| 八ヶ岳の特殊な生物生態 TOP 八ヶ岳創成 北八ヶ岳(冷山) 八ヶ岳岩石探訪 |  |
|
|
|
| 目次 |
|
| 1)八ヶ岳の特殊な生物生態 八ヶ岳の場合 地衣類 |
|
| 2)北八ヶ岳の地衣類 白駒池周辺の地衣類 天祥寺原のサルオガセ |
|
| 3)地衣類と苔の共存 地衣類の構造 |
|
| 4)蘚苔類 種子植物 シダ植物・コケ植物 藻類 蘚類と苔類 |
|
| 北八ヶ岳麦草峠付近の白駒の池湖畔の ドウダンツツジ・ナナカマド・ダケカンバなどが 赤や黄色に鮮やかに色付きます。 |
八ヶ岳では 亜高山帯と高山帯の境界を「森林限界」付近(約2,500m) に置かれることが多い。当然、風衝地・方位・地形条件によって境界は上下に変動する。八ヶ岳でも亜高山帯と高山帯の標高区分は重要な「目安」として示されているが、明確な線引きではなく、植生や地形条件などの環境によって緩やかに移行する。例えば南八ヶ岳の岩稜地では森林限界がやや低くなる傾向がある。
そのため「亜高山帯=1,600~2,500m」「高山帯=2,500m以上」という区分は便宜的なもので、実際には連続的な移行帯を形成している。
八ヶ岳の場合
亜高山帯は、およそ1,600m~2,500m、主な植生は、シラビソ-オオシラビソ林・コメツガ林などの樹林帯が広く分布し、林床が暗いため植物群落は発達しにくい。上部ではダケカンバ林が形成され、風衝地で、しかも雪の影響を受けやすい。
 |
| シラビソは、本州中部の亜高山帯(標高1,600~2,500m)に広く分布。八ヶ岳では低めの亜高山帯で優占。 常緑針葉樹で樹高20~25m。葉は短く枝に密着し、濃緑色で裏に白い気孔帯。 積雪には弱く、太平洋側の比較的雪の少ない山岳に多い。 密度の高い林冠を作るため、林床植物が発達しにくい。 |
 |
| オオシラビソは、本州中部~東北の亜高山帯。八ヶ岳の亜高山帯低部では、シラビソが優占。亜高山帯高部ではオオシラビソと混生する。樹高25~30m(最大40m)。球果がシラビソより大きく、若枝に赤褐色の短毛が密生。 多雪環境に強く、日本海側の山岳に優勢。東北地方では亜高山帯の代表種 シラビソとともに「縞枯れ現象」を起こす。 |
 コメツガは日本の固有種。本州の青森~紀伊半島・四国・九州の山岳地帯。 コメツガは日本の固有種。本州の青森~紀伊半島・四国・九州の山岳地帯。樹高20~30m。枝を横に広げ円錐形の樹形。葉は米粒のように小さく不揃い。土壌が未発達な場所でも生育可能。寿命が長く大木になる。八ヶ岳では、北八ヶ岳にコメツガ林が広く分布し、シラビソ・オオシラビソと混生する。 八ヶ岳には樹齢400年以上の巨樹があり、森の守り神のように崇められてきた。 |
| 縞枯れ現象は、亜高山帯のシラビソ・オオシラビソ優占林で顕著に見られる現象で、帯状に立ち枯れが進み、世代交代を繰り返すことで山肌に白い縞模様が現れる。 コメツガは、混交林の構成種として縞枯れに加わる。北八ヶ岳の縞枯山や茶臼山で顕著に観察でき、コメツガもその森の一員。 |
コマクサは、砂礫地の代表種で「高山植物の女王」と呼ばれ、南八ヶ岳の硫黄岳や横岳周辺に群落を形成する。
ウルップソウは、稀少種で、本州では八ヶ岳と北アルプス北部にしか分布せず、砂礫地に群落を作る。
キバナシャクナゲの群生地は国の天然記念物となり、硫黄岳や横岳周辺に大規模な群落を形成し、八ヶ岳を代表する高山植物と言える。シャクナゲの仲間では特に背が低く、高さ20~60cm程度。梅雨が終われば、本格的夏山シーズンになるが、その前、まだ山には登山者の姿がまばらなころ、高山の明るい稜線上ではキバナシャクナゲが咲きほこる。
梅雨時に咲く高山植物は多いが、多くの植物はその中の適した場所に生育しているが、キバナシャクナゲは風衝地の稜線の岩場や雪田、ハイマツの下などさまざまな環境の場所に適応する。ただ、その開花時期は風衝地が6月からと早いのに対して、雪田の個体は8月に咲くものもある。
主峰赤岳から硫黄岳に至る稜線上に咲くハクサンイチゲは、稜線の草地に群落を作り、白い花で彩る。黄緑色の柱頭を囲むように黄色い雄しべが密に取り囲んでいる。白くて花びらのように見える部分は、実は萼片である。それでは、花弁はどこにあるのかと図鑑を調べてみれば、花弁はないとの記載あり。
オヤマノエンドウは、紫色の花をつけ、ハクサンイチゲと混生して群落を形成するが、八ヶ岳の赤岳から横岳にかけての稜線の岩場に紫色の小さな花が、へばりつくように生えていた。
ミヤマダイコンソウは、八ヶ岳では、風当たりの強い岩場に生える多年草で、日当たりのよい尾根に、こんもりと点在する。高山の岩礫地や草地などに生える多年草で、高さは10~30cm、全体に黄褐色の毛が密生する。黄色い花に、丸く大きな葉の縁がギザギザなので、すぐこの花と分かる。
日本の固有種であるツクモグサは希少種で、北海道の利尻島、本州では八ヶ岳と白馬岳にのみ分布する。八ヶ岳でも前横岳から三叉峰周辺にのみ自生しており、他の峰では確認されていない。
岩場に群落を形成するキンポウゲ科オキナグサ属の多年草、草丈は5〜15cmと低く、岩場に張り付くように生育。その美しい花は高山植物の中でも特に早く咲くことで知られている。
花は淡いクリーム色で、全体が白や緑の産毛に覆われる。開花期は、6月上旬〜中旬、他の高山植物に先駆けて咲く。氷河期の残存植物として岩場に群落を形成する極めて希少な高山植物、絶滅危惧種に指定され、採取や接触は禁止、登山道から観察できるが、立入禁止区域には入らないことが求められている。
氷河期の残存植物(氷期遺存植物)とは、更新世の氷河期に広く分布していた植物が、氷期後の温暖化によって分布域を失い、現在では高山や寒冷地など限られた環境にのみ生き残っている種を指す。更新世の約258万年前~1万年前の氷河時代には、寒冷な気候が日本列島にも広がり、低地にも寒地性植物が分布していた。
氷期が終わり温暖化すると、これらの植物は低地から姿を消し、寒冷な環境が残る高山帯や北方地域における「希少種」として残存した。
地衣類
地衣類は「菌類(主に子嚢菌や担子菌)」が主体的に、藻類と共生して形成される複合体である。その共生の仕組みは、菌類が「住処」と「水分」を提供し、藻類が「光合成産物(有機栄養)」を供給する。この密接な共生関係によって「地衣体」という独特の構造が形成されている。国際植物命名規約でも、地衣類の学名は「構成する菌類」に与えられている。そのため、地衣類は植物ではなく、菌類に属する生物とされる。藻類(主に緑藻やシアノバクテリア)と共生しているため、見た目や生態が植物に似ている部分もあるが、植物は自ら光合成を行い無機物から有機物を合成する無機栄養autotrophyを作る。地衣類は光合成能力を持たない菌類が主体であるが、その菌類は共生する藻類が光合成によって合成した無機栄養を有機栄養として利用して生きている。形態的には他の生物の有機物を分解して栄養を得るコケ植物に似ているが、内部構造は全く異なる。
(有機栄養とは、他の生物が合成した有機物を摂取して生命活動に必要なエネルギーや物質を得る栄養様式のことで、これは「従属栄養【異栄養】」とも呼ばれる。植物が無機物から自ら有機物を合成する「無機栄養【独立栄養】」と対比される。)
地衣類は、菌類と藻類が共生関係を結んでできた複合体で、国際植物命名規約上は、その複合体を構成する共生菌に基づき地衣類を分類している。従って地衣類は、系統的に一つのまとまりを成す分類群ではなく、複数の系統から生じた、藻類との共生という生態的あるいは生理的な特徴を共有(=「地衣化」)する菌類の総称と言える。
一方、地衣類は、一般には蘚苔(センタイ)類などとともに「こけ」と認識されていることが多いが、「こけ」は「むし」などと同じく雑多な小さな生物群の総称であり、
専門用語では、「コケ植物」は学術的な総称であり、「蘚苔植物」は、蘚類+苔類を中心にした呼び方(ツノゴケ類も含める場合あり) で、「蘚苔類」も同義で使われることが多い。地衣類のことを「こけ」と呼んでも間違いではないが、コケ植物(あるいはコケ類)というと間違いになる。見た目はコケに似るが、分類学的にはコケ植物bryophyteとは全く異なる。
蘚苔植物という呼び方は、コケ植物全体を指す日本語表現のひとつである。蘚苔植物=コケ植物の別名であり、学術的にはコケ植物全体を指すため、蘚苔植物はコケ植物bryophyteに含まれるというより、同義語と考えるのが正確である。
コケ植物とは、維管束を持たず、胞子で繁殖し、半数体の配偶体世代が優勢となる陸上植物の一群で、蘚類・苔類・ツノゴケ類の3つに大別される。従って地衣類には、当然含まれない。
コケ植物の場合、配偶体が主体で、地面に広がる「苔」として目にする部分、つまり緑色で光合成を行い、葉や茎のような形を持つ。仮根で基質に付着し、水や養分を直接吸収する。精子器(雄器:ゆうき)や卵器(雌器:しき)を形成し、受精を行う。
胞子体は、従属的で、受精卵から発生し、配偶体の上で寄生的に育つ。光合成能力はほとんどなく、栄養は配偶体から受け取る。胞子嚢(カプセル)を形成し、胞子を散布する。大きさは小さく、配偶体の「付属物」のように見える。
例えばスギゴケであれば、緑の葉状体が「配偶体」=苔そのもの。その先端に細い柄とカプセル状の「胞子体」が乗っている。胞子体は茶色っぽく、光合成はほとんどせず、配偶体に養分を依存している。
つまり、普段「苔」と呼んでいる緑の部分は配偶体であり、コケ植物の生活の主体は配偶体世代である。コケ植物は陸上植物の初期進化を示す存在で、配偶体主体 → 胞子体主体への転換が、シダや種子植物への進化の大きなステップとなった。
かつて地衣類は、菌と藻の共生体とは考えられていなかったために、その学名は自ずと地衣類の体全体、つまり菌と藻の複合体に付けられていたことになる。しかし、後に菌と藻の共生体であることが明らかになったため、菌だけに対し新たな学名を与えようとした研究者が出てきて、多くの新しい学名を作り出している。また、複合体に対する学名とみなすのであれば、共生藻の種類が違うとき、学名を変えるべきであるかといった疑問も生じている。そこで、「地衣類の学名は、菌に対しつけられたものとみなす」という取り決めがなされ、これが現在の国際植物命名規約にも受け継がれている。
この考えに基づき、地衣類独自だった分類体系を、菌類の分類体系に組み込もうとする努力が20世紀中ごろから始められ、現在に至っている。生息する代表的な地衣類、サルオガセ類Usnea sppは、冷涼な亜高山帯針葉樹林の樹幹や枝に糸状に垂れ下がる典型的な地衣と言える。
サルオガセは大気汚染に弱いため、清浄な空気の指標となる地衣類であり、木に寄生せず、空気中の水分を吸収して生きるため、霧や湿度の高い森で繁茂する。八ヶ岳の苔の森や霧の多い山域が、まさに絶好の生育条件を満たしている。
八ヶ岳で特に多く見られるのは、北八ヶ岳の蓼科山や縞枯山、そして横岳周辺、サルオガセが登山口付近からすでに木々にぶら下がる姿が見られる。さらに白駒池周辺の原生林など、いずれも湿度が高く霧が多い冷涼な一帯に群生する。南八ヶ岳の赤岳や硫黄岳など岩稜が多く乾燥気味であるが、サルオガセは見られる。しかし、北八ヶ岳ほど豊富ではない。
目次へ
白駒池周辺の地衣類
 |
| 「天祥寺原」という地名は、かつてこの地に存在したとされる天祥寺という寺院跡に由来すると考えられている。ただ、寺院の創建年代・宗派・役割などを示す記録は残っておらず、廃寺となった経緯も不明と言う。 八ヶ岳周辺には修験道や山岳信仰に関わる寺院・坊跡が点在しており、天祥寺もその一環として存在した可能性がある。天祥寺原は蓼科山の南麓に広がる亜高山帯の高原で、修験者や参拝者が立ち寄るには適した平坦地であり、周囲はシラビソ林やダケカンバ林に囲まれ、湿原や草原が点在するため、生活や修行の場としても利用された可能性がある。 標高約1980m、蓼科山(2531m)の南麓に広がる高原地帯。北西に双子山(2223m)と双子池、北に北横岳(2480m)、東に大河原峠、南に竜源橋からの登山道が接続。 北八ヶ岳は「森の八ヶ岳」と呼ばれるほど森林が豊かで、天祥寺原もその典型的な亜高山帯植生を示す。 針葉樹林:シラビソ・オオシラビソ・コメツガが主体。トウヒやチョウセンマツが混じる場所もある。 広葉樹林は、ダケカンバ、ネコシデ、ナナカマドなどが点在。 低木・草本は、コヨウラクツツジ・カニコウモリ・シノザサ・シダ類。 亀甲池や双子池周辺には、湿原性植物が見られ、苔類が一面を覆う場所も多い。 季節の花:初夏にはレンゲツツジ、夏にはニッコウキスゲ、秋にはススキやアキノキリンソウなどが群落を作る。 蓼科山登山の「天祥寺原コース」として知られ、竜源橋から天祥寺原を経て将軍平に至るルートがある。周回ルートでは双子池や雨池を経て麦草峠へ抜けることも可能。 北八ヶ岳は「森の八ヶ岳」と呼ばれるほど森林が豊かで、天祥寺原もその典型的な亜高山帯植生を示す。 針葉樹林:シラビソ・オオシラビソ・コメツガが主体。トウヒやチョウセンマツが混じる場所もある。 広葉樹林:ダケカンバ、ネコシデ、ナナカマドなどが点在。 低木・草本:コヨウラクツツジ、カニコウモリ、シノザサ、シダ類。 湿原植物は、亀甲池や双子池周辺には湿原性植物が見られ、苔類が一面を覆う場所も多い。 初夏にはレンゲツツジ、夏にはニッコウキスゲ、秋にはススキやアキノキリンソウなどが群落を作る。 竜源橋は蓼科山南麓を流れる滝ノ湯川に架かる橋で、その上流域に位置している。周囲は亜高山帯の針葉樹林で、霧がかかりやすいため、サルオガセ(地衣類)が樹木に密集して垂れ下がる景観がよく見られる。竜源橋は蓼科山南麓を流れる滝ノ湯川に架かる橋で、その上流域に位置している。周囲は亜高山帯の針葉樹林で、霧がかかりやすいため、サルオガセ(地衣類)が樹木に密集して垂れ下がる景観がよく見られる。 |
エバーリア属は、灰緑色〜淡緑色の樹枝状地衣、樹皮に着生し、枝から糸状に垂れ下がる。香料原料としても知られる「オークモス」などを含む。
アレクトリア属も、細長い糸状の地衣で、黄緑色〜灰色。「髭状地衣」と呼ばれ、針葉樹林帯に多い。空気中の水分を吸収して生育する。
ブリオリア属は、暗褐色〜黒褐色の糸状地衣。サルオガセに似るが色が濃く、細い糸状で樹木から垂れ下がる。北方の冷涼な森林に広く分布している。
白駒池周辺では、サルオガセの白い糸状群落に加え、黒褐色のブリオリアや黄緑色のアレクトリアが混じり、森全体が「髭をたくわえた巨木」のように見える。
白駒池周辺で見られる上記の糸状地衣類以外に、苔の森の林床に広がるパルメリア属Parmeliaceaeやエバーリア属Everniaなどの葉状地衣類が、緑灰色や黄緑色を呈し、苔と混生して森の床を彩る。
また、ロブリア属Lobariaやグラファス属Graphisなど痂状地衣類(クラスト状Crustose)は、葉状や樹枝状のように立体的に広がらず、基物となる岩肌や樹皮の表面に強く固着して「かさぶた状」に薄く広がる形態を持つ。また溶岩地形や倒木の林床に葉状・痂状地衣類が苔と共存し、緑の絨毯を敷き詰める。
(痂状(「かじょう」または「かさぶたじょう」、Crustoseとは、藻類や地衣類が岩や土壌などの基質の上に平たくしっかり固着した状態を指す 。痂状は、裏面に下皮層がなく地衣体が直接、基質に密着、あるいは溶け込んでいるように見える。)
天祥寺原のサルオガセ
八ヶ岳火山群は古八ヶ岳期(約100万年前以前)と新八ヶ岳期(約50万年前以降)に大きく分けられる。白駒池溶岩は「新八ヶ岳期」の比較的若い火山活動による。白駒池周辺の更新世に噴出した安山岩質溶岩は、更新世中期のチバニアン期~後期の約28万~10万年前に形成されたと考えられている。この時期には、蓼科山や縞枯山、稲子岳などでも安山岩質溶岩が噴出し、現在の北八ヶ岳の「原状地形」を形成した。
滝の湯川の上流左岸付近の天祥寺原の原状地形は、蓼科山を含む北八ヶ岳火山群の噴火活動によって形成された溶岩流地形で、更新世に噴出した安山岩質溶岩(角閃石・紫蘇輝石・普通輝石を含む)が広がり、現在の緩やかな原状地形を形成した。溶岩流は厚さ数m規模で広がり、侵食を受けにくい塊状溶岩の特徴を残している。
この標高約1,980mに広がる高原状の大地は、苔むす針葉樹林と笹原が織りなす静謐な景観が特徴、植生はダケカンバやミズナラの広葉樹林からシラビソ・カラマツの針葉樹林、さらにハイマツ帯へと高度に応じて変化する。蓼科山や北横岳の麓に位置し、周囲には双子山・大岳などの山々が東方に連なる。登山道は笹原や林床の柔らかい土道が続き、歩きやすい区間が多い。
表面は風化・侵食を受け、苔や地衣類が繁茂する湿潤な環境となり、亜高山帯針葉樹林(シラビソ・コメツガ)に覆われている。周囲には亀甲池や双子池などの火山性凹地が点在し、池沼群とともに「森と池の八ヶ岳」と呼ばれる『苔むす森と静かな湿原的景観』を生み出している。ここは、北八ヶ岳特有の「深い原生林」から「静かな池沼群」へとつながるルートの中継地でもある。蓼科山や北横岳への登山ルートの分岐点となり、周囲の山々を見渡せる開けた場所もある。
周囲の池沼群、双子池・亀甲池などと同様に、湿潤な気候が地衣類や苔の繁茂を支えている。植生帯としては「苔と地衣類に彩られた亜高山針葉樹林」が核心的な特徴で、主体はシラビソやコメツガで、林床には苔類が厚く広がる。
サルオガセは、樹木に垂れ下がる糸状の地衣類で、湿潤で冷涼な環境を好むが、冬季にはサルオガセに氷晶が付着し、白銀のレースのような幻想的な姿を見せる。適度な積雪と寒さも、サルオガセの生育には好条件になっている。
トゲシバリ類Bryoria sppは、小鱗片状から樹枝状に発達し、直径5~20cmほどのマットを形成する。それはカラタチの枝に似た樹枝状に分枝する独特な構造を持ち、光環境に適応した生態と見られる。この地衣類の体の樹状部分の擬子柄(ぎしへい)は、多くは中空のストロー状で、繰り返し分枝している。表面には藻類層があり、内部は菌糸主体の髄層で、藻類層の腹面に穿孔(小さな孔)が開いている場合があり、内部の髄層と通じている。その樹枝状の立体構造によって、光を効率的に受けながら、雨や霧から水分を取り込む。この擬子柄を持つ地衣類は、乾燥尾根や湿地周辺など極端な環境にも適応している。
擬子柄の「藻類層 → 髄層」への無機栄養の移動は、地衣類の基本的な構造と機能に深く関わっている。そのバリアを兼ねた表皮層は、外界からの水分・無機塩類を取り込む吸収面でもあり、擬子柄の表面は微細な孔や粗い構造を持つため、毛細管現象で水分を保持しやすい。雨水・霧・露に溶け込んだ無機塩類(硝酸塩・アンモニウム・リン酸塩・カリウム・カルシウムなど)が、表皮層や藻類層を通じて吸収される。藻類層に光合成を行う緑藻や藍藻が生息し、有機物(糖類)を生成する。さらに下の 髄層は、菌糸が主体で、通気性が高く、水分や無機栄養の移動路となる。菌糸主体の髄層がそれを拡散・保持して全体に分配する仕組みになっている。
髄層は菌糸の網目構造でできており、拡散と毛細管輸送によって水分・無機塩類が内部へ移動する際には、 髄層は「スポンジ」のように働き、藻類層からの栄養を保持し、全体に分配する。全体への分配は、擬子柄の中空構造が、空気や水分の移動を助け、内部の菌糸が栄養を各部分へ拡散する。この仕組みにより、擬子柄全体が「小さな循環系」の機能を果たす。
トゲシバリ類は、サルオガセと混生することも多い。また、葉状地衣で、冷涼湿潤な環境に適応し岩や倒木に着生するハナゴケBryoria spp.と同様、樹枝状に枝分かれする地衣類と共生している藻類は緑藻であり、光があたる表側は青緑色である。裏側は灰色であり、穴がたくさんあいている。生育立地はハナゴケと同様に痩せた尾根筋の亜高山針葉樹林や湿地の周辺である。乾燥する尾根筋と湿地の周辺という、乾燥と過湿の両極端な場所に生育しているわけであるが、その共通点は十分な光が当たるということにある。
ウメノキゴケは樹幹や岩に着生する大型の葉状地衣、クロゴケ類は岩や樹皮に密着する暗色の地衣で、いずれも大気環境に敏感な「指標種」で、八ヶ岳では北八ヶ岳の白駒池周辺や縞枯山・北横岳などの亜高山帯針葉樹林でよく観察される。
クロゴケ類は「葉状地衣類」に分類される。岩や樹皮など安定した基物に付着し、光合成産物を利用して極めてゆっくり成長する生物で、年間の生長量は 3mm以下 と非常に遅く、長寿命で環境変化に敏感である。葉状地衣類のクロゴケ類は、紙状で表裏があり、裏面から伸びる「偽根」で基物に付着する。この葉状地衣類の内部構造は、上皮層・藻類層・髄層・下皮層の四層に分化しており、藻類層で光合成が行われる。
粉芽や裂芽などにより栄養繁殖を行い、菌糸と藻類が一体となった微細な構造が飛散して新しい個体を形成する。大気汚染や酸性雨に敏感で、環境指標生物として利用される。都市部では減少し、空気の清浄な山地に多く見られる。
北八ヶ岳の白駒池周辺は、苔と地衣類の宝庫で、白駒池畔の苔の森は「日本の貴重なコケの森」に選定され、ウメノキゴケやクロゴケ類も樹幹や岩上に見られる。縞枯山や北横岳では、亜高山帯針葉樹林にサルオガセなどの地衣類が豊富。クロゴケ類は岩場や樹皮に群落を形成する。天祥寺原や亀甲池周辺では、湿潤な環境でウメノキゴケが樹幹に着生している。
南八ヶ岳の赤岳や横岳の高山帯岩場にもクロゴケ類が見られるが、分布は北八ヶ岳ほど豊富ではない。
ウメノキゴケ類Hypogymnia spp.は比較的明るい樹幹や林床の倒木、岩に着生し広がる大型の葉状地衣、直径20cmに達する大形の葉状体。湿潤環境を好み、大気汚染に敏感。乾燥時は灰白色、湿潤時は緑色を帯びる。縁は波打ち、中央部に裂芽(れつが)を密生。梅の木やサクラなどの庭木にもよく着生する。
ウメノキゴケ類の「裂芽」とは、地衣体の表面に直接形成される小さな顆粒状や円筒状の突起で、菌糸と共生藻が一体となった栄養繁殖器官であり、これが親体から離れて新しい場所に定着すると、そのまま新しい個体へと成長する。裂芽は菌類の菌糸と藻類細胞がセットになった微細な塊で、親の地衣体から直接形成される。幹や岩の表面に広がるウメノキゴケの中央部をよく見ると、細かい粒状のざらつき=裂芽が確認できる。ウメノキゴケの場合、地衣体の背面中央に顆粒状~円筒状の裂芽が密生するのが特徴である。顕微鏡でなくても肉眼でざらつきとして見える場合がある。
ウメノキゴケは子器(胞子を作る器官)をつけることが稀で、主に裂芽によって繁殖する。裂芽はそのまま新しい個体になるため、都市部のように胞子が藻類を見つけにくい環境でも効率的に増えることができる。裂芽は、ウメノキゴケ類が環境に広がるための「種子のような役割」を果たす栄養繁殖器官で、胞子とは異なり、すでに藻類と菌類がセットになっているため、着生すればすぐに新しい地衣体へと成長できる。
この菌類と藻類の共生体は、光合成を行い、菌類が水分保持や紫外線防御を担う。大気汚染、特に二酸化硫黄に弱く、環境指標種として利用される。日本では本州・四国・九州・沖縄に広く分布し、都市部では減少傾向があるが、森林や農村部では普通にみられる。
クロゴケ類Cladonia spp.は、複数の地衣類グループに含まれる暗色の葉状・固着地衣で、代表例にクロボシゴケPyxine subcinereaなどがある。岩や樹皮に密着し乾燥にも適応する暗色の地衣で、小型で地表や苔の間に生育する。また林床の苔群落に混じって見られる。日当たりの良い岩の上に生息しているクロゴケは、乾燥に強く、小さな群落を作り、水分を吸収すると瞬く間に、黒かったコケはみるみる水を吸って緑の葉を広げ、光合成を始める。いずれも大気環境に敏感な「指標種」で、八ヶ岳では北八ヶ岳の白駒池周辺や縞枯山・北横岳などの亜高山帯針葉樹林でよく観察される。
この灰緑色~黒色の地衣体は、岩や樹皮に密着し、縁に粉芽を生じる。その群落は暗色で目立ち、乾燥に強い。岩石や樹皮上に広がり、比較的乾燥した環境にも適応。成長は遅いが安定した基質に長期間生育する。菌類と藻類の共生による栄養獲得のためか、環境変化に敏感で、森林の安定度や大気質を反映する。
目次へ
 北八ヶ岳の茶臼山のシラビソ・コメツガ林床は、苔類が圧倒的に優占する中に、地衣類が点在する「静かな共演の場」となり、特にサルオガセやクロゴケ類は、冷涼湿潤な亜高山帯の象徴的な地衣類であり、林床の苔とともに森に特異な美しさを醸し出している。
北八ヶ岳の茶臼山のシラビソ・コメツガ林床は、苔類が圧倒的に優占する中に、地衣類が点在する「静かな共演の場」となり、特にサルオガセやクロゴケ類は、冷涼湿潤な亜高山帯の象徴的な地衣類であり、林床の苔とともに森に特異な美しさを醸し出している。登山道周辺は「苔の聖地」と呼ばれるほど苔が豊富で、その表面や林床に地衣類が付着する。特にオニハナゴケCladonia uncialisなど、矮性低木群落内に生える種が印象的である。また茶臼山周辺は縞枯山と同様に「縞枯れ現象」が見られる地域で、立ち枯れたシラビソ林がモザイク状に広がる。この環境は倒木や立ち枯れ木に依存する地衣類に生育基盤を提供し、独特の群落構造を演出する。
茶臼山の林床は、タマゴケ・ヒノキゴケ・スギゴケ・ホウオウゴケなどの蘚類を中心に、ゼニゴケやキリシマゴケなどの苔類、そしてツノゴケ類が加わり、豊かな「苔の絨毯」を作り出している。これらは湿潤で冷涼な環境に適応した植物群であり、茶臼山の自然美を象徴する。その間にクロゴケ類やハナゴケ類が点在し、微小な群落を形成する。その間の倒木や立ち枯れ木にはサルオガセ類やウメノキゴケ類が着生し、縞枯れ現象で生じた開けた林床環境に多様性をもたらしている。
茶臼山(標高2384m)は森林限界直下に位置し、これにより、シラビソ・オオシラビソやコメツガ林床の苔群落に加え、稜線の露岩やハイマツ低木群落に高山性地衣類が生育し、亜高山帯特有の地衣類と同時に鑑賞ができる。また、縞枯れ現象と結びついた地衣類群落は、森林動態と地衣類の関係を理解する上で重要なフィールドとなっている。
地衣類の構造
地衣類は、菌類(主に子嚢菌)と藻類(緑藻やシアノバクテリア)が互いの弱点を補い合うことで共生した。子嚢菌は分解者や共生者として陸上生態系を支え、緑藻は光合成によって水域の一次生産者として働く。
子嚢菌は、真菌類の大きなグループで、全真菌の約70%を占める。真菌類とは、カビ・キノコ・酵母などを含む「菌界」に属する真核生物で、ただ光合成を担う葉緑体を持たないため、外部の有機物に酵素を分泌し分解して得られる栄養分を吸収する「従属栄養性」の生物群である
緑藻は、植物が光を利用してエネルギーを生成するための基本的な構成要素であるクロロフィル a・b を持ち、葉緑体内にデンプンを蓄える。単細胞(クロレラ)、群体(ボルボックス)、多細胞(アオサ、マリモ)など多様な体制で、陸上植物に近縁な系統もあり、進化的にも重要である。海から淡水まで広く分布し、土壌や岩上にも生育する。塩湖や氷雪など特殊環境にも適応する種もある。他生物との共生例も多く、地衣類や繊毛虫、ヒドラなどと共生する。水域の一次生産者として光合成を行い、生態系に有機物と酸素を供給する。海産緑藻(アオサ類)は沿岸生態系を支える主要な生産者で、食用(アオノリ、クロレラ)や健康食品、農業用資材としても利用される。
緑藻は、地衣類などで共生関係を結ぶ。光合成ができない菌類が「住まいと水分」を提供し、水分やミネラルを保持し、乾燥やその他の極限環境から藻類を守る。緑藻は光合成によって炭水化物を生産し、菌類に栄養を供給する。両者の細胞が密接に組み合わさり、「地衣体」という特殊な構造を形成する。この地衣体は単独の菌や藻では見られない組織分化を示し、独立した生物のように振る舞う。
最古の地衣類化石は、古生代デボン紀の約4億2千万年前に確認されているが、その系統は絶滅したと考えられる。現生の地衣類の祖先は、次の石炭紀(約3億6千万年前~3億年前)以降に出現したと推定される。地衣類化lichenizationは複数の菌類系統が独立に起こす収斂進化であり、単一の祖先からではない。
新しい火山島や裸岩の表面に最初に定着し、風化を促進して土壌生成に寄与するなど、砂漠や高山など極限環境でも生育可能で、生態系の重要な先駆者として、共生により生じる二次代謝産物(地衣成分)は、耐寒性や抗菌性など特殊な機能を持ち、非共生の菌類には見られない特徴を保持する。
地衣類の成立は、「菌類の住まい+藻類の光合成=新しい生命体」 という方程式で表せるが、進化の過程でこの関係が安定化し、複数の系統で繰り返し生じた結果、地球上に広く分布する独特な共生体が完成された。
藻類が菌類に「光合成による栄養」を供給するという交換関係が進化の中で安定化し、独立した生物のように見える「地衣体」を形成した。森林や岩場で独創的な光景を創作する生態系を生み出した。しかも、高山の地上、岩場や砂礫地など栄養塩に乏しい場所でも環境適応する。
 |
| 北八ヶ岳では、北横岳や坪庭周辺の矮性低木群落や露岩地にオニハナゴケが特に多く見られる。 亜高山帯針葉樹林の林床や倒木上にも見られるが、特に日当たりの良い稜線部やハイマツ帯で群落を形成する。特に、坪庭や北横岳稜線が典型的な生育地となる。 岩や倒木など養分の乏しい基質に定着し、初期植生の一員として土壌形成に寄与する。水分保持力が高く、周囲の苔や微小植物の生育を助ける。 乾燥や寒冷に強く、亜高山帯から高山帯の厳しい環境に適応している。 茎状の地衣体が立ち上がり、小さな花のような形態を持つため「オニハナゴケ」と呼ばれる。群落を作ると、苔や他の地衣類と混生し、緑と白のモザイク状の景観を生み出す。 |
八ヶ岳の環境と地衣類の特徴の関連は、その生育環境にあり、北八ヶ岳は標高2000〜2500mのなだらかな地形、亜高山帯針葉樹林やハイマツ低木林、露岩地が広がる。こうした環境では、亜高山帯の地衣類と高山性の地衣類が混在している。その生育環境は、主に高山の地上の、岩場や砂礫地など栄養塩に乏しい場所で、その先駆種pioneer speciesとして、裸地や痩せ地に定着し、他の植物が生育できる基盤や環境を整える。
地衣類は、晴天時には乾燥して硬くなり、雨や露で水分を吸収すると柔らかくなり光合成を行う。痩せ地や高山環境の指標となる存在である。
オニハナゴケの基本葉体は「顆粒状」で、地表に付着するその地衣類の初期葉体が「小さな粒の集合体」のように見える状態を指す。表面に皮層を備えず、微細な剥き出しの顆粒が地表に散在する。オニハナゴケではこの顆粒状葉体には、付着を助けるような「粘着性」の機能はなく、顆粒はむしろ一時的な構造で、早期に消失する性質を持っている。地衣類は、菌糸の網目構造によって基質(土壌や岩石表面)に絡みつくように付着する。これは「粘着性」に頼らず、物理的な絡みつきや浸透による付着である。
オニハナゴケはウスニン酸などの二次代謝産物を含みむが、これらは抗菌性や防御に関わるものであり、粘着性を付与するものではないとされている。代わりに子柄(樹枝状の部分)が発達する。子柄は高さ約5cm、繰り返し二叉分枝し、不等長で仮軸を形成しやすい。分枝角度は広く、90度を超えることもある。表面は黄色を帯び、平滑で光沢がある。その群落形成は、直径20cm以上のマットを作ることもある。
顆粒状葉体は初期定着の役割を果たし、裸地や岩場に付着して菌類と藻類の共生を安定化させる。一方、ネズミなどの小動物による食害は群落の破壊や再生遅延を招き、高山帯の植生形成に影響を与えることがある。
シカがオニハナゴケを食べることは、自然界でよく知られている。特に北米やユーラシア大陸の寒冷地に生息するシカ科の動物にとって、オニハナゴケは重要な食料源となる。地衣類は栄養源として優れており、特に炭水化物やミネラルが豊富で、冬場に他の植物が雪に覆われると、オニハナゴケはシカにとって貴重な食料となる。雪の下から掘り起こして食べもする。
八ヶ岳ではシカの増加によって森林や林床植生が大きな影響を受けており、北八ヶ岳特有のオニハナゴケなどのコケ類も食害や環境変化によって衰退している。シカは樹皮を食べるため、シラビソなどの樹木が枯死し、森が乾燥化する。結果としてコケ類が生育しにくくなり、ササが優占する環境へと変化している。北八ヶ岳の特徴である豊かなコケ群落が減少し、特にオニハナゴケのような湿潤環境を好む種は特に影響を受けやすいため、景観そのものが変わりつつある。
ハナゴケ類(Cladonia属)は、トナカイやネズミなどの小動物の餌になることが知られている。高山帯では冬季の食料が乏しいため、地衣類が重要な栄養源となる。群落が部分的に食べられることで、マットの連続性が失われ、その再生速度が遅れ、群落の回復には数年~数十年かかる場合がある。
食害によって裸地が再び露出し、風食や乾燥の影響を受けやすくなる。食害は群落の更新を促す一方で、過剰な摂食は植生の定着と再生を阻害する。高山帯の「脆弱な植生基盤」に直接影響を与えるため、群落の分布や密度に大きな変動をもたらす。
地衣類は、世界で約2万種、日本では約1800種が知られている。岩や樹皮を覆い、環境指標生物として大気汚染や湿度の変化に敏感である。「○○ゴケ」という和名が多いため、コケ植物と混同されやすいが、分類的には菌類である。
八ヶ岳の地衣類は 亜高山帯〜高山帯に適応した多様な種が存在する。オニハナゴケやハナゴケ類などが代表的、調査研究により、環境指標としての重要性や景観的価値が明らかになっている。
地衣類は「菌類(主に子嚢菌や担子菌)」が主体であり、藻類と共生して形成される複合体である。国際植物命名規約でも、地衣類の学名は「構成する菌類」に与えられている。
地衣類は光合成能力を持たない菌類が主体で、藻類が光合成で作った栄養を利用して生きている。形態的にはコケ植物に似ているが、内部構造は全く異なる。
目次へ
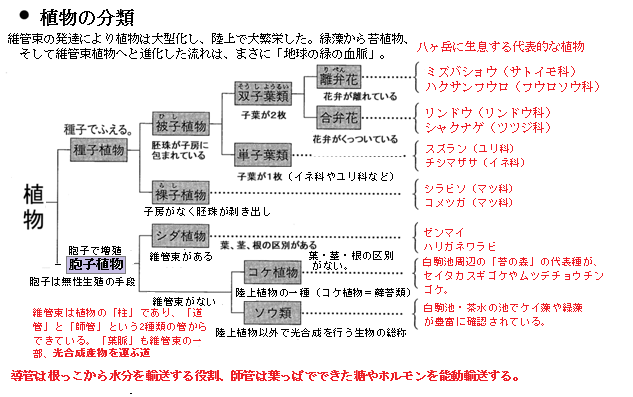 蘚苔類は「コケ植物」と呼ばれる陸上植物の仲間で、胞子で繁殖し、維管束を持たないという特徴を持っている。蘚苔類の分類上の位置は、真核生物で植物界Plantae、しかも陸上植物の一部、大きく
蘚類(コケ類)・苔類(タイ類)・ツノゴケ類 の3つに分けられる。
蘚苔類は「コケ植物」と呼ばれる陸上植物の仲間で、胞子で繁殖し、維管束を持たないという特徴を持っている。蘚苔類の分類上の位置は、真核生物で植物界Plantae、しかも陸上植物の一部、大きく
蘚類(コケ類)・苔類(タイ類)・ツノゴケ類 の3つに分けられる。つまり、光合成を行う葉緑体を持ち、独立栄養生物として生きている蘚苔類は、キノコやカビと混同されることもあるが、蘚苔類は光合成を行うため確実に植物界に属する陸上植物である。ただ維管束を持たないため、 シダや種子植物のように水や養分を運ぶ管が発達していない。
種子ではなく胞子を散布して増える。配偶体世代が主体で、「コケの本体」は染色体のが n の配偶体で、胞子体はその上に付随して発達している。日本では、約1665種。世界では、約2万種以上が知られてる。
ゲノム1組【"n"と略される】を持った雄性配偶子male gamete【♂配偶子】、または精細胞sperm cellと、やはりnを持った雌性配偶子female gamete 【♀配偶子】、または卵細胞egg cellとが融合して、ゲノム2組【"2n"と略される】を持った受精卵ができる。
種子植物
 |
| 植物も動物も、初めは個体の一部を分裂させて、新しい個体を作る無性生殖で増殖した。やがて、雄と雌の合体という有性生殖という、複雑な繁殖方法を進化さていった。それも、最初は海の中での進化で始まった。
別個体同士による細胞合体である有性生殖は、遺伝子の交換という生物の環境適応や進化のためには極めて重要であった。 有性生殖も、初期においては、雌雄差は殆どない。それが鞭毛を備えて動ける精子と、母体に寄生したまま卵細胞に分化したため、その雌雄の両性が、はっきりと分かれるようになる。 植物では、機能が異なる2つの多細胞体ができた。精子と卵子を作る配偶体と、配偶体のもとになる胞子を作る胞子体とにである。 それぞれは、独立する方向へ進化し、配偶体と胞子体という2つの世代が循環する「世代交代」を、海の中で確立した。 植物は、海の中で進化し、いろいろな器官を整えたが、その中には陸上で生活する上で障害となるものもあった。特に精子は、水中を泳ぐには都合がよい構造であるが、陸上植物では、水がなければ精子は泳げないので受精ができない。 そこで、水が殆どないところでも有性生殖を可能にする進化がなされた。この陸上という新たな環境条件に適応するためには、遺伝子の交換を伴う有性生殖が重要な意味を持つ。この改造に成功した植物群が、種子植物であった。 やがて種子のみならず、花を持つ植物が、有性生殖に関わる構造自体を改善し、海辺から内陸へ進出し、広大であるが不毛な大地を、生命力あふれる豊かな緑野へと改造し続けていった。 |
イチョウとソテツでは、花粉粒が送粉によって運ばれ、花粉管も伸びる。その後、花粉管から精子が放出されて短い距離を泳いで卵細胞に達する【花粉管受精+精子受精】。
シダ植物・コケ植物
シダ植物・コケ植物では、雄性配偶子は小さくて鞭毛を持っていて水中を泳ぐことができる【遊泳性がある雄性配偶子、「精子sperm」】。雌性配偶子は大きくて移動力がない。精子が水中を泳いで卵細胞に達して受精が起こる【精子受精】。
シダ植物やコケ植物の受精に必要な水は、微細な局所的な水膜と言える。森林の林床や岩の上に生えるコケは、雨が降った後や朝露で表面が濡れる。そのとき、葉や茎の表面に薄い水の膜が広がる。
土壌や苔の群落はスポンジのように毛細管水を保持する。細胞や葉の隙間に微細な水路ができ、そこを精子が泳ぐ。小さな水滴が胞子体や葉の間をつなぎ、精子はその水滴の中を移動する。
精子は鞭毛を持つ。顕微鏡で見るとオタマジャクシのように泳ぐ。卵細胞は造卵器archegoniumの中にあり、そこへ精子が水膜を通って到達する。この移動距離は数mm〜数cm程度あるが、コケやシダにとっては大冒険である。
藻類
藻類の精子受精は、種類によって仕組みは異なるが、基本的には「運動性を持つ精子が卵や造果器に到達し、核融合を行う」ことで成立する。
藻類は、両方の配偶子の両方が鞭毛を持っていて移動するものが多数を占める他、精子受精をするものは、両方の配偶子とも鞭毛を持たず、細胞が接したときに融合して受精が起こるものなど、さまざまなタイプがある。藻類の精子受精は、種類によって仕組みが異なるが、基本的には「運動性を持つ精子が卵や造果器に到達し、核融合を行う」ことで成立する。紅藻では水流に乗った非運動性の精子(精子体)が受精毛に捕捉され、褐藻や緑藻では鞭毛を持つ精子が遊泳して卵に到達する。
緑藻のボルボックスVolvoxは、群体を形成する緑藻の一種であり、ボルボックス属に属する生物種の総称である。ボルボックスは池や川、田などの淡水に生息し、水のきれいな場所であれば普通に見られる。水田では、日向の水際のごく浅い泥の上を見つめると、肉眼でも薄緑色の粒として見ることができる。春から秋にかけて無性生殖で繁殖し、環境が悪化してくると有性生殖を行い、乾燥を耐える接合子を形成し冬を越す。
この接合子は環境が悪化したときに生き延びるための「耐久体」として機能する。春になり環境が良くなると発芽し、再び無性生殖を行う。
緑藻の精子は鞭毛を持ち、遊泳して卵に到達する。卵に接触すると膜融合が起こり、核が合体して小型で球状の接合子を形成、受精卵は厚い壁を作り、環境が整うまで休眠する。球状の形態は表面積を最小化し、水分喪失を抑える。
多くの緑藻で接合子は球形または楕円形、その接合子は通常の栄養細胞よりもはるかに厚い細胞壁を持つ。この壁は多層構造で、セルロースや糖タンパク質、時にスピロゲラなどではリグニン様物質を含み、乾燥や紫外線に強い。
接合子は暗色化することが多く、これは光酸化ストレスから核やDNAを守る役割を果たす。乾燥や強光下でも安定性を維持できる。接合子内部では代謝が極めて低下し、核は二倍体のまま休眠する。水分が戻ると減数分裂を行い、環境条件が整うと発芽して新しい藻体へ。
• 緑藻・褐藻 → 精子は運動性を持ち、卵へ泳いで到達。
• 紅藻 → 精子は非運動性で、水流に漂い受精毛に捕捉される。
共通するのは、精子核と卵核の融合により接合子が形成され、次世代へ繋がることにある。
(緑藻における胞子体2nと配偶体nの違いは、染色体数【2nかn】と役割【胞子nを作るか配偶子nを作るか】にある。胞子体2nは二倍体2nで減数分裂により胞子nを作り、配偶体nは一倍体nで有糸分裂により配偶子【精子・卵】一倍体nを作る。
蘚類(コケ植物)の配偶体nが配偶子nを作るときに行うのは 減数分裂ではなく、有糸分裂mitosisである。蘚類の配偶体gametophyteは すでに 1倍体(n)であれば、配偶子(精子・卵)も 1倍体(n) でなければならない。そのため、染色体数を変えない有糸分裂によって配偶子が作られる。 配偶体nからできる配偶子が”n”なのは、それは、配偶子は 減数分裂ではなく、普通の細胞分裂(有糸分裂)で作られるからである。
減数分裂は「2n → n」にする反応であり、染色体数を半分にする特別な分裂であれば、
2倍体(2n) → 減数分裂 → 1倍体(n)
つまり、減数分裂は2nの細胞に固有の反応である。
蘚類の配偶体が 有糸分裂mitosisによって配偶子を作る流れ、つまり蘚類の配偶体が配偶子をつくるのは有糸分裂であることは、一般的な植物生物学の知識として確立している。
蘚類の配偶子が「有糸分裂」で作られる流れは、蘚類の配偶体(n)は、雄器(精子をつくる器官)や雌器(卵をつくる器官)を形成する。その内部で、次のように 普通の細胞分裂(有糸分裂) が起こり、配偶子が生まれる。
1倍体(n)の配偶体の細胞が分裂準備に入ると、その細胞の核が、DNAを複製して 同じ1倍体のまま 分裂の準備をする。減数分裂のように染色体数を半分にする必要がない。
その有糸分裂(M期)は、一般的な有糸分裂と同じ段階を踏む。
• 前期:染色体が凝縮し、紡錘体が形成される。
(紡錘体は、細胞分裂時に現れる「染色体を動かす微小管の装置」で、染色体を整列・分配する。細胞分裂が終わると消える。)
• 中期:染色体が細胞中央に並ぶ(赤道面に並べる)
• 後期:姉妹染色分体が引き離し、両極へ均等に運ぶ。
• 終期:核膜が再形成される
• 細胞質分裂:細胞が2つに分かれる
こうして 同じ遺伝情報をもつ1倍体細胞が2つ生まれる。
分裂してできた細胞が配偶子へ分化する
• 雄器であれば、分裂した細胞が 精子(鞭毛をもつ)に分化
【蘚類では、配偶体(葉状体や茎葉体)の表面に雄器が作られると、その内部の細胞は「精子を作る運命」にプログラムされる。雄器の内部には 雄性器官androecium特有の化学シグナルが満ちていて、そこに入った細胞は「精子母細胞」へと分化する。
これは動物の精巣で精原細胞が精子になるのと、発生学的には少し似ている。
雄器内部の細胞は、分裂を繰り返しながら“精子母細胞”へと運命づけられる。
雄器の中にある細胞は、最初は普通の体細胞に近い状態であるが、 雄器特有のシグナル(植物ホルモン様物質や局所的な遺伝子発現)によって、
• 分裂回数が制御される
• 特定の遺伝子がオンになる
• 鞭毛形成に必要な構造タンパク質が作られる
といった変化が起こり、精子母細胞へと変わっていく。雌器であれば、分裂した細胞が 卵 に分化する。
蘚類の精子は、胞子とは違い 減数分裂では作られない。 精子母細胞が体細胞分裂をして、最終的に2本の鞭毛を持つ運動性の精子(精子細胞) へと分化する。】
ここで重要なのは、配偶子は「分裂の結果」ではなく「分裂後の細胞の分化」で生まれることである。
つまり、
有糸分裂 → できた細胞が配偶子へ成熟する
という流れになる。
配偶子の完成により、精子は鞭毛をもち、水の中を泳げるようになる。卵は雌器の内部で成熟し、受精を待つ。
水があると精子が泳ぎ、受精へ。これは蘚類の生活史の特徴である。
「有糸分裂で配偶子が作られる」のは、蘚類だけの特有な現象ではない。すべての陸上植物(コケ・シダ・種子植物)に共通する特徴である。「有糸分裂」で配偶子がつくられるのも、植物の生活環が 世代交代(2倍体と1倍体の交代) で成り立っているからである。
配偶体が1倍体(n)あれば、配偶子は 1倍体(n) でなければならない。染色体の数を変えない「有糸分裂」で配偶子を作るというのが、植物の普遍的な仕組みとなる。
配偶子はどの植物でも「1倍体の配偶体が有糸分裂でつくる」というのが大原則になる。
では「減数分裂」はどこで起こるのか。
植物では減数分裂は 配偶子をつくるためではなく、胞子をつくるため に使われる。
• 胞子体(2倍体) → 減数分裂 → 胞子(1倍体)
• 胞子 → 発芽 → 配偶体(1倍体)
• 配偶体(1倍体) → 有糸分裂 → 配偶子(1倍体)
この流れは、コケでもシダでも花の植物でも変わらない。
• 「有糸分裂で配偶子をつくる」のは 蘚類だけの特殊性ではない。
• 植物界全体に共通する基本原理
• 特徴的なのは、蘚類では 配偶体が生活環の主役であること。
(シダや種子植物では胞子体が主役)
蘚類(コケ植物)で「配偶体が生活環の主役である」というのは、生活環の中で、もっとも大きく、長く生き、独立して生活しているのが配偶体であるという意味である。
コケでふつうに見えている緑の本体は配偶体(n)である。
• 地面に生えているふわふわした緑の部分
• 茎や葉のように見える部分
• 光合成をして栄養をつくり、自立して生活している
• 生殖器官(造卵器・造精器)をつくる
これが配偶体である。
胞子体(2n)は配偶体の上に“乗っているだけ”である。
• 茎の先に生える「蒴(さく)」と呼ばれる胞子嚢
• 受精後にできるが、配偶体に寄生したまま成長する
• 自分では光合成をほとんどせず、栄養を配偶体からもらう。 など独立した生活ができない。
緑藻の減数分裂は、胞子体2nから減数分裂によって単相nの胞子を形成し、それから配偶体nへ発生するという世代交代過程の一部である。動物のように配偶子n形成だけで減数分裂が起こる事はなく、また緑藻では配偶体nと胞子体2nの両方が多細胞体として存在する点が特徴と言える。
減数分裂の細胞学的特徴 は、第一分裂で相同染色体nが対合し、二価染色体2nを形成。 相同染色体2nが分離して染色体数が半減(2n → n)する。相同染色体とは、父方と母方からそれぞれ受け継いだ同じ種類の染色体のペアのことで、形や大きさ、遺伝子の並び方がほぼ同じで、減数分裂の際に対合して分離する。
その一方は母親由来、もう一方は父親由来で、長さ、セントロメア(動原体)の位置、染色パターンがほぼ同じであり、遺伝子座、つまり同じ遺伝子が同じ位置に存在するが、対立遺伝子は異なる場合がある。例えば、ある遺伝子座に「A型」と「a型」のように異なる情報を持つことがある。減数分裂での役割は、第一分裂で相同染色体が対合し、交差(乗換え)によって遺伝子が交換される【キアズマChiasma】)。これにより子孫に遺伝子の多様性が生まれる【ヒトでは 23対のうち 22対が相同染色体】。
第二分裂では.、姉妹染色分体が分離し、最終的に染色体数が半分の細胞(n)が4つできる。 この過程は動物の配偶子形成と同じ基本構造を持つが、緑藻ではその産物が胞子体と配偶体の世代交代に組み込まれる点が大きな違いとなる。
世代交代の仕組み
• 胞子体(2n) → 減数分裂 → 胞子(n) → 発芽 → 配偶体(n) → 有糸分裂 → 配偶子(n) → 受精 → 接合子(2n) → 胞子体(2n) へ
【コケ植物の配偶体は通常、私たちが目にする緑色の植物体である。この配偶体は、植物や藻類、菌類などの世代交代において、相同な染色体を1組のみ持つ多細胞体を指す。有性生殖の配偶体であれば、単相(n)の細胞からなる雌性または雄性の配偶子を形成する。
配偶子は、成熟した一倍体の生殖細胞であり、受精を目的として異性の配偶子と結合することができる。配偶子には、雄性配偶子(精子)と雌性配偶子(卵子)の2種類がある。配偶子は、受精によって新しい配偶体を形成するために必要な細胞で、受精が行われると、配偶子が融合して二倍体の接合子が形成され、これが成長して新しい配偶体になる。】
• この循環を「世代交代」と呼び、緑藻では同形世代交代【胞子体と配偶体がほぼ同じ形】や異形世代交代【大きさや形が異なる】など多様なパターンが存在する。
•胞子体:二倍体、胞子を作る世代
•配偶体:一倍体、配偶子を作る世代
•両者が交互に現れることで、緑藻は環境変動に適応する多様な生活環を進化させてきた。)
蘚類と苔類
蘚類(せんるい)と苔類(たいるい)は、どちらも「コケ植物」に属すが、形態や胞子体の特徴に大きな違いがある。蘚類はスギゴケやミズゴケのように、「硬く丈夫な茎と葉が分かれる茎葉体と胞子体」を持ち、群生して地面にしっかり根付く。 八ヶ岳では、スギゴケやミズゴケが冷涼で湿潤な環境に群生し、林床や湿地で重要な役割を果たしている。これらは「蘚類」に属し、苔類とは葉の構造や胞子体の特徴が異なるため苔類には分類さない。ミズゴケは、一般的に胞子体を形成することが少なく、枝葉などから容易に新たな個体を再生する。オオミズゴケの胞子体は、黒光りしている胞子嚢が特徴で、比較的短時間で分解するため、出合うことは少ない。その胞子体は配偶体の上で生活し、配偶体より小さく、配偶体の光合成に依存している。ミズゴケの胞子散布時には、胞子が爆発的に射出され、蓋は胞子と共に吹き飛ばされる。
 |
| 白駒池(標高約2,115m) の周囲は「苔の森」として知られ、セイタカスギゴケやコセイタカスギゴケが群落を形成している。2008年、日本蘚苔類学会は「日本の貴重なコケの森」に選定した。 北八ヶ岳の縞枯山・北横岳・茶臼山周辺では、コメツガやシラビソ林の林床にスギゴケ類が広がり、幻想的な苔の森を形成している。 |
 |
| スギゴケの胞子体は、配偶体(茎葉体)の上に形成される「蒴」と「蒴柄」から成り、胞子を作って散布する。胞子体には、光合成能力はほとんどなく、養分を配偶体から受け取って成長する。 蒴(胞子嚢)は蒴柄の先端に形成されるカプセル状の器官で、円柱形に近く、先端に尖った帽(キャップ状の構造)を持つ。表面は長い毛に覆われることが多く、乾燥や外的環境から胞子を守る。 配偶体の形成は、雄株には造精器(精子をつくる器官) 、雌株には造卵器(卵細胞をつくる器官)が形成される。両者の間で水を介した受精が行われる。つまり、雨や霧などの水分が媒介となって精子が卵へ到達する。水が存在する湿潤環境が受精の必須条件になる。 配偶体の造卵器内で卵と精子が結合し、受精卵(接合子)ができる。 受精卵が分裂を繰り返し、蒴柄と蒴を形成する。 核相は2n(複相)の蒴の内部で減数分裂が起こり、多数の胞子が形成される。蒴の頂部が開いて胞子が放出され、風や水滴によって広範囲に運ばれる。 |
蘚類(スギゴケなど)では蒴の口に「歯」のような構造が並ぶ。その蒴歯は湿度に敏感で、乾燥すると開き、湿ると閉じる性質を持つ。これにより胞子は一度に放出されず、環境条件が整った時に少しずつ散布される。蒴の上部には蓋があり、成熟すると外れて胞子が出やすくなる。 蒴を覆う「帽(ぼう)」は発達途中の胞子を保護し、動物による食害から守る役割も果たしす。基本的には風散布が主であるが、雨滴の衝撃で胞子が飛び出すこともある。一部のコケでは昆虫や小動物の体表に付着して運ばれる「付着散布」や、消化管を通過して発芽する「消化散布」も確認されている。
多くの蘚類では胞子散布は春から初夏(5〜6月)に集中する。これは湿度や風の条件が胞子の発芽に適しているためで、胞子散布は、風・水滴・動物 を利用して広がる。つまり、蒴は単なる「胞子の袋」ではなく、環境条件に合わせて 段階的・時期的に調整 される。蒴は、環境に応じて胞子を少しずつ解き放つ精密な調整装置と言える。
北八ヶ岳の白駒池周辺や麦草峠付近の『苔の森』が、スギゴケやミズゴケの美しい大群落に感激する代表的な場所となっている。冷涼で湿潤な亜高山帯の針葉樹林の原生林の林床が、薄い土壌でありながら豊かな腐植層を育て、局所的で細やかな地形の変化を示す微地形による水分保持などにより、苔の繁茂を支える環境を整えている。白駒池周辺(北八ヶ岳) は標高約2,100mに位置し、一面の苔で覆われる『苔の森』として知られ、観光・研究の両面で注目されている。
麦草峠〜縞枯山周辺のコメツガ林やシラビソ林の林床は、標高が高く、土壌が薄く栄養が乏しいため、他の植物との競合が少ないため、多様な苔類群落が先駆的に広大に繁茂する環境を整えている。加えて冷涼な気候と適度な降水が苔の繁殖を促す。白駒池周辺では、セイタカスギゴケやコセイタカスギゴケがよく見られる。イワダレゴケやチシマシッポゴケなどと混生し、多様な群落を形成している。
セイタカスギゴケは比較的背丈が高く湿潤な環境に群落をつくるスギゴケ類で、コセイタカスギゴケはやや小型で冷涼な山地の林床や斜面に繁茂する。両者とも雌雄異株で、胞子によって新しい場所に定着し群落を形成する。セイタカスギゴケの雄株は茎頂に造精器(雄器官)をつけ、雌株は蒴(胞子体『造卵器【雌器官】』)を形成する。雄株の茎頂に造精器が集まり、内部で多数の精子がつくられる。精子は鞭毛を持ち、運動能力がある。
胞子体は葉緑体を持たず、成長及び胞子を作るためのエネルギーは全て配偶体に依存する。蒴の中で胞子が成熟すると減数分裂が起こり、たくさんの胞子が形成される。蒴の頂部が開き、その減数分裂でできた胞子が風に乗って散布される。胞子散布後、胞子が湿った地表に落ちると発芽し、糸状の「原糸体」が伸びる。これがコケ植物の幼形で、ここから配偶体が分化する。原糸体が広がり群落をつくる。 その後配偶体の形成は、雄株には造精器(精子をつくる器官) 、雌株には造卵器(卵細胞をつくる器官) が形成される。両者の間で水を介した受精が行われる。つまり、雨や霧などの水分が媒介となって精子が卵へ到達する。水が媒介となるため、湿潤環境が受精の必須条件になる。つまり、放出された胞子は、発芽後、原糸体という微小な体を経て、配偶体に成長する。光合成能力が高く、しかも苔類の中では比較的乾燥にも耐える。
精子が卵細胞に到達すると受精が成立し、二倍体の胞子体が雌株の上に育つ。胞子体は栄養を雌株から受け取りながら成長する。次世代へ 胞子体の蒴で再び減数分裂が起こり、新しい胞子が形成され、散布されることで次世代へと循環する。
(胞子体は受精後に形成される世代で、核相は2nで、蒴内部で「胞子母細胞」が分化し、そこで減数分裂が行われて単相nの胞子が得られる。蒴内には中心のコルメラcolumellaと周囲の胞子形成組織sporogenous tissueがあり、後者から胞子母細胞が生じる。
蒴の壁【外皮】が成熟に伴って乾湿で変形する性質をもち、歯状器など開口部の構造の機械的役割により、乾燥条件次第で効果的な散布が可能になる。
減数分裂のステップ は、胞子母細胞の形成に始まり、その蒴の胞子形成組織で、体細胞分裂により前駆細胞が増え、各細胞が大きくなって胞子母細胞へ分化する。核相は2n 。
胞子体では「胞子母細胞2n」が減数分裂を行い、染色体数が半減して n(単相)の胞子が形成される。
胞子体から胞子形成までの流れは、受精卵(接合子2n)が分裂・成長して胞子体になる。その胞子体は二倍体(2n)で、蒴(胞子嚢)の内部に「胞子母細胞」が分化して形成される。
胞子母細胞は2nの染色体を持ち、その減数分裂は 2回連続の分裂(第一分裂と第二分裂)から成る。
減数第一分裂(染色体数が半減する分裂)は、
前期:相同染色体が対合し「二価染色体」を形成。
中期:二価染色体が赤道面に並ぶ。ここでキアズマChiasmaを介して遺伝的組換え(交叉)が起こり、多様性が生まれる。
後期:相同染色体が分離し両極へ移動する【二細胞期へ】。
終期:核膜が再形成され、細胞質分裂が起こり、染色体数が半分(n)になった細胞が2つできる。
減数第二分裂(染色体数は変わらず、DNA量が半減する分裂)では、
第一分裂でできた2つの細胞がさらに分裂する。
中期:染色体が赤道面に並ぶ。
後期:姉妹染色分体が分離。
終期:核膜形成と細胞質分裂。
結果として 4つのn細胞(胞子) が得られる。
つまり、各二細胞が連続して第二分裂を行い、姉妹染色分体が分離。最終的に四分子(テトラッド;減数分裂の際に形成される4つの姉妹染色体のグループを指す。)となる。 4つの娘核を生み【核相はn 】、四分子期に各胞子の初生壁が形成され、その後、外層(しばしばスポロポレニンを含む耐久性の高い外壁)が沈着する。 テトラッド分離酵素の作用や機械的過程で、4個の独立した胞子に分かれる。
ポイントを整理すれば、
胞子母細胞(2n) → 第一分裂後(n) → 第二分裂後(n)
最終産物:4つの単相(n)の胞子は、相同染色体のランダム分配(遺伝的組換え)により遺伝的多様性の要因となる。
胞子母細胞は「親の染色体を半分にして子孫へ渡す工程の場」となり、第一分裂で染色体数を半分にし、第二分裂でコピーを分けることで、最終的に4つの胞子を完成させる。
蒴壁と開口部構造が成熟し、乾燥時に開きやすく、湿潤時に閉じやすい性質が整う。これにより、風散布向きのタイミングで、蒴の頂部が開き、胞子が放出される。
蒴頂部が開き、乾燥・気流に合わせて胞子が拡散する。胞子は適地で発芽して原糸体を生じ、その後に雄株・雌株の配偶体へ分化し、次の受精サイクルへ繋がる。
これらのうち、「蒴の中で減数分裂が起こり胞子が形成され、成熟後に頂部が開く」という点はセイタカスギゴケで観察事実として記されている。)
八ヶ岳全体では254種以上の蘚類が確認され、希少種や新種も報告されている。ミズゴケモドキなど絶滅危惧種も近年八ヶ岳で確認されており、学術的にも重要な地域として認識されている。
 |
| 高標高環境(約2100m)の白駒の池は、日本で最も標高の高い天然湖のひとつで、周囲は亜高山性針葉樹林(コメツガ・シラビソなど)に覆われている。この冷涼で湿潤な環境が、スギゴケ(左)やミズゴケ(右)の旺盛な生育を支えている。 スギゴケ類は倒木や岩の表面に群落を形成し、乾湿の変化に強い傾向を.示す。一方、ミズゴケ類は池周辺の湿地や水際に優占し、水分保持力が高いため、局所的に小さな湿原環境を作り出す。 ミズゴケはスポンジ状の細胞構造を持ち、池周辺の水分保持に大きく寄与する。スギゴケは比較的乾燥にも耐え、林床の安定化に役立つ。両者の共存は水分動態の緩衝作用を生み、原生林の生態系を支えている。 |
八ヶ岳ではホソバミズゴケなどが確認されており、地域の湿原や原生林の生態系に不可欠な存在になっている。八ヶ岳のホソバミズゴケSphagnum girgensohniiは、亜高山帯針葉樹林帯のシラビソやコメツガ林の林床に多い。八ヶ岳の調査では、ホソバミズゴケは森林床の腐植土が厚い場所に限って群落を形成することが確認されている。溶岩流や岩塊の微地形は小さな窪地をつくり、雨水や雪解け水が滞留しやすい。
ホソバミズゴケは乾燥に弱く、常に湿った環境を必要とするため、こうした窪みが生育に最適な環境となる。他の蘚類よりも厚い腐植層を好み、その窪地に堆積した有機物が重要な基盤となる。林床や斜面下部の凹地など、湿潤で腐植層の厚い場所に群落を形成する『森林性ミズゴケ』である。湿原だけでなく森林内の腐植土上に生育する点が特徴で、八ヶ岳では溶岩流や岩塊による微地形の凹地に多く見られる。ただ、森林床全体には多様な蘚苔類が生育する。ホソバミズゴケは窪地という限られたニッチ環境に適応することで、より乾燥に強い耐性力のあるコケ類との競合を避けている。結果として「孤立したコロニー」を形成する。林床に離散的なコロニーを作り、他の蘚類よりも厚い腐植層上を好んで生育する。多数の孔がある葉の透明細胞は、水分を保持する能力が高く、湿潤環境の維持に貢献する。
ホソバミズゴケが、雌雄同株または雌雄異株であるのは、繁殖成功率と遺伝的多様性の両立を図るためで、同じ個体に雄器官(造精器)と雌器官(造卵器)が存在するため、近接した環境でも受精が成立しやすい。特に孤立した窪地や小群落で生育するホソバミズゴケにとって、少ない繁殖機会を逃さないことが種の保存上重要になる。雌雄同株であれば、個体数が少ない環境でも繁殖可能で、局所的な絶滅リスクを減らせる。一方、異なる株間で受精が行われば、近交弱勢を避け、遺伝的多様性を維持できる。これは環境変動や病害への耐性を高める。異株性は群落内での遺伝的交流を促進し、長期的に健全な集団を維持する。
その胞子は直径30〜33µmの黄褐色、その生息域は溶岩流や岩塊による微地形の窪みに限定されるようだ。その高い水分保持能力により、八ヶ岳の湿潤な森林生態系を支える重要な蘚類となる。
八ヶ岳の白駒池周辺の『苔の森』を歩くと、湿原だけでなく林床の暗い凹地にホソバミズゴケの群落が見つけられる。高さ15〜20cmほどで丈夫な茎を持ち、黄緑〜灰緑色。枝葉は覆瓦状(ふくがじょう)に密生し、上半部が強く反り返る。「覆瓦状に密生する」とは、ホソバミズゴケの枝葉は卵状披針形(らんじょうひしんけい)で、基部が丸みを帯びて広く、先端が細く鋭く伸びる葉形が互いに斜めに重なり合いながら茎を覆うように並ぶ。瓦屋根のように「ずらして重ねる」ため、隙間が少なく、全体が密な層を形成する。瓦を重ねたように枝葉が互いに重なり合い、茎を取り囲むように密に配置される形態であるため、水分の.保持・光の効率的利用・外的環境からの保護に役立つなど多くの利点が備わる。
(卵状披針形の卵状(卵形)とは、基部が広く、先端に向かって狭まる形。卵を縦にしたような輪郭。)
披針形とは、細長く先端が鋭く尖り、基部がやや広い形。槍の穂先に似ている。
卵状披針形とは、両者の中間で、基部が卵形のようにふくらみ、先端に向かって披針形のように細く尖る。つまり、基部が丸みを帯びて広く、先端が細く鋭く伸びる葉形を示す。)
ホソバミズゴケの枝葉の上半部は強く反り返り、葉の背面には楕円形の孔が多数存在する。この形態により、水分保持の効率が高まり、光合成に必要な表面積を確保しつつ群落を安定させる。ただ、ホソバミズゴケを含むミズゴケ類は、茎葉体には気孔を持たず、胞子体(朔)の頸部にのみ気孔を備えている。代わりに、透明細胞の孔と葉緑細胞の組み合わせで水分保持とガス交換を行い、湿潤環境に特化した生態戦略をとっている。光合成やガス交換は、葉のこの特殊な細胞構造を通じて行われ、種子植物のように葉の表面に気孔を持たず、ミズゴケ類の茎葉体では二酸化炭素の取り込みを気孔に依存していない。気孔がないことで蒸散による水分損失を防ぎ、窪地や腐植土上で安定して生育できる。水分保持と光合成の効率的な仕組みにより、泥炭地で炭素を固定し、地球規模の炭素循環に関与する。
ミズゴケの葉は一層の細胞からなり、死んだ細胞で、大きな孔を持ち水を保持する透明細胞hyaline cellsと、光合成を行う生きた葉緑細胞chlorophyllose cellsからなり、二酸化炭素は水に溶けた炭酸イオンの形で透明細胞から供給され、葉緑細胞は光合成に利用される。ホソバミズゴケの枝葉透明細胞には、背面に多数の楕円形の孔、腹面に少数の円形の孔があり、水分やガスの出入りを助ける。ミズゴケ類は胞子体の一部に気孔を持ち、そこでガス交換が行われる。
 |
| ホソバミズゴケの群落は主に北八ヶ岳の亜高山帯針葉樹林の林床や湿原に多く見られ、特に白駒池周辺の『苔の森』や白駒湿原に豊富。 『苔の森』では、ホソバミズゴケを含む多様な蘚苔類が群落を形成している。湿度が高く、林床に厚い腐植層に特に多く見られる。ワタスゲなど湿原植物とともにホソバミズゴケが広がり、湿原の基盤を支える。 |
 |
| 蒴蓋が外れて胞子が放出され、風や水滴によって広範囲に拡散する。湿った基質に落ちると胞子が発芽し、糸状の原糸体を形成。原糸体は光合成を行い、次世代の配偶体を育てる。 原糸体から芽が立ち上がり、ホソバミズゴケ特有の茎葉体へと成長する。 胞子散布による新規定着と、栄養繁殖(茎の断片からの再生)を組み合わせて群落を維持する。 |
光合成速度は光強度や温度に強く依存し、種ごとに最適条件が異なる。泥炭湿地では低温・低栄養環境に適応し、炭素固定に大きく寄与する。
ホソバミズゴケの特徴ある枝葉の形態は、卵状披針形の枝葉が覆瓦状(ふくがじょう)に密生し、透明細胞には背面に多数の楕円形の孔、腹面に少数の円形の孔がある。その断面では腹側が広い台形状をしており、光合成細胞が効率的に光を受ける構造になっている。
その生息域が溶岩流や岩塊の窪みなど、森林床の湿潤で腐植土が厚い場所に限定されるため、乾燥回避と水分保持に特化した光合成戦略を持つが、他のミズゴケ類と基本メカニズムは同じで、透明細胞が水と炭酸イオンを供給し、葉緑細胞で光合成を行う。
その違いは種ごとの形態と環境応答にある。
オオミズゴケS. palustreは、湿原で大規模群落を形成し、光合成速度が比較的高い。ヒメミズゴケやイボミズゴケは、光阻害や温度依存性が強く、環境条件に敏感である。ホソバミズゴケは、森林床の窪地に特化し、光合成細胞の配置がやや異なる。
共通点は、ミズゴケ類は気孔を持たず、透明細胞と葉緑細胞の二重構造で光合成をする。その違いは、ホソバミズゴケは枝葉の形態と細胞配置が特殊で、森林床の湿潤環境に適応し、他のミズゴケは湿原や泥炭地で群落を形成し、光合成速度や環境応答が異なる。
ホソバミズゴケとその他のミズゴケ類の「光合成の仕組みの違い」を具体的に整理すると、細胞配置・葉形態・環境応答の三点で差が見えてくる。ホソバミズゴケの枝葉は卵状披針形で、覆瓦状に密生している。透明細胞hyaline cellの背面に多数の楕円形の孔、腹面に少数の円形孔があり、水分保持とガス交換を効率化している。葉緑細胞は腹側に広い台形状で配置され、森林床の弱い光でも効率的に光合成できる。
湿原性ミズゴケ、例えばオオミズゴケ S. palustreとアオミズゴケ S. cuspidatumの葉はより細長く、その透明細胞の孔の配置が種によって異なる。葉緑細胞は透明細胞に挟まれるように配置され、強い光環境でも光合成効率を保てる。しかも、水面近くに群落を形成するため、光量が比較的豊富で、光合成速度が高い。
ホソバミズゴケの方は、森林床の窪地に生育するため、光が弱い環境に適応する。枝葉の特殊な形態と細胞配置で「弱光・湿潤な窪地」で最適化し、光合成速度は遅いが安定性は高い。森林床の微地形に依存し、小規模・孤立的な群落を形成して、局所的な炭素固定に寄与する。
• 光合成速度は低めだが、低光量でも安定して維持できる。
• 水分保持能力が高く、乾燥に弱いが窪地の湿潤環境で安定する。
湿原性ミズゴケは、湿原や泥炭地で「強光・水位変動」に適応し、光合成速度が高く、群落規模も大きい。広大な群落を形成し、泥炭地の主要構成種となり、地球規模の炭素循環に大きく関与する。
• 泥炭湿地や開けた湿原に群落を形成するので光が強い環境に適応する。
• 光合成速度は高く、群落全体で炭素固定量が大きい。
• 水位変動や強光に対しても比較的素早い応答を示す。
苔類はゼニゴケ、ジャゴケのように「柔らかく脆い葉状体や短命な胞子体」を持ち、油体など独特の細胞構造を備える。油体は、特にコケ植物に見られる細胞内小器官であり、主に二次代謝産物を蓄積する役割を担う。油体は、細胞内に特定の生理活性物質を高濃度で蓄積することができ、これにより植物は効率的に防御することができる。これらの生成化合物は、抗菌作用や抗ガン活性を持つものが多く、植物が外部のストレスや食害から身を守る。
苔類の特徴は、茎葉体タイプと葉状体タイプの両方がある。葉に中肋はないが、葉が大きく裂けて背腹の分化が見られる。それは単なる形態の違いではなく、陸上生活への適応に伴う器官形成の進化を物語っている。陸上に進出した植物は、光を効率よく受ける必要があり、苔類の中でも「植物体である葉状体を形成する(ゼニゴケ型)」から「腹葉が茎の幅の約2倍あり、2裂し、裂片が開き、基部は片方の葉と癒合する茎葉体(ウロコゴケ型)」へ進化するにつれ、光を受ける背面dorsalと基質に接する腹面ventral)が明確に分かれ、機能分化が進んだ。
(ウロコゴケ類のような茎葉体の苔では、背側(上側)に左右2列の側葉があり、腹側(下側)に1列の腹葉という 3列配置 が基本構造。腹葉は、側葉よりもずっと小さく鱗片のように見えるが、茎の下面に沿って並ぶ小形の葉茎が地面や腐木にしっかり貼りつき、また腹側の小さな葉により、水膜が保持されやすくなる。その背腹性の強調は、陸上生活への適応として、光を受ける背面と基質に接する腹面が機能分化し、それぞれの能力を向上させた。)
胞子体は短命で弱々しく、蒴【胞子囊】は球形や円筒形で、成熟すると4つに裂けて胞子を散布する。その原糸体は塊状で、ひとつの胞子からひとつの配偶体ができるだけである。柔らかく脆い感じがある。
目次へ