Top 車山高原 車山の山菜 車山日記 車山ブログ 車山高原の野鳥 車山7月日記 車山8月日記
| DNA 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精 ブドウ糖とデンプン 植物の運動力 光合成と光阻害 |
 |
| 目次 |
| 1)チラコイド反応と「ATP」「NADPH」 |
| 2)エネルギー生産とATP合成 |
 効率の良いエネルギー生産が、地球上の生命の進化に大いに貢献した。しかも、エネルギー生産とATP合成を行なう効率のよい機構が、生物の歴史の初期に出現していた。
効率の良いエネルギー生産が、地球上の生命の進化に大いに貢献した。しかも、エネルギー生産とATP合成を行なう効率のよい機構が、生物の歴史の初期に出現していた。その仕組みには、膜における電子伝達が使われる。それが、今現在でも、地球上の生命が生存していくための中心的な仕組みとなっている。
細胞は、膜を使って行われるこの機構により、様々な材料からエネルギーを獲得する。光合成による光エネルギーの化学結合エネルギーへの変換や、細胞呼吸(cell respiration)により食物分子から大量のATP生産などで、この仕組みが重要な役割を果たす。
膜における電子伝達は、最初は細菌として30億年前に出現した。その細胞の子孫たちは、現在、地球上の多種多様な生命体に進化し、陸上と水中のあらゆる場所に生存している。かつて取り込まれた真核細胞内で、葉緑体とミトコンドリアとして、自律性を維持したまま不可欠な存在となっている。
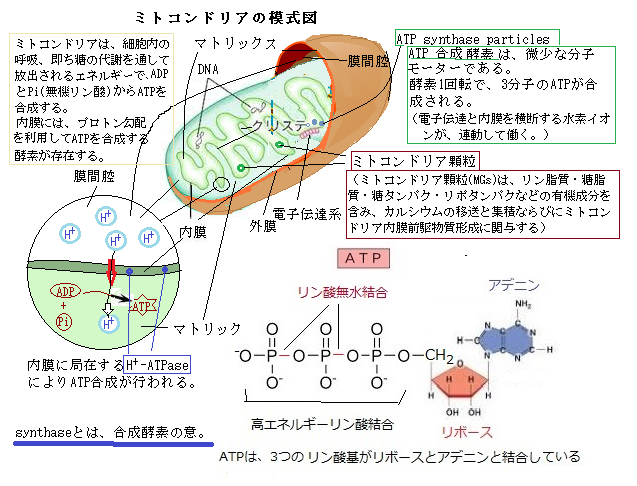
葉緑体は、ミトコンドリアと構造は似ている。その重要な違いは、ミトコンドリアよりも大きく、チラコイドと呼ばれる区画がもう1つ加わることにある。ミトコンドリアの内膜は、折り畳められ、多数のクリステ(Cristae)を作っている。クリステは、ミトコンドリア内膜の中に、化学反応を起こすため、表面積を広げ、特徴的なひだ構造を形成して、好気呼吸の能力を最大化させている。その内膜には、電子伝達系やATPを作るATP合成酵素など、酸化的リン酸化にかかわるタンパク質が含まれている。
葉緑体の内膜にも、様々な膜輸送タンパク質が埋め込まれているが、光合成装置は含まれていない。その内膜が取り囲む広い空間がストロマ(stroma)と呼ばれ、ミトコンドリアのマトリックに相当し、代謝に関わる酵素が多数含まれている。その中に、平たい円盤状の袋が層状に積み重なる第3の膜、チラコイド膜(thylakoid membrane)があり、光補足性・電子伝達系・光合成でATPを作る出すATP合成酵素などが存在する。
チラコイド反応では、その反応の最終段階でNADPH (ニコチンアミドアデニンジヌクレオチドリン酸)と並んで、もう一つの化学エネルギーが作られる。それが「ATP(アデノシン3リン酸)」という物質だ。 植物が光をエネルギー源としているとしても、直接光を使って細胞を反応させているわけではない。エネルギーを使う反応には、動植物を問わず、多くの場合ATPのエネルギーが使われる。
ATPは高エネルギーリン酸化合物として、生体のエネルギー代謝で重要な働きをする。 ATPはアデノシン(Adenosine)という物質にリン酸が3つくっつく構造をしている。アデノシンはアデニンとリボースからなるヌクレオシド(nucleoside は塩基と糖が結合した化合物)の一つで、神経系に多く存在する物質でDNAやATPの材料となる。生化学過程でもATPやADP(アデノシン2リン酸;Adenosine diphosphate)の一部としてエネルギー輸送に関わったり、環状AMP(環状アデノシン一リン酸;cAMP)としてシグナル伝達に関わったり、タンパク質をリン酸化する酵素を活性化したりする。
光合成の第1段階は、チラコイド膜にある電子伝達系が、電子伝達のエネルギーを利用してプロトン(プロトンprptonとは陽子や水溶液中の水素イオンを言う。ここでは水素イオンを指す)をチラコイド膜に包まれたチラコイド内腔に取り込むと、その結果生じたプロトン勾配を利用して、ATP合成酵素によりATPの合成をする。
「光化学系Ⅱ」の反応中心に電子が流れ、光エネルギーにより反応中心にあるクロロフィルスペシャルペアが励起状態になる。反応中心ではクロロフィルスペシャルペアが一連の電子運搬体の直ぐ隣に在って、励起したクロロフィルスペシャルペアから1個の電子が移動性の電子運搬体であるプラストキノン(plastoquinone)に渡される。次に水分解酵素によって水から引き抜かれた1個の電子がクロロフィルスペシャルペアに戻される。水から電子の引き抜きに関わるマンガン原子のクラスターは、2分子の水から4個の電子が引き抜き、水から引き抜かれ酸素がその副産物として放出される。地球上の大気中にあるほぼすべての酸素O2を作り出してきたのが「光化学系Ⅱ」のマンガン原子のクラスターを含む水分解酵素の働きによるとは驚きである。
その光合成と酸化的リン酸化と異なるのは、電子伝達系に送られた高エネルギー電子が、太陽光のエネルギーを吸収したクロロフィルに由来するからだ。その上、高エネルギー電子が、最終的に到達する場所が、O2ではなく、NADP+に与えられて、その電子を還元されNADPHが生じる。
近くにある第2の光化学系「光化学系Ⅰ」も、同時に太陽光からのエネルギーを捕捉する。これらの「反応中心」は、その高エネルギー電子を別の移動性電子運搬体に渡し、電子は酵素の一つに運ばれてNADP+をNADPHに還元するのに使われる。これら2つの光化学系の連携した働きにより、ATP(光化学系Ⅱ) とNADPH(光化学系Ⅰ)の両方が生産され、光合成の第2段階で使われる。
その第1段階での「チラコイド反応」で生成されたATPとNADPHに続く第2段階でのCO2から糖への変換は、ATPとNADPHを用いて「ストロマ」内の「炭素固定回路」でCO2が同化(固定)され、糖・アミノ酸・脂肪酸など多様な有機分子が作られる。
光合成を大雑把に表現すれば、生合成過程といえる。植物細胞がCO2から有機分子を合成するには、ATPというかたちでの大量エネルギーの投入と、活性運搬体NADPHによる大量な還元力の供給が欠かせない。 これらの「チラコイド反応」や「ストロマ反応」は、相互に巧緻なフィードバック機構で繋がっているが、実態は、植物は適切な好条件下でしか、糖は作り出せない。
炭素固定に必要な酵素の中には、暗条件では不活発になり、太陽光を受け取って始まる電子伝達反応で、再活性化されるものもあるのだ。 チラコイド内腔は、細胞の他の小器官とは完全に切り離されて密封されている。そうした構造であるため、葉緑体はミトコンドリアよりも大きなH2勾配が作られる。
葉緑体には、H2が運び込まれるための小さな専用区画があるため、大きなH2の濃度差が生じる。チラコイドのH2勾配に蓄えられるエネルギーの大部分は、チラコイド内腔とストロマの間のH2の濃度差に依存している。
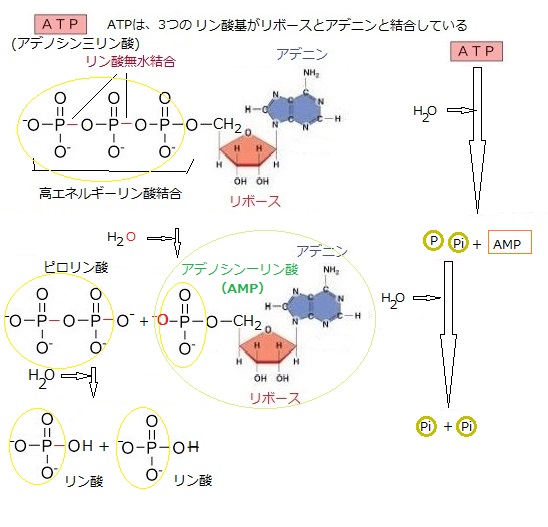 ATPのアデノシンにつく3つリン酸はそれぞれがマイナスの電荷をもっているため、3つのリン酸それぞれが反発しあい、その結合が切れた時に、その反発エネルギーが放出される。こうしてATPがリン酸を2つだけ持つADPと1個のリン酸に分解される。その際に放出されたエネルギーは、ちょうど細胞内の様々な反応を進めるのに必要なエネルギー程度の大きさになっている。このためATPの分解と組み合わせることによって、細胞内の反応を進めることができるようになる。
ATPのアデノシンにつく3つリン酸はそれぞれがマイナスの電荷をもっているため、3つのリン酸それぞれが反発しあい、その結合が切れた時に、その反発エネルギーが放出される。こうしてATPがリン酸を2つだけ持つADPと1個のリン酸に分解される。その際に放出されたエネルギーは、ちょうど細胞内の様々な反応を進めるのに必要なエネルギー程度の大きさになっている。このためATPの分解と組み合わせることによって、細胞内の反応を進めることができるようになる。「ATP」は、「ADP(アデノシン2リン酸)」という物質に分解されると時に、エネルギーを放出し、それがあらゆる生命活動の源になる。植物を含むすべての生物は、細胞分裂や運動、体を作る物質の合成など、生きていくために必要なエネルギーの受け渡しに、この「ATP」のエネルギーを利用する。
「ATP」を「ADP」に分解しエネルギーを取り出した後、一定の条件下でエネルギーを与えると、「ATP」を再生産する。このグルグル循環する再生産とその利用、及びその役割の大きさから「ATP」を、「エネルギーの通貨」とも呼ぶ所以である。またある物質とリン酸が結合する反応のことを、「リン酸化」といい、「ADP」から「ATP」が作られることも「リン酸化」の一つである。 この「ATP」が持つ化学エネルギーが、後続する「ストロマ反応」で重要な役割をする。
物資に電子を受け渡す還元反応を引き起こすには、「チラコイド反応」における光エネルギーのように、外からのエネルギーの投入が必要だ。「ATP」の化学エネルギーによって二酸化炭素が還元され、電子を受け取りブドウ糖が合成される。植物は、この「ATP」も光のエネルギーを元手に作られている。
目次へ
生物にとって効率のよいエネルギー生産は不可欠である。特に、ATPは数百もの代謝反応でエネルギーの受け渡しに関わっている。動物細胞・菌類や一部の細菌のATPは、その食物の酸化的分解で放出されるエネルギーによって合成され、植物細胞や一部の細菌では、光エネルギーの捕獲反応で得られる。
植物の光合成は、結局は光エネルギーを元手にした生合成過程と言える。植物細胞がCO2から有機分子を合成するには、ATPという大量のエネルギー入力と、活性運搬体NADPHと言う大量の還元力の供給が必要だ。
植物細胞内のシアノバクテリアと言う自由生活性の光合成生物は、一対の「光化学系(PS)」による化学反応によりATPとNADPHを合成する。その一対の構造は似ているが、反応中心のクロロフィルスペシャルペアから放出される高エネルギー電子の扱いが異なる。
第1の「光化学系(PS)」は、歴史的な理由にから「光化学系Ⅱ」と呼ばれるが、光エネルギーを吸収すると、その反応中心のクロロフィルスペシャルペアは、水から取り出された電子を運ぶ電子伝達系の最後に位置する電子受容体として働いている。その水から水分解酵素の働きにより取り出された電子は、光合成電子伝達系の一部であるプラストキンと言う移動性の電子運搬体に渡される。その際に酸素が副産物として発生する。
プラストキンは、高エネルギー電子をシトクロムb6-f複合体というプロトンポンプ(プロトンポンプは、生物体内で光エネルギーなどを利用して水素イオンを輸送する)によってくみ上げられ、移動性の電子運搬体であるプラストシアニン(銅を含むタンパク質)に運ばれる。このプロトンポンプはミトコンドリア内膜のプロトンポンプと同様に、電子の移動によって電気化学的プロトン勾配を作る。プラストシアニンは、この電子を「光化学系Ⅰ」に渡し、励起状態のクロロフィルスペシャルペアが失った電子に置き換わる。この「光化学系Ⅰ」が次の光を吸収すると、この電子のエネルギー状態が更に高くなる。
「光化学系Ⅱ」を引き継ぐ「光化学系Ⅰ」も、太陽光からのエネルギーを捕捉すると、こちらの反応中心クロロフィルスペシャルペアの電子のエネルギー状態は高まり、別の移動性電子運搬体フェレドキシン(ferredoxin)に渡された時には、NADP+をNADPHに還元するのに必要な高エネルギー電子に変わっている。こうして2つの「光化学系(PS)」が縦に並び連携して働き、「光化学系Ⅱ」ではATPが、「光化学系Ⅰ」ではNADPHが生産され、2つの「光化学系(PS)」の高エネルギー電子を効果的に生成させている。
水の電子の強固な結合を、2つの「光化学系(PS)」それぞれが取り込んだ光が提供する余分なエネルギーをつぎ込んで、水から電子を取り出し、電子が緩やかに結合するNADPHを還元し、電子伝達系を働かせてチラコイド膜を越えてH+移動させる。しかもATP合成酵素は、光由来のエネルギーの一部を取り込んでATPを生産している。
アデノシン三リン酸(ATP)は、アデノシンという物質に3つのリン酸基(P)が、2つの「リン酸無水結合」で結合されている。これらの結合が切れる時に多量のエネルギーが生じる。特に先端のリン酸基は、加水分解で離れ易く、他の分子へ移ると同時にエネルギーが発生して、エネルギーを必要とする生合成反応を進める。
一例をあげれば、ATP分解酵素の働きによってATPが加水分解すると、ひとつのリン酸基(P)がはずれてADP(アデノシン二リン酸)になり、その際にエネルギーを放出する。このエネルギーを使って筋の収縮が行われる。 筋繊維の中に蓄えられているATPの量は僅かなので、激しい運動では短時間で使い果たしてしまう。したがって長時間運動を続けるには、ADPからATPを再合成してATPを供給し続けなければならない。ATPの加水分解では、細胞内で様々な過程を進行させるエネルギーを供給する、この両者の反応によりATPが循環する。
目次へ