| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環
| 目次 |
| 1)植物の能力 |
| 2)維管束組織 |
| 3)植物の組織分化 (根圧)と(木部) |
| 4)管状要素(trachearry element)trachearry=管状の、仮導管の |
植物は、胚から発生する多細胞生物であり、二酸化炭素から有機物を光合成により作り出し(独立栄養性)、一般的に固着性であるため陸上環境に適した生物と言える。この植物の定義では、蘚類(せんるい:スギゴケ・ミズゴ・ギンゴケなど)から顕花植物の系統までが含まれる。
光合成を行う重要な特性は、植物とその殆どが水棲である緑藻の間で共有されている。ただ、陸上植物に共有されている、陸上で生きるために獲得された形質は、藻類にはないが、どちらの系統の葉緑体にも、光合成色素としてクロロフィルaとクロロフィルb、及びβ-カロチン(カボチャ、ニンジンなど、色鮮やかな緑黄色野菜などに多く含まれるカロテノイドの一種)が含まれている。
植物と緑藻は、究極の集光装置を備え、光合成により生体としての有機化合物を合成し増殖していく独立栄養生物であり、しかも地球上随一の一次生産者である。いずれも太陽の光エネルギーを利用して、水と二酸化炭素から炭水化合物を合成する。それが光エネルギーを化学結合させ化学エネルギーに変換して蓄積し、水と大気中の二酸化炭素から生合成した炭水化合物から、多糖・タンパク質・核酸・脂質など巨大分子が構成される。
 植物は、水や無機養分を土壌から光合成器官や成長中の器官へと輸送する仕組みと、光合成器官から非光合成器官へ光合成産物を輸送する両方の仕組みを持つ。
植物は、水や無機養分を土壌から光合成器官や成長中の器官へと輸送する仕組みと、光合成器官から非光合成器官へ光合成産物を輸送する両方の仕組みを持つ。その役割を担う維管束組織系が、植物の各部分を繋ぐ輸送網を作る。篩部は葉で作られた光合成産物を根に運ぶ。光照射下で光合成された蓄積代謝産物を、植物成長のエネルギー源とする。
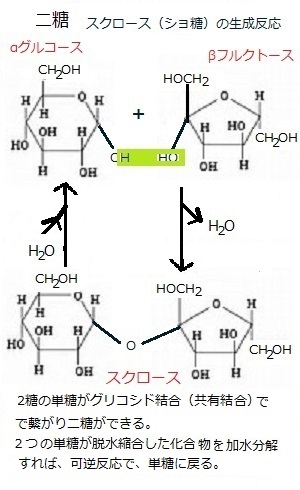
多くの植物におけるCO2の光合成同化過程では、サイトゾルcytosol(細胞質ゾル;細胞質のコロイド状の溶質で、細胞質からリボソームなどの細胞内小器官や、細胞内外の運動に必要な物理的力を発生させる繊維状構造の細胞骨格を除いた部分)でスクロースsucrose(グルコース=ブドウ糖とフルクトース=果糖がC-1,2-グリコシド結合した二糖類)が、葉緑体中でデンプンが作られる。
日中に、スクロースは、葉のサイトゾルから様々なシンク組織に転流する一方、デンプンは葉緑体中に、高濃度なデンプン粒として蓄積される。夜になるとCO2同化は止まり、葉緑体に蓄積されたデンプンの分解が始まる。
葉緑体中のデンプン量には、夜間を通して減少し、その分解産物はサイトゾルへ流れる。この篩部の組織は成熟しても、木部と違い細胞壁がセルロースとともに、木材の構成成分としてリグニンlignin化されることはない。
Ligninとは、「木材」を意味するラテン語 lignum を原語とする。セルロースとともに木材を構成する重要な高分子化合物である。 水や無機栄養を根から運ぶ木部を構成する細胞群を管状要素と言う。すべての維管束細胞に見られる仮導管と、主に被子植物に見られる比較的短い導管要素の2種類がある。
仮導管は、被子植物・裸子植物・シダ類や他の維管束植物に存在し、導管要素は、裸子植物とグネツム目と呼ばれる裸子植物中の1グループと、いくつかのシダ植物のみに見られる。
仮導管と導管要素の成熟には、二次細胞壁の形成と、それに続く細胞質やすべての内容物を失う細胞死が関与する。残るのは、厚い木化した細胞壁だけで、中空の管となり、その中を比較的小さい抵抗で水が流れる。
死細胞となる。導管要素は、両端で繋がり、最大で0.7mmほどの幅を持つ円柱状の導管を形成する。一次細胞壁のみを持つ原生木部細胞が管状要素へと成熟する過程で、セルロースが螺旋状に沈着し、疎水高分子のリグニンにより強化され二次細胞壁ができ、その細胞壁に堆積して木質化を起こし、植物体を強固にする。
目次へ
 殆どの植物では、木部が水輸送の経路をほぼ独占している。1mの高さの木では、その植物体の水輸送経路の99.5%以上を占めている。高木となると、木部の水輸送経路は、より高い割合となる。生きた細胞層を通した水の輸送と比べ、木部はより抵抗の少ない単純な経路で貢献している。
殆どの植物では、木部が水輸送の経路をほぼ独占している。1mの高さの木では、その植物体の水輸送経路の99.5%以上を占めている。高木となると、木部の水輸送経路は、より高い割合となる。生きた細胞層を通した水の輸送と比べ、木部はより抵抗の少ない単純な経路で貢献している。水中で進化した植物が、陸上で生存するには、いくつかの切実な課題を克服しなければならない。特に水の獲得と保持である。その陸上環境に対応するため、植物は根と葉を進化させた。根は植物を固定し、水と栄養(ミネラル)を吸収する。葉は光を受けてガス交換をする。植物が大型化すると、根と葉の距離はますます遠くなる。このため根と地上部の間では、根で吸収した水や栄養を葉に送り、葉で合成された光合成産物を根に送る効率的な長距離輸送システムを進化させた。
木部は、植物の地上部分に根系からの水やミネラルを輸送する組織であるが、篩部は根を含む成長している組織や貯蔵組織に、成熟葉から、光合成産物特に糖を輸送する組織である。篩部はまた、食物ホルモンやタンパク質などの制御分子の輸送を介してソースとシンクとの間で信号を伝達し、植物体全体への水及び種々の化合物の再配分を行う。これらの分子のすべてが、篩部を通して輸送される糖と共通の働きをする。篩管を介して再配分される化合物には、木部を通してはじめて成熟葉に運ばれるもの、修飾されずに成熟葉からそのまま再配分されるもの、再配分の前に代謝返還を受けるものなどがある。
篩管液は、「上向き」または「下向き」にだけ輸送されるのではなく、また、その輸送方向は重力に従うものでもない。篩管液は、糖のソースと呼ばれる供給源から、シンクと呼ばれる代謝や貯蔵の場へ輸送される。
ソース(source)は、糖を組織外へ輸送するすべての器官をさし、多くの場合、自身が必要とする以上の光合成産物の合成能をもつ成熟葉を指す。光合成産物という用語は、光合成により同化され生成物を言う。もう一つのソース組織の例に、貯蔵物質を他の器官へ輸送する貯蔵組織がある。たとえば、二年生のテンサイ(砂糖大根)の野生種の貯蔵根は、一年目にはシンク(sink)としてソース葉から輸送されてくる糖を貯蔵する。2年目の成長期には、同じ貯蔵根が今度はソースとして機能する。糖は地上部へ輸送されて新しい茎の成長に使われ、その茎は生殖成長を行うようになる。
シンクは、植物のすべての非光合成組織と自らの成長や貯蔵に必要な光合成産物を生み出さない光合成組織である。根・塊茎・成長中の果実、および未成熟葉は、正常な発達に炭水化物の供給が必要であるから、すべてがシンク組織である。
篩部輸送は、単純にソースからシンクへの輸送であると言えるが、特定の輸送経路はそれほど単純ではなく、シンク組織とソース組織の距離、発展段階や維管束の繋がり方、輸送経路の変化などにより左右される。すべてのソース組織が、すべてのシンク組織へ糖を供給するわけでなく、特定のソースから、特定のシンクへ供給される傾向がある。
目次へ
 植物は、母細胞から供給される養分を使い胚から発生する。この養分を胚に蓄積できることで、陸上で大型化する事が可能になった。「胚発生」は、受精卵と言う単細胞から、ある特徴を持ちながら、基本的体制を維持した一つの多細胞体に変化する過程を表す。
植物は、母細胞から供給される養分を使い胚から発生する。この養分を胚に蓄積できることで、陸上で大型化する事が可能になった。「胚発生」は、受精卵と言う単細胞から、ある特徴を持ちながら、基本的体制を維持した一つの多細胞体に変化する過程を表す。種子植物の「胚発生」は、1個の細胞の受精卵(接合子)から、成熟した種子の中に、かなり複雑な個体を作り出す。そのため胚発生には、多くの発生過程上の違いが生じ、植物それぞれに特異な基本的な構造を形成する。
これらの過程には、形を作る「形態形成」、機能的に組織化された構造体を作る「器官形成」、細胞が分化して機能的に明確な組織となる「組織形成」などがある。こうした基本構造に欠かせないのが、シュート(shoot;茎と節からなる部分)と根の頂端に生じる頂端分裂組織である。これらが無限とも言える栄養成長を支えていく。その胚発生の最終過程に、胚を長期にわたって不活性化する複雑な生理学的変化「休眠」と、その後の環境情報をシグナルとして反応し、成熟胚が再び活性化する過程、「発芽」を迎える。
植物は、特定の生殖細胞を除き、固着性で移動することはない。その代り、一生、生存に不可欠な光・水・無機養分などの供給源をターゲットとし成長する。それに伴い進化をする。
向日性とは、太陽によって誘導される葉の運動である。太陽光の軌道に添い、葉の受光を最大化することを「正向日性」と呼ぶ。一部の種では、葉は強光を避けるように動き、温度上昇や水の損失を最小限にする。このように葉が太陽光を避けることを「負向日性」と呼ぶ。大豆などの一部の植物は、水が十分にある環境では、「正向日性」となり、渇水ストレスが負荷されると「負向日性」となる。植物は、環境変化に対応して、能動的に動いている。
固着性の植物が、どのようにして無機養分を収集し、利用するかについての研究領域を「無機栄養」と呼ぶ。無機養分とは、窒素・リン・カリウムなどの元素で、植物は無機イオンの形態で、それらを土壌から獲得している。無機養分は、すべての生物の間を循環しているが、その大部分は、植物の根系から吸収され生物圏に入ってくる。根はその大きな表面積で、土壌水中の低濃度の無機イオンを吸収する能力を持つため、植物は無機養分を効率よく吸収できる。根から吸収された無機養分は、植物の異なる部位に運ばれ、多くの生物反応に関わる。菌根菌や窒素固定細菌のような生物が、根とともに無機養分の獲得に関わることも多い。
根の表面と土壌の接触は、根による効率的な水の吸収には不可欠で、その接触による水吸収にともない根や根毛は成長しながら最大化する。根毛(root hair)は、表皮細胞が糸状に派生したもので、根の表面積を極限にまで増やし、土壌からのイオンや水の吸収能力を格段に高める。播種後3ヵ月の小麦は、根毛が根の表面積の60%以上を占めていたという。
目次へ
木部における溶質の蓄積が根圧を作り出す。もし若い芽生えの茎を土壌から少し出たところを切断すると、何時間にもわたって切り口の木部から液を排出することが多い。そのため、かなりの陽圧(物体の内部の圧力が外部より高い状態)が測定できるという。その地上部の切口、あるいは幹にあけた穴に取り付けた根圧計で測定すると、時には0.2MPa(メガパスカル)より高い値にも達すると言う。2~5気圧ともいわれるが、一般には格段に低い。
根圧は、葉による蒸散流が完成するまでの一時的な水の上昇に関与する補助的な役割を果たすものと考えられている。根の代謝活性に左右されるので、根圧の発生は能動的な働きとみなされている。
蒸散が低いか、あるいはない場合、根は土壌からイオンを吸収し、それを木部に輸送し続けるので、木部には正の静水圧(密閉空閑内に水を押し込むときに発生する圧力)が形成される。木部液中の溶質の蓄積は、木部の浸透ポテンシャルを低下させ、その結果、この木部のポテンシャルの低下が水吸収の駆動力となり、今度は、木部中に陽圧を発生させる。
植物を含むすべての生物は、成長や生殖はもとより、高度に組織化された構造を保ちながら修復するため、絶え自由エネルギーを必要とする。生化学反応・溶質の蓄積・長距離輸送などの過程は、全て自由エネルギーの入力により駆動される。
水の化学ポテンシャルは、水の自由エネルギー状態を示す。熱力学では、自由エネルギーを、仕事(力 × 距離)をする能力(potential)として表す。
化学ポテンシャルは、相対値に過ぎない。化学ポテンシャルとは、ある状態における物質の化学ポテンシャルと、同じ物質の標準状態における化学ポテンシャルの差で表される。水の化学ポテンシャルも、水に関する自由エネルギーで表される。それで、水は、化学ポテンシャルが高い所から、低い所へ、エネルギーの入力を加えることなく自発的に流れる。植物の水ポテンシャルに影響を与える主な要素は、「濃度」・「圧力」そして「重力」である。
溶質ポテンシャルあるいは浸透ポテンシャルと呼ばれるのは、水ポテンシャルに対する溶質の効果を表す。むしろ、溶質は水を薄めることなり自由エネルギーを減少させる。則ち、溶質と水の混合は、その系の無秩序さとエントロピーを増加させるため、自由エネルギーを減少させることになる。溶質ポテンシャルでは、溶質自体の固有の性質に、通常、左右されないことを意味する。
エントロピー増大の法則に従うすべての系は、溶質は溶けている溶媒全体に拡散する。溶質が移動できる範囲は、細胞には選択的透過性を持つ膜があるため、細胞内の溶質濃度が、細胞外の溶液よりも高ければ、外部の水は細胞内へ拡散するが、溶質は細胞外へ拡散しない。この選択的透過性を持つ膜を通過する水の正味の移動を「浸透」と呼ぶ。
ところが、選択的透過性を持つ膜が、自由に拡散する溶質を、破裂するまで受け入れ程、細胞は膨張し続けるだろうか。細胞壁を持たない動物細胞では起こるが、極めて強い細胞壁を持つ植物細胞は、水が細胞内に流入し細胞壁に圧力がかかれば、細胞壁は変形に対して大きな抵抗を示して、水の流入を細胞壁の圧力によって、細胞の体積の増大を物理的に抑制する。これが細胞内の「静水圧」の上昇なのである。
実質的には、多細胞系からなる根の組織は、まるで浸透膜であるかのように、溶質の蓄積によって木部中に陽圧を生じさせる。
気体と違い、液体や固体が凝集して一定の体積をもつのは、分子・原子またはイオン間に引力が働くからで、この力を凝集力という。しかし、凝集力だけに頼れば、固体や液体はその体積を縮めるだけである。そのため、短い距離間では反発力が作用している。一般に,凝集力は反発力に比べると遠くまで作用する。凝集力には、分子間の静電引力やファンデルワールス力、水分子同士の水素結合やイオン間で働く電荷移動力などがあげられる。
根圧は、土壌の水ポテンシャルが高く、蒸散速度が小さいときに、最も生じやすい。蒸散速度が増すにつれ、水は植物を通して輸送されて素早く大気へ失われるので、イオンの取り込みにより、溶質濃度の高まりに伴い発生する陽圧は、木部中では生じない。
根圧を生じるような植物は、葉の縁に水滴を付けることが多い。それが排水という現象で、木部の陽圧は、葉の縁の葉脈の末端に繋がる排水組織と呼ばれる特殊化した孔を通して木部液を排出する。
朝、草の葉の先端に見られる「露滴」は、実際はその排水組織から排出された液である。排水は、夜のように蒸散が低下し、相対湿度が高いとき最もよく見られる。
根圧は、髙いイオン蓄積速度によって生じる不可避な結果を反映している。夜の木部内での陽圧は、気泡の溶解を助け、それによりキャビテーション(cavitation)の有害な作用が打ち消される。Cavitationとは、空洞化、空洞形成の意味であり、「道管内の小さな気泡が大きく膨張することにより起こる水柱の張力の崩壊」を言う。
目次へ
植物は、重力に逆らい太陽に向かって成長する。そのため自分自身の重量を支えるための頑丈な構造を持つ。植物細胞は、動物細胞と異なり丈夫な細胞壁で支えられている。この薄い層は、足場となるセルロースcellulose微繊維と、その間隙を埋めるマトリックス(matrix)である多糖類や糖タンパク質(タンパク質を構成するアミノ酸の側鎖に糖鎖が共有結合しているもの)とその他の高分子化合物からなる。
マトリックス高分子とセルロース微繊維は、共有結合と非共有結合が混在した結合様式からなるネットワーク構造を形成している。マトリックには、細胞壁の物理的・化学的性質を修飾する酵素やその他の成分も含まれている。多くの細胞では、一次細胞壁の内側に、リグニン化した二次細胞壁が肥厚状態で沈着している。この状態が一番顕著な木部では、その細胞壁が長距離水輸送と茎の構造支持を担う。
セルロースは、細胞壁の主要な繊維状成分となる多糖で、規則性の高い領域と低い領域をもった微繊維を、細胞膜上で合成され、その水に不溶な疎水性の部分で高い引っ張り強度を持つ。最も単純なセルロース微繊維(cellulose microfibril)は、約3nm(1nm = 10-9m)幅の細い構造からなり、細胞壁に強度を与えている。微繊維の向きで細胞膜の強度が変わる。各微繊維は、C1とC4の炭素原子が隣り合う-β結合で直鎖状に繋がったグルコース鎖が、18~24本(18本が一番多い)、同じ方向に向いて密集して束になり、分岐の多いグルカン側鎖の分子間で広範囲なO-H-Oの水素結合をし、非常に規則性の高い芯構造を形成する。
グルカン(glucan)は、グルコース(glucose)単位が一列に連結したもので、糖から構成された高分子化合物の一般名称で、多糖と同義である。セルロースは、グルコース単位の環状分子が、上下前後逆になって繋がって直鎖状に並ぶ。
芯構造を取り巻く周囲のグルカン側鎖は、芯部よりも柔軟性があり、その配置は水分子や表層のマトリックス多糖類との相互作用に影響される。微繊維の表面には、親水性の部分と疎水性の部分があり、その親水面では積み重なったグルコース鎖の側面から伸びる極性の高い-OH基が密集して分布し、疎水表面ではグルコースの環状構造の面上に非極性のC-H基が密集している。
 ミトコンドリア内膜中に含まれるマトリックスは、液体分散媒(分散している物質の微細な粒子を分散相、分散相が入り込んだ気体や液体を分散媒という)のコロイド状の溶液(ゾル)であるが、植物細胞壁のマトリックスは、セルロース微繊維以外の多糖類で、一次細胞壁では、ペクチンとヘミセルロース及び糖タンパク質から構成される。
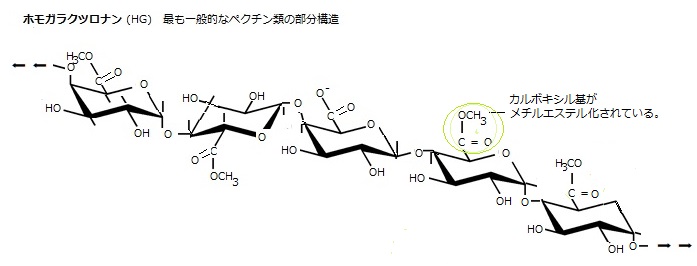
ミトコンドリア内膜中に含まれるマトリックスは、液体分散媒(分散している物質の微細な粒子を分散相、分散相が入り込んだ気体や液体を分散媒という)のコロイド状の溶液(ゾル)であるが、植物細胞壁のマトリックスは、セルロース微繊維以外の多糖類で、一次細胞壁では、ペクチンとヘミセルロース及び糖タンパク質から構成される。ペクチンは、ガラクツロン酸のような酸性糖とラムノース・ガラクトース・アラビノースなどの中性糖を含むものが典型である、中性複合多糖類などからなる複雑で多様なヘテロ多糖類で、細胞壁中ではゲル(コロイド溶液が流動性を失ってゼリー状となったもの)を構成し、セルロースとヘミセルロースからなる網状構造の内部を埋めるように存在している。
ペクチン類は、一次細胞壁の親水性のゲル形成成分である。殆どの一次細胞壁に最も含まれる成分で、水和したゲルを作り、その中にセルロースやヘミセルロースが埋め込まれている。ペクチン類は、親水性の充填剤として、セルロースの網状構造の凝集や崩壊を防ぐとともに、巨大分子が細胞壁内を移動する際の律速(反応の速さを決定する最も主な要因)を左右する細胞壁間隙の大きさを決定する。
ペクチン類は、特に中葉(middle lamella: lamellaとは薄板)に多く、3つの細胞が接する中葉では特段に重要性が増す。中葉は、隣り合う細胞が接する面の境界にある薄い層である。その主要な機能は、細胞間の柔軟な接着層となることであるが、例えば、カビ・キノコなど真菌のような病原体が、植物細胞に侵入すると、シグナル伝達物質としてペクチン由来のオリゴ糖が遊離する、それが防御応答を誘発し、病原体の侵入を抑制もする。
エクアトリアル配置とは、糖の環状構造が水平方向に結合する立体配置を言う。C1・C4-β結合とは、C1とC4の炭素原子間で隣り合うグルコースの環状構造が、β立体配置のグルコシド結合をしているという意味である。
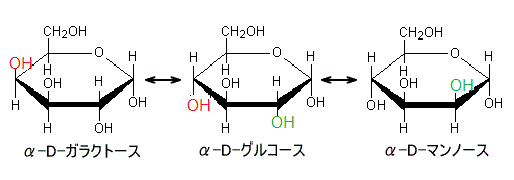
D-グルコースの、1位の炭素原子に結合する-OHと、6位の-CH2OHが環平面に対して反対側にあるものをα体、同じ側にあるものをβ体と呼ぶ。
2個の単糖がグルコシド結合(共有結合)で繋がって二糖ができる。この反応は、2個の分子が水分子1個を失って結合するもので、一般的な脱水縮合反応である。この逆反応が加水分解で、水が加わる。
ペクチンとヘミセルロースは、マトリックス多糖類とも呼ばれる。植物細胞壁は、足場となるセルロース微繊維と、それを埋める高分子性のマトリックスからなる。マトリックスは、植物種や細胞型、あるいは細胞壁内の部位により異なる。真正双子葉植物(eudicots)の典型的な一次細胞壁は、ペクチンを多く含み、セルロースやヘミセルロースは少ない。二次細胞壁では、セルロースとヘミセルロースを多く含み、そのヘミセルロースの分子種も一次細胞壁とは異なる。また二次細胞壁には、芳香族化合物の重合体であるリグニンを含み、その量は細胞壁の種類ごとに異なる。
一次細胞壁はペクチンを多く含むため、二次細胞壁よりも含水量が高くなり、細胞が拡大する際には、細胞壁の伸展力となる。二次細胞壁は、セルロース・ヘミセルロース・リグニンの構造が、密に押し詰められて、含水量が乏しくなるが、それが強度や圧縮耐性を高めることになる。
目次へ
水や無機塩類を根から運ぶ木部を構成する細胞群を管状要素と言う。全ての維管束植物に見られる仮道管(tracheid)と、主に被子植物に見られる比較的短い導管要素(vessel element)の2種類がある。いずれも成熟した段階で死細胞となる。導管要素は両端で繋がり、最大0.7mmほどの幅を持つ円柱状の導管(vessel)を作る。
一次細胞壁のみを持つ原生木部細胞が管状要素へ成熟する過程で、セルロースが螺旋状に沈着し、リグニングにより強化された二次細胞壁ができる。
細胞伸長が停止すると、細胞の長軸の上端と下端の細胞壁に大きな穿孔が作られる。細胞の側面では、壁孔以外の部分で二次細胞壁の肥厚化が進む。もともとは、隣り合った細胞間での情報交換を仲介し、タンパク質・核酸などの様々な巨大分子の径路として働いてきた原形質連絡(plasmodesma)が密集していた領域に形成された壁孔は、次第に隣り合った細胞を結ぶ細胞壁中の通路となった。原形質連絡は、多細胞植物体の細胞間の細胞壁を貫く細い細胞質の膜で、これによる細胞間の連絡通路をいう。
植物細胞は、原形質連絡を介して、互いに繋がった連続体を形成している。この連続体をシンプラスト(symplast)と呼ぶ。細胞質と通路の中心を通るデスモ小管と呼ばれる小胞体由来の膜が管を満たし、隣接する細胞間の小胞体と繋ぐ。シンプラストを通した細胞から細胞への分子の移動を可能にする。
太さ40~50㎚前後で、両細胞の細胞膜がこれを通じてつながって細い管をつくり、その中を細胞質が通っている。通路の隙間の大きさは、その内側表面に並んだ球状タンパク質によって制御されている。ウイルスと同じ大きさの粒子までは通すことができる。物質とくに高分子物質の流通に役だっていると考えられる。仮道管と導管要素は、プログラム細胞死により細胞死を起こす。これにより硬い二次細胞壁からなる、かつ周囲の細胞と壁孔と連絡した管の束が残される。
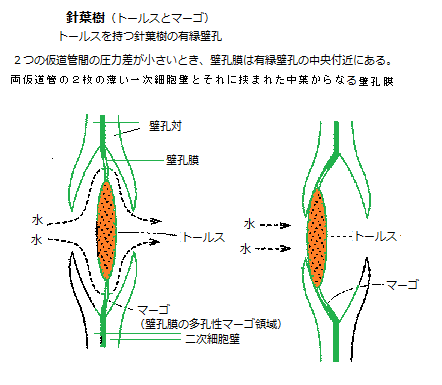 針葉樹では、仮道管の壁孔膜は中心が顕著に肥厚しており、トールス(torus)と呼ばれる。その周りには、多くの隙間を持ってトールスと上下の細胞壁を繋ぐセルロース微繊維構造のマルゴ(margo)がある。
針葉樹では、仮道管の壁孔膜は中心が顕著に肥厚しており、トールス(torus)と呼ばれる。その周りには、多くの隙間を持ってトールスと上下の細胞壁を繋ぐセルロース微繊維構造のマルゴ(margo)がある。壁孔膜の大きな直径と多くの隙間を持つマルゴは、多孔性で比較的柔軟で水を効率よく通すことができる。厚く肥厚したトールスは、バブルのように働き、壁孔対の空間の中央にあれば、壁孔は開孔している。
隣接した仮導管でキャビテーションが起きると、壁孔膜を挟んで圧力差が生じて、壁孔膜が水の入っている仮導管の側へ素早く移動する。その円形あるいは楕円形の厚くなった部分が壁孔に移動して、入り口を塞ぎ気泡の侵入を防ぐのだ。こうした有縁壁孔の構造は、通水の効率と、危険な気泡が隣の仮道管に広がるのを防ぐという安全性を同時に実現している。
ごく少数の例外はあるが、他の全ての植物の仮道管や導管要素の壁孔膜には、トールスがない。針葉樹以外の壁孔膜の水に満たされた孔は、極めて小さいので、それが気泡の移動の障壁となる。このような両タイプの壁孔膜が、気泡の拡散を防ぐ。木部内の気泡による水流の遮断を「塞栓」と呼ぶ。
細い管の中に液体が流れるためには、途切れない液体の流れが不可欠である。万が一。管に気泡が入り空気塞栓ができても、壁孔を通して隣の管に流し変えて塞栓部分を迂回できる。
植物は、蒸散により水を失い続けるため、脱水を防ぐ仕組みを進化させた。 植物は、木部キャビテーションの影響を、いくつかの方法で最小限に抑えている。木部中の水輸送通路は、互いに連絡されているので、原理的には1つの気泡の広がりで、全ての仮道管が満たされる可能性がある。だが、膨張する気泡は、壁孔膜の微細な孔を通過できないので広まらない。
仮道管は、長く伸びた中空の紡錘形で、高度に木化した細胞壁を持つ死んだ細胞である。垂直に重なり合う配置である。細胞壁は、細胞どうしを接着している細胞間層に、薄い一次壁と厚い二次壁が堆積してできており、壁孔はこの二次壁が欠如した部分、一次細胞壁のみにある非常に小さな領域である。植物細胞の細胞壁の一部で、隣り合う細胞同士の水分などの通路となる壁の薄い部分をいう。水は、この細胞壁の側面にある多くの壁孔を通って、仮道管同士の間を流れる。1つの仮道管の壁孔は、多くの場合隣り合う仮道管の壁孔と向かい合うように配置されている。これを壁孔対(pit pair)と呼ぶ。壁孔対は、仮道管間の移動抵抗の少ない水の通路を構成する。隣り合う壁孔対に挟まれた、水を通す多孔性の層は、両仮道管の2枚の薄い一次細胞壁とそれに挟まれた中葉からなり、壁孔膜(pit membrane)と呼ばれている。 また、木部の毛細管は互いに繋がっているので、1つの気泡で完全に水の流れが止まることはない。しかも、水は遮断された道管と隣接する水で満たされている道管を使って迂回できる。木部の仮道管と道管の長さに限界があるため、水の流れに抵抗を与えるが、逆に、それが、キャビテーションの影響を回避させることに繋がっている。
木部から、気泡を除去することもできる。植物の中には、木部に陽圧(根圧)が生ずるものもある。陽圧は気泡を収縮させて、その気体を水に溶け込ませる。また多くの植物は、二次成長を行い、毎年新しい木部を形成するが、道管を新しくすることによって、キャビテーションによる水輸送機能の欠点を回避している。
最近の研究では、木部の水が張力を受けている際も、キャビテーションが修復されていることが観察されている。 葉から大気へ流出する過程で、水は木部から葉肉(ようにく)細胞の細胞壁の中に引き込まれる。細胞壁から蒸発して葉の空気間隙に入る。その水蒸気は、気孔を通って葉から出ていく。
葉の生きた組織を通した水の移動は、水ポテンシャル勾配によって調節される。しかし、水蒸気の移動は拡散によるため、最後の蒸散流の段階では、水蒸気の濃度勾配に支配される。
植物を含むすべての生物は、成長や生殖はもとより高度に組織化された構造を保ちながら修復するために、絶えず自由エネルギー必要をとする。生化学的反応・溶質の蓄積・長距離輸送のような過程は、すべて自由エネルギーの入力により駆動されている。
水の化学ポテンシャル(chemical potential)は、水の 自由エネルギーの定量的表現である。熱力学では、自由エネルギーは、仕事をするための能力potentialを表す。化学ポテンシャルは、物質が持つ自由エネルギーのうち、仕事を行うに利用可能なものを言うが、それは相対値に過ぎない。化学ポテンシャルとは、ある状態における、ある物質の化学ポテンシャルと、同じ物質の標準状態における化学ポテンシャルの差で表せられる。その単位とは、物質1molあたりのエネルギーであるので、単位はJ/mol-1で示される。
葉からの蒸散は、2つの主な要因に頼る。葉内の空気間隙と外気との水蒸気濃度の差と、その径路の拡散抵抗に関係する。大気の水蒸気濃度の測定は容易だが、葉のそれを決定するのは難しい。葉の空気間隙の体積は小さいが、水が蒸発する湿った細胞表面の面積は比較的大きい。空気間隙の体積と、葉の体積との比は、マツの葉では5%、トウモロコシの葉では10%、オオムギでは30%、タバコでは40%である。
空気間隙の体積とは対照的に、水が蒸発する葉内の表面積は、葉の面積の7~30倍である。体積に対してその表面積の割合が高いため、葉内の速い蒸気平衡が可能となる。こうして葉内の空気間隙と、水が蒸発している細胞壁表面の水ポテンシャルが平衡に近いと見られる。
蒸散の起きている葉内の水ポテンシャルの範囲内では、平衡水蒸気濃度は飽和水蒸気濃度より2%以内の差である。これにより測定が容易な葉温から、葉内の水蒸気濃度を測定することを可能にした。空気の飽和水蒸気量は、温度とともに指数関数的に増加するので、葉温は蒸気速度に大きな影響を持つ。
| 葉から水分消失にいたる径路の4カ所における相対湿度 | |
| 場所 | 相対湿度 |
| 葉内の空気間隙 (25℃) | 0.99 |
| 気孔のすぐ内側 (25℃) | 0.97 |
| 気孔のすぐ外側 (25℃) | 0.47 |
| 外気 (20℃) | 0.50 |
裸子植物gymnospermは、ギリシャ語でnaked seedを指す語が語源であり、naked(néɪkɪd)とは、「裸の・裸体の・毛ない」の意味である。種子植物が進化する過程での初期の形態で、針葉樹・ソテツ類・イチョウと薬用植物のマオウなどのグネツム類があり、現在では1,000種類が確認されている。その最大のグループが針葉樹coniferで、マツ・モミ・トウヒ・セコイアなどがある。
Coniferとは、球果植物、球果植物門Coniferophytaとは、その種子がかさ状の構造に包まれるものを指す。球果の中に剥き出しの種子を持つ点で、被子植物と区別される。マツ門とも言う。現生種はすべて木本である。
約1億4,500万年前に現れた被子植物angiospermは、単子葉植物・真正双子葉植物とモクレン類を含む基部被子植物の3グループからなる。種子を包む心皮(しんぴ)を持つ点で、裸子植物と区別される。カナダ・アラスカ・ユーラシア大陸北部の巨大な針葉樹林を例外として、最もよく見る植物が被子植物である。現在約37万種の被子植物が同定されている。 分類学者によればコンピュータモデリングで更に1万7,000の未記載種の存在が予想されている。
angiospermは、ギリシャ語でvessel(容器・うつわ)、 seedを指す語が語源となっており、種子植物が獲得した最も顕著な形態的特徴が花であり、そのため種子植物は顕花植物とも呼ばれる。種子植物を2つに分けたうちの一群。種子が露出する裸子植物に対して、雌性の胞子葉に相当する心皮が1つまたはいくつか集ってできた子房の内部に、種子が保護されている植物をまとめた群である。
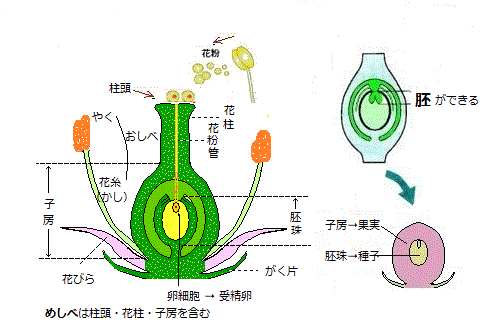
「胚発生」とは、単細胞(受精卵)から、ある特徴を形成しながら、基本的な形態を維持した一つの多細胞体に変化する過程を表す。殆どの種子植物の胚発生では、花の雌ずいの中の、胚珠と呼ばれる特殊な構造の内部で起こる。胚の発生過程の順序が予想できるのは、胚が母親の珠皮の中にきちんと包み込まれることが、種子の形成には重要だからであり、そこには植物の基本的なパターン形成過程の中で、特に重要な過程がいくつか含まれているからである。
そこには極性を確立する過程もある。胚の極性は、胚の中の位置に応じて、どの細胞に分化するのかを決める枠組みを与える。この枠組みの中で、複数の細胞群が表皮や皮層、維管束組織などに機能的に特殊化する。
頂端分裂組織を作る細胞群は、シュートと根のそれぞれの成長点に形成し、胚発生以後の栄養成長では、組織や器官を次々と生み出していく。胚発生が終わると、多くの生理的変化が起こり、胚は長い休眠状態になり、過酷な環境状態を凌いでいくこになる。
目次へ