葉緑体と光合成
Top 車山高原 車山の山菜 車山日記 車山ブログ 車山高原の野鳥 車山7月日記 車山8月日記

☆早春のスミレ ☆車山高原の笹
☆車山高原のシジュウカラ
☆諏訪の植生 ☆諏訪に生息する哺乳類
☆諏訪の狐 ☆車山高原の狐
☆DNA ☆生物進化と光合成
☆葉緑素とATP ☆植物の葉の機能
☆植物の色素
| 目次 |
| 1)葉緑体の構造と光合成 |
| 2)光合成反応の詳細 |
| 3)光化学系チラコイド反応 |
| 4)葉緑体の構造と窒素同化 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成
花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1)
遺伝子の発現(2) 遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝
減数分裂と受精 対立遺伝子と点変異 疾患とSNP 癌変異の集積 癌細胞の転移 大腸癌 細胞の生命化学
イオン結合 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 シグナル伝達 キク科植物
陸上植物の誕生 植物の進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応
根による水吸収 稲・生命体 胞子体の発生 花粉の形成 雌ずい群 花粉管の先端成長 自殖と他殖
フキノトウ アポミクシス 生物間相互作用 バラ科 ナシ属 蜜蜂 ブドウ科 イネ科植物 細胞化学
1)葉緑体の構造と光合成
 光合成を行なう植物の葉緑体は、タンパク質・脂質・色素(クロロフィルとカロテノイド)・核酸(DNAとRNA)・アミノ酸・リン酸化合物・イオン・糖類などで構成されている。
光合成を行なう植物の葉緑体は、タンパク質・脂質・色素(クロロフィルとカロテノイド)・核酸(DNAとRNA)・アミノ酸・リン酸化合物・イオン・糖類などで構成されている。核酸は、生物の細胞核中に多く含まれる、塩基・糖・燐酸からなる高分子物質である。デオキシリボースにより構成されるデオキシリボ核酸(DNAと略される)と、リボースによるリボ核酸(RNAと略される)とに大別される。
DNAは細胞核の染色体として存在し、遺伝子の中核となる。ストロマには独自の葉緑体DNAが含まれ、それと対応して独自のリボソームがここに含まれている。RNAは、DNAとともに遺伝子を発現するために貢献する多様な種類のタンパク質の合成に関与する。
脂溶性の脂質や色素の大部分とタンパク質の一部は、葉緑体を包む2重の包膜やチラコイド膜を構成する。その他の水溶性の成分の大部分は、ストロマ・外包膜と内包膜の膜間腔(まくかんくう;まくかんこう)に存在する。チラコイド膜で囲まれた内側の部分はチラコイド内腔(ルーメン)ともいう。
光合成は非常に複雑な反応で、大きく2つの段階に分けて理解される。前半が「光(ひかり)エネルギー変換反応」で、光エネルギーを化学エネルギーに変換する生化学反応を起こさせる。
光エネルギーとは、電磁波の一種である光がもつエネルギーを言う。光エネルギーは光に含まれる光子の数と光子の周波数によって決まる。
「光エネルギー変換反応」は、チラコイド膜の内部で起こることから「チラコイド反応」とも呼ばれる。
「化学エネルギー」とは、化学物質内部に蓄えられたエネルギーのことで、物質を燃焼させると光や熱が発生するのは、物質内の「化学エネルギー」が光や熱エネルギーに変換されるからである。光エネルギーを元手に「化学エネルギー」を作り出す。それが「チラコイド反応(光エネルギー変換反応)」で起きている。
光合成の後半部分が、二酸化炭素(CO2)を還元してブドウ糖(C6H12O6) を合成し、植物内に二酸化炭素を固定する「炭素同化反応」である。この反応は、葉緑体のストロマで起きることから「ストロマ反応」と呼ばれ、これには「チラコイド反応」で作られた「化学エネルギー」が使われる。
この2つの反応は連動して起こるが、「チラコイド反応」は光エネルギーに依存するが、「ストロマ反応」は光エネルギーを直接的には利用しない。
チラコイドの膜上には、「チラコイド反応」を司るいくつかのタンパク質複合体が埋め込まれている。この中の2つは光エネルギーを受け取る「光化学系(PS)」というタンパク質複合体である。タンパク質複合体は、タンパク質と様々な分子が結合した物質である。「光化学系(PS)」には、「光化学系Ⅰ」と「光化学系Ⅱ」の2種類があり、それぞれがいくつものタンパク質や分子と結合している。反応の順番は、「光化学系Ⅱ」が先に働き、その後に「光化学系Ⅰ」が作動する。
光合成は、二酸化炭素(CO2)を還元してブドウ糖(C6H12O6)を合成する反応だ。「還元」とは物質が電子を受け取る反応で、二酸化炭素は還元されて電子を受け取り、それによりブドウ糖が合成される。ちなみに「酸化」は、「物質が電子を失うこと」であるから、「酸化」と「還元」という2つの反応は、必ず同時に起きる。ある物質が「酸化」されて失ったその電子が、別の物質に受け渡されるため、それを受け入れた物質が「還元」されたことになる。
光合成では、炭素一つで構成される二酸化炭素(CO2)分子6個が、光合成反応の結果、6つの炭素が結合したブドウ糖(C6H12O6)1個に変換される。つまりブドウ糖(C6H12O6)は、原子の数では「6(CH2O)
」と表示できる。光合成は、6つある二酸化炭素(CO2)の酸素原子(O)の1つが2つの水素原子(H)に置き換わった反応と言える。光合成は、二酸化炭素(CO2)を還元してブドウ糖(C6H12O6)を合成する反応であった。
「光化学系(PS)」は光のエネルギーを電子のエネルギーに変換し、「チラコイド反応」全体で電子を与える力、すなわち「還元力」を生み出す原動力となっている。
目次へ
植物の光合成反応は、空気中の二酸化炭素を吸収し、それを還元して、安全な形の有機物、すなわちブドウ糖(C6H12O6)を合成する唯一の自然化学反応である。
葉の中には細胞があり、その細胞の中を顕微鏡で見ると緑色の粒が見られる。長径10μm以下と小さい。それが葉緑体である。光合成はその緑色の粒の中で起こる。
葉緑体は2重の膜で囲まれている。その内側の内包膜は、共生したシアノバクテリアの細胞膜であった。その外側の外包膜は、シアノバクテリアを取り込むために窪ませた宿主の細胞膜と考えられる。内包膜と外包膜の間にある膜間腔は、10~20nm程度とごく狭く、ミトコンドリアの膜間腔とは異なり、その機能は不明である。
植物の光合成反応は、植物細胞内にある葉緑体の中で行われている。シアノバクテリアの細胞膜であった内膜の内部には、扁平な形状をしたチラコイド膜がぎっしりと積み重なったようになっている。その膜そのものに、クロロフィルやカロテノイドを含んだ光合成に関わるタンパク質が多く存在する。これらのタンパク質は、環境の変化に応じて互いに結合する相手を変えながら光合成反応の調節を行い、光エネルギーを化学エネルギーに変換する光反応( ひかりはんのう)の場となっている。
光合成反応を調節している光化学系タンパク質と太陽からの光エネルギーを集める「光化学系Ⅱ」の集光アンテナタンパク質、すなわち「アンテナ複合体LHG」がチラコイド膜に存在し、光合成色素の働きにより、膜内でその配置や互いの結合を変えることで、植物は光環境変化に適応しつつ太陽の光エネルギーを効果的に捕える。
すると光合成色素の分子に含まれる電子が、光エネルギーを受け取り活性化する。これを「励起状態(れいきじょうたい)」という。励起状態化した光エネルギーは、「LHG」の分子を媒介として伝搬され、最終的には「反応中心複合体」の「反応中心クロロフィルa」にまで受け渡される。そのクロロフィルaが吸収できない波長の光を集めるために、カロテノイド(橙色)・フィコエリトロビリン(赤色)・フィコシアノビリン(青色)などの別の光合成色素が使われる。光合成色素は葉緑素だけでなく、黄金アカシヤのカロテノイドや赤シソのフィコエリトロビリン・藻類に含まれる青色の色素タンパク質フィコシアノビリンも、光合成の主要な集光色素とし機能している。
可視光線は、電磁波のうち、ヒトの目で見える波長のもの、いわゆる光のことである。可視光線に相当する電磁波の波長の下界はおおよそ360-400 nm、上界はおおよそ760-830 nmである。可視光線より波長が短くなっても長くなっても、ヒトの目には見えなくなる。可視光線より波長の短いものを紫外線、長いものを赤外線と呼ぶ。「可視光」もエネルギーを持っている。それが光のエネルギーで、葉緑体が吸収する「赤色光」と「青色光」が、光合成を作動させるエネルギー源となっている。葉が緑色に見えるのは、葉緑体が主に緑以外の「赤色光」と「青色光」を吸収して光合成を行なうからである。
コケシダ・種子植物の葉の表面での吸収率は、クロロフィルa(青緑)やクロロフィルb(黄緑)、カロテノイドのカロテン(橙)とキサントフィルのルテイン(黄)などでは、赤色光や青色光帯に対する吸収率は、緑色光よりも高い。赤色光と青色光の葉の表面での吸収率は90%位、緑色光の表面での吸収率は60%ぐらいだという。
葉の表面で吸収されなかった緑色光は、葉裏の海綿状組織で乱反射を繰り返す中に、緑色光は70%~80%ほどませ吸収される。葉が緑色に見えるのは、赤色光や青色光も反射していますが、緑色光が青色光や赤色光より多く反射するからだ。我々には緑色の光が目に入るので、葉は緑だ、と認識する。
光を吸収しているのは、葉緑体の中に在る「光合成色素」と呼ばれる物質である。クロロフィルa(青緑)やクロロフィルb(黄緑)、カロテノイドのカロテン(橙)とキサントフィルのルテイン(黄)などが、葉緑体内の「アンテナ複合体LHC」と結合し、光のエネルギーを受け取る。
「光合成色素」いずれもが、緑周辺の波長を吸収していない。光合成色素の内、葉緑体にもっとも多く含まれるのが「クロロフィル」、それにはいくつかの種類があり、光合成生物のほぼすべてに共通して見られる「クロロフィルa」は、光合成反応で特別な役割を果たしている。
目次へ
光合成の前半部で起こる「チラコイド反応」を司るいくつかの分子が、チラコイドの膜上に埋め込まれている。その中の2つが、光のエネルギーを受け取る「光化学系(PS)」というタンパク質複合体だ。「光化学系(PS)」には「光化学系Ⅰ」と「光化学系Ⅱ」の2つがあり、反応の順番は「光化学系Ⅱ」が先に作用して、その後に「光化学系Ⅰ」が作動する。
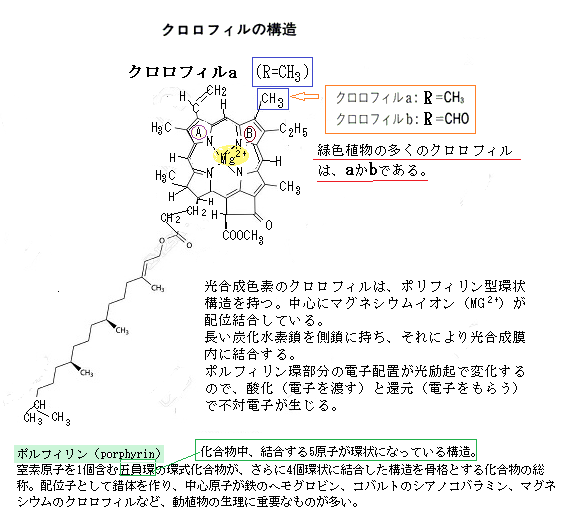 光エネルギーの反応を別の視点からみると、「アンテナ複合体LHC」に結合している様々な光合成色素が、幅広い波長の光を集め、その光のエネルギーを、光合成反応にスイッチを入れる「反応中心複合体」に受け渡しをする。その「反応中心複合体」の主要部分が「反応中心」と呼ばれる「クロロフィルa」なのだ。最終的に「クロロフィルa」が光のエネルギーを受け取る。陸上植物では、「クロロフィルa(青緑)」と「クロロフィルb(黄緑)」の割合が約3:1と知られている
光エネルギーの反応を別の視点からみると、「アンテナ複合体LHC」に結合している様々な光合成色素が、幅広い波長の光を集め、その光のエネルギーを、光合成反応にスイッチを入れる「反応中心複合体」に受け渡しをする。その「反応中心複合体」の主要部分が「反応中心」と呼ばれる「クロロフィルa」なのだ。最終的に「クロロフィルa」が光のエネルギーを受け取る。陸上植物では、「クロロフィルa(青緑)」と「クロロフィルb(黄緑)」の割合が約3:1と知られているそのため「クロロフィルa」は「主色素」と呼ばれ、その他は「補助色素」と呼ばれることがある。
「反応中心クロロフィルa」という特殊な状態のクロロフィルは、クロロフィルが2分子で1つの反応中心を作っている。この「反応中心クロロフィルa」に光エネルギーを集中的に集めている。多くの場合、他のクロロフィルから、吸収した光エネルギーをもらうが、「反応中心クロロフィルa」は、自らも直接、光エネルギーを吸収する。そのため反応中心ではないクロロフィルが、光エネルギーを集める役割に変化が生じた。
チラコイドの膜上における反応を詳細に語れば、「光化学系Ⅱ」の「アンテナ複合体LHC」が、光エネルギーを集める役割の「アンテナクロロフィル」の働きにより太陽のエネルギーを捕える。
その「アンテナクロロフィル」が光エネルギーを受けて活性化し、その「アンテナクロロフィル」に含まれる電子が励起状態となり、その電子を他の物質に与える還元力を持つ。しかも、その光エネルギーは「アンテナ複合体LHC」の分子を媒介として伝搬され、最終的には「光化学系Ⅱ」における光合成反応にスイッチを入れる「反応中心複合体」のまさに「反応中心クロロフィルa」にまで受け渡される。
結果、「光化学系Ⅱ」の「反応中心クロロフィルa」の電子が励起状態となり、その電子を他の物質に与える還元力を持つようになる。その電子を受け取るのが「電子伝達鎖」と呼ばれる分子群で、受け取った電子は「光化学系Ⅰ」に受け渡される。その電子を「光化学Ⅰ」に受け渡したときに酸化され、再び電子を受け取りやすい状態に戻る。
この「電子伝達鎖」は、もともと電子を受け取りやすい状態にあり、「光化学系Ⅱ」から電子を受け取った時に還元され、その電子を「光化学系Ⅰ」に受け渡した時に酸化され、再び電子を受け取りやすい状態に戻る。
「光化学系Ⅰ」でも「光化学系Ⅱ」とよく似た反応をする。「アンテナ複合体LHG」に還元され、「アンテナ複合体LHG」が捕えた光のエネルギーが、「反応中心複合体」にある「反応中心クロロフィルa 」に伝わると、光のエネルギーによって励起状態となり、その「反応中心クロロフィルa」の電子が飛び出す。
この時に放出された電子は、「光化学Ⅰ」に結合した物資の間を次々と移動し最終的に別のタンパク質に渡される。最終的に受け取るのが、「NADP/ NADPH」と言う物質で、この物質は電子の受け渡しにより「NADP」と「NADPH」という2つの状態を行き来する。それでこの不可解な表記になった。「NADP」が電子を受け取る(還元)時に水素イオン(H+)と結合し「NADPH」へと変わり、反対に電子を失しない(酸化)水素イオン(H+)が分離し「NADP」に戻る。
還元される「NADP」は電子を受け取る能力がある。即ち相手から電子を奪う「酸化力」を持った「酸化剤」となっていた。また「NADPH」は相手に電子を与える「還元力」があり「還元剤」となっていた。
「チラコイド反応」の最後に「光化学Ⅰ」から電子を還元され、水素イオン(H+)と結合した「NADPH」は、このあとの「ストロマ反応」で重要な役割を果たす。二酸化炭素(CO2)に電子を与える「還元力」を持った物質としてブドウ糖(C6H12O6) を作る原動力の一つとなる。この「NADPH」の「還元力」が、「チラコイド反応」によって作られる「化学エネルギー」の一つとなる。
光合成は、光や温度、二酸化炭素濃度が一定の条件を満たした場合に、連続して起こる反応である。植物は光のエネルギーを活用し、通常では還元ができない物質に電子伝達(酸化還元)作業を繰り返し、その生命の存続を託したようだ。「光化学Ⅰ」の「反応中心クロロフィルa 」は、光のエネルギーをもらうと一時的に強い還元剤となって、通常では還元できない相手に電子を渡すことができるようになる。言い換えれば自発的に進行しない電子伝達(酸化還元)反応が、光のエネルギーによって進行する。
しかもやがて一連の作業が終わり、何もしなければ、次には逆の反応が生じ、還元した電子の酸化作用が復活し反応中心に戻る可能性がある。これを避けるために、電子を受け取った物質の反応は速く別の物質に渡されると、それが更に別の物質に渡される、という仕組みが作られている。この電子の伝達によって、利用できる自由な光エネルギーは次第に減少するため、無駄にエネルギーを消耗させないよう反応を一方向に進めて行く。それは生物全体にみられる極めて重要な生存の摂理となっている。
一方、電子を渡した後の「光化学Ⅰ」の「反応中心クロロフィルa 」は、他の「アンテナ複合体LHG」から電子を受け取り元の状態に戻る。生体内で還元剤として使われる「NADPH」を作るため、それよりも、もっと強い還元剤を光のエネルギーを使って作るのが「光化学Ⅰ」の役割だ。
目次へ
脂質二重膜で存在するチラコイド膜は、膜が1枚ずつ独立した形状をしている場合と複数の膜が重なり多重になっている場合がある。一般的にコケ植物も含め陸上植物では、1枚の胞膜で存在する「ストロマラメラ」と、所々に10枚程で積み重なった「グラナ」呼ばれる積み重なる構造で散在している。「ストロマラメラ」は重なりのない1枚の胞膜として存在し、そのストロマラメラは複数の「グラナ」を繋ぐような形で屈折を繰り返す1本の道筋となる物質で連結されている。それ以外には、大きな構造もなく、グラナの間隙をストロマが満たす。そのストロマには独自の葉緑体DNAが含まれ、それと対応して独自のリボソーム(Ribosome)もここに含まれている。
リボソームは、動植物の細胞内の双方にあり、生物には重要にして不可欠な機能を果たしている。葉緑体のリボソームRNA(rRNA)の塩基配列は、シアノバクテリアの中にも含まれている。リボソームは、mRNA(messenger RNA)の遺伝情報を読み取ってタンパク質を合成するという場である。
葉緑体を生じさせる細胞内共生が1回だけ起きたという進化が、その後の地球の運命を変えた。次に、植物や二次共生藻類の起源が、シアノバクテリアを細胞のなかに取り込んで葉緑体と共生する進化過程を経たことに始まる。
葉緑体やミトコンドリアは他の細胞器官と異なって、それぞれが半自律的に分裂によって増殖し、しかも独自の遺伝子を持っていることが知られている。葉緑体自身がDNAを持っている。リボソームはRNAとタンパク質からなる巨大な複合体である。細胞はリボソームの生成に多大なエネルギーを割いている。真核生物ではリボソームの構造がより複雑になっている。それを元に蛋白質合成をするためのリボソームRNA(rRNA)の塩基配列は、mRNAの遺伝情報を読み取ってタンパク質を合成する構造体を葉緑体は独自に備えている。
植物は光合成によって、大気の主成分である二酸化炭素から作られる炭水化物だけでは生きられない。生物の細胞は、炭水化物に加えてタンパク質(アミノ酸の結合体)や核酸(DNA・RNA)、及び脂質(糖質と並ぶ生物のエネルギー源)など様々な有機物と、水や多種類の無機元素からできている。生物の生体内には、多種類の微量元素が存在する。それが酵素の触媒として、様々な役割を果たしている。その多種類の微量元素の成分が、生物そのものの維持に重要な役割を果たしている。「クロロフィルa」の分子構造には、マグネシウム(Mg)が配位されている。
葉緑体は光合成以外に、根から取り込んだ窒素分をアンモニアに変え、これを素材にして、生体を構成するアミノ酸やタンパク質などの有機窒素化合物を合成する窒素同化の反応の場としての役割を担う。
窒素は核酸やタンパク質を構成する重要な原子でありながら、大気中もっとも多い窒素源は、大気中で4/5を占めているも、その分子状の窒素(N2)は、科学的に不活発で多くの生物には利用できない。
植物が窒素同化作用の材料として利用できる窒素源は、無機窒素化合物と分子状の窒素(N2)の2群になるため、高等植物は、土壌中のアンモニアと硝酸イオンを窒素源として成体に取り込み、アミノ酸や核酸などの有機物に変化させる。畑では、肥料とした散布されたアンモニアは、バクテリアによる「硝化作用」によって亜硝酸イオンを経て硝酸イオンに酸化される。アンモニアと硝酸イオンしか窒素源として言及されないのは、それら以外の窒素源は環境中にあまり存在しないからという。
同様にイオウ分をアミノ酸に取り込むイオウ同化、さらに生体膜の成分として重要な脂肪酸の合成も葉緑体が担う。
目次へ