィル・トムソンCharles Wyville Thomsonが率いる科学者のチームは、大西洋横断電信ケーブルの将来の位置を調査している間に、大西洋の中央部で大きな隆起を発見した。そして海嶺の存在は1925年にソナーによって確認され、ドイツのミーティア遠征隊によって南アフリカ共和国西ケープ州にあるアグラス岬周辺からインド洋まで広がっていることが解明された。
1950年代、マリー・サープ、ブルース・ヒーゼン、モーリス・ユーイングなどによる地球の海底の地図作成により、大西洋中央海嶺には奇妙な渓谷と尾根の海底地形があり、その中央の谷は地震学的に活動しており、多くの地震の震源地であることが明らかになった。ユーイング、ヒーゼン、サープは、この海嶺が地球上のすべての海の底にある40,000 kmの長さの本質的に連続した中央海嶺のシステムの一部であることを発見した。この全球的な海嶺システムの発見は、海底の広がりの理論と、アルフレッド・ウェゲナーの大陸移動とプレートテクトニクスの修正された形での膨張の理論の受け入れにつながった。この海嶺は、中生代ジュラ期の約1億8,000万年前に始まった超大陸パンゲアの分裂の中心であった。
大西洋中央海嶺には、海嶺の軸に沿ってほぼ全長にわたって走る深い地溝帯が含まれている。この裂け目は、隣接する構造プレート間の実際の境界を示しており、マントルからのマグマが海底に到達し、溶岩として噴火し、プレートの新しい地殻物質を生成する。
赤道付近の大西洋中央海嶺は、大西洋の最も深い場所の1つである最大深度7,758 mの狭い海底海溝であるロマンシュ海溝によって、北大西洋海嶺と南大西洋海嶺に分かれている。しかし、この海溝は、北アメリカプレートと南アメリカプレート、ユーラシアプレートとアフリカプレートの間の境界とは見なされていない。内容としては、大西洋中央海嶺は世界最大の海底山脈であり、その長さは約16,000kmにも及ぶ。
新生代古第三紀最期の時代の漸新世Oligoceneは、約3,390万年前から約2,303万年前までの期間を指し、この古第三紀の最後の時代はルペリアン期Rupelian(約3,390万年前〜2,810万年前)とチャッティアン期(約2,810万年前〜2,303万年前)の2つに区分され、地球の環境と生物の進化においてそれぞれ特筆すべき時代であった。
|
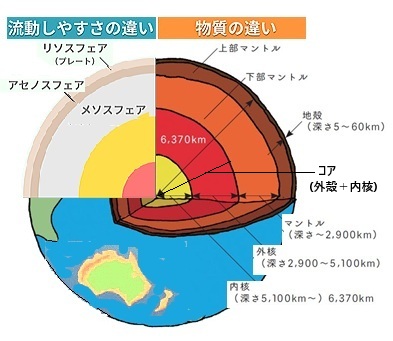
| 地球の内部構造は、リソスフェアLithosphere・アセノスフェアAsthenosphere・メソスフェアMesosphereの層ごとに異なり、それぞれの層が地球の動力学に重要な役割を果たしている。
リソスフェアは、地殻と上部マントルの一部から成る硬い外層で、厚さは約100 km程度、この層は、地殻プレートを形成し、地震や火山活動などの地質活動がここで起こる。
アセノスフェアの構造は、マントルの上部に位置する粘性のある層で、厚さは約100 km〜200 km、その特徴は、岩石が部分的に溶融していて、非常に柔らかく、上部のリソスフェアのプレートがこの層の上を動く。
メソスフェアの構造は、下部マントルに対応する層で、アセノスフェアの下に位置し、その厚さは約2,200 kmほどになり非常に硬く厚い層で、その特徴は高温高圧のため、地球内部の熱伝達が行われる。
リソスフェアはアセノスフェアの上を滑るように動き、これがプレートテクトニクス運動を引き起こし、地震や火山活動が発生させる。 メソスフェアは地球内部からの熱を伝え、対流を通じてアセノスフェアへと熱を移動させる。この熱の移動がプレート運動のエネルギー源となる。メソスフェアの成分は、主にシリケイト鉱物、例えば、カンラン石、輝石などから成っている。これらは高温高圧の環境下で結晶構造が変化し、密度が高くなる。その他の成分は、鉄やマグネシウムが豊富に含まれており、それによって鉱物の化学組成が形成されている。 |
大陸プレートの移動と海洋の拡大は、地球の気候にも影響を与えた。大陸の分離により、気温や降水量のパターンが変化し、新しい気候帯が形成された。これにより、新しい生態系が形成された。大西洋の拡大により、新しい海底が形成され、海洋の広がりが増加した。これにより、海洋の生態系や海流のパターンも変化した。比較的温暖で安定した気候が広がったが、やがて気候の冷却と乾燥に伴い、森林が縮小し、草原が拡大し植生が変化した。
漸新世の初期には、一時的に気温が低下し、気候が不安定になる現象が見受けられたが、このチャッティアン期の気候変動は、同時期の大海退や動物の大量絶滅と関連していると考えられている。北極の海氷や南極大陸の氷床がこの時代に形成されたという見解もある。
大西洋中央海嶺は「大西洋中央ライズ」と呼ばれる高まりの上に位置しており、これはアセノスフェアasthenosphere(ギリシャ語 astheneia;「弱さ、虚弱」)のマントル対流によって海洋地殻とリソスフェア(岩石圏)が押し上げられることによって形成されている。そのリソスフェアlithosphere(litho-は、「石、岩」)の下にある高温で岩石としては流れやすいマントルはアセノスフェアと呼ばれる。プレートとアセノスフェアの境界は、少なくとも温度のみで考える場合、明瞭なものではないが、温度が徐々に変わる境界がある。そのリソスフェアとアセノスフェアの境界を作る原因は部分融解あるいは水であるとする考えもあるが、大西洋のアセノスフェアのマントル対流は、地球内部のマントルが地表近くから深さ2,900kmまで広がる層で、プレートの下部を構成している。マントルは岩石でできた固体であり、液体ではないが、マントルの成分が地球中心部の熱で温められて上昇し、地表近くで冷えて下降するため、長い時間でみれば一種の対流運動を行っている。
マントル対流によって海洋地殻とリソスフェアが押し上げられることがある。このプロセスは、海洋地殻がマントルの上に沈み込むことで発生する。この沈み込む海洋地殻は、マントルの熱と圧力によって溶け、マントルの流動を促進する。この流動は、地殻の上にあるリソスフェアを押し上げ、火山活動や地震などの地質学的な現象を引き起こすことがある。
目次へ