| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透
| 目次 |
| 1)拡散 |
| 2)浸透 |
| 3)水ポテンシャルwater potential |
| 4)溶質ポテンシャル |
| 5)圧ポテンシャル |
| 6)重力 |
| 7)植物細胞の水ポテンシャル |
拡散diffusionは、物質の濃度が場所によって異なるとき、これを放置すると物質の移動が起こり、全領域にわたって濃度が均一になる現象である。これは、分子の熱運動によって起こる。
異種の物質の混合系が不均一な濃度分布をもつとき、一様な分布に近づこうとする現象を拡散または分子拡散という。また、浸透は隔膜を通じて起る拡散である。
拡散は気体・液体だけでなく、固体の中でも起る。拡散現象は気体・液体または固体の媒質の中で、原子・イオン・分子の群が移動することで、一つの物質による他の物質における透過作用で、気体を通じた気体の移動、液体を通じた液体の移動、固体を通じた固体の移動などが起こる。
岩石を構成する鉱物結晶の内部では、各原子は温度などの外的条件によって振動しており、常に微小な距離を移動する確率をもっている。外部から化学成分の濃度勾配や応力歪などの力が加わり、さらに温度の上昇や格子欠陥などの要因が加わると原子は移動しやすくなる。一様な温度の静止混合流体中の濃度が均一でないときには、濃度を一様な分布に近づける変化が起こる。また温度の分布が不均一ならば熱拡散thermal diffusionが起こる。
媒体が固体の場合、個体内部の成分が固体媒体の内部を移動する拡散現象が主要な役割を果たすことになる。
岩石のような多結晶の集合体の中で拡散が行われる場合、拡散の行われる結晶の場所によって、成分の移動結果はかなり異なる。結晶を作っている内部で、原子・原子団・分子・イオンなどによる、立体的で周期的な規則正しい格子状の配列を結晶格子と言う。一般に結晶内部では結晶格子を置換しながら、原子が格子内を流れる格子拡散lattice diffusionが行われる。格子拡散でも結晶中に欠陥が存在すると速くなる。 これらは物質を構成する微粒子の熱運動によるもので、分子運動論によって輸送現象として解明されている。一般に物質が移動する場合の駆動力は、各場所における成分の化学ポテンシャルの勾配により生じ、その結果、物質が移動する現象が拡散である。
また、風によって煙が広がるのは、流体の流れが乱れて生じる渦の攪拌により、流体系中の物質が混合されるためで、渦拡散eddy diffusionと呼ぶ。流速が大きくなるほど著しい。
細胞機能の諸過程は、細胞内外への分子の輸送に依存している。拡散は、濃度の高い所から低い所への物資の自発的移動である。細胞レベルの大きさでみると、拡散は物質の主要な輸送様式である。選択的透過性をもつ境界面を介した水の拡散は、浸透osmosisと呼ばれる。
拡散は、分子のランダムな熱運動により生じる。溶液中の分子は静的ではない。拡散とは、分子が緩やかに混合され、その結果、濃度差が解消される。分子は連続的に動いており、互いにぶつかりあって運動エネルギーを交換している。どのような分子でも、衝突後にはランダムな軌道をとるとみられ、これらのランダムな動きが分子に正味の移動をもたらす。
最初に、異なる分子を含む2つの材料を接触させる。気体・液体・個体で起こる拡散は、気体中が最も速く、液体中では遅れるが、個体中が最も遅い。
溶液を等しい体積の2つの区画(AとB)に分ける平面を仮定する。すべての分子はランダムに動いているので、いずれの溶質分子も一定時間にある確率で仮想平面を横切る。ある一定の時間にAからBへ移動する分子数は、その時間内の最初のAの分子数に比例する。またBからAへ移動する分子数は、Bの分子数に比例する。
もしAの濃度がBより高いなら、BからAへの溶質分子の移動よりも、AからBへ移動の方が多い。この場合、AからBへ溶質の正味の移動が起きていることになる。従って、各々の分子がランダムに動いても、拡散は濃度の高い方から低い方へ正味の分子移動が起こる。各々の分子は独立して動くから、それぞれの区画内の分子数は等しくなり、正味の移動が少なくなる。この平衡状態では、2種類の分子がランダムに均一に分布する。そのためすべての地点と時刻で溶質中の2分子それぞれの合計は一定している。
均等に分子が分布する傾向は、熱力学第2法則second law of thermodynamicsで説明される。熱の流れは高温部から低温部に向かって不可逆に起きることを表す法則である。この法則はさまざまな形で表現される。たとえば「外部になんら変化を残さずに、熱が低温部から高温部へ移動することはない」(クラウジウスの原理)や「外部になんら変化を残さずに、熱が全部仕事に変わることはない」(トムソンの原理)などである。両者は一見異なっているように思われるが等価であることが証明できる。この法則は、自発的な過程は自由エネルギーの減少、あるいは無秩序の方向へ進行することを表す。拡散は最も低いエネルギー状態に向かい、物質が移動する自然な流れを言う。
拡散係数diffusion coefficientは、熱力学的な指標で、ある特定の媒体内で拡散する分子の平均速度で、その物質の分子が移動する容易さを示す比例定数である。拡散係数は、物質の特性であり、媒体と温度の両方に依存している。その係数が大きいほど、物質が速く拡散することを示す。大きな分子の拡散係数は小さく、空気中の拡散は、液体中の拡散より一般的に1万倍速く、温度が高いと更に速く拡散する。
フィックの拡散の第一法則Fick's first law of diffusionは、流体に溶けているある物質に注目する。流体にある物質が、ある部分は濃く、ある部分は薄い濃度差(濃度勾配)がある状態だと、流体の中で、その物質に流れが生じる。逆に言うと、流体のなかの濃度が均一であった場合、その物質に流れは生じない。物質の流れは、いつも濃度が濃いほうから薄いほうへ向く。その濃度勾配による流れを定量化していくために、まずフラックスfluxという概念を導入した。フラックスとは、「単位時間当たりに、単位面積を通過する物質の量」と定義される。フラックスの駆動力は濃度勾配であり、濃度勾配が大きいほどフラックスの絶対値は大きくなる。
拡散は、短い距離では効果的である。一定時間後には、分子のランダムな動きにつれて溶質分子集団の先端は、スタート地点から遠ざかる。しかし、個々の分子は行ったり来たりするようなランダムウォークをするので、ある物質が一定距離を拡散するのに必要な平均時間は、距離の2乗に比例する。
直径50㎛の細胞の水中を、グルコース分子が横切るのに要する時間は、平均2.5秒である。それでは、同じグルコース分子が、水中を1m拡散するだけで約32年かかることになる。それでも、溶液中の拡散は、生物の微細な細胞では、大いに有効なのである。
目次へ
浸透osmosisは、選択的透過性をもつ障壁を横切る水の正味の移動を表す。植物細胞の生体膜は、この原理により物質を選択的透過selectively permeableする。生体膜は、水や電荷のない小さい物質を、電荷のある物質や大きな溶質よりも、速やかに通過させる。細胞内の溶質濃度が、細胞外の溶液よりも高い場合、水は細胞内に拡散するが、細胞内の溶質は細胞外へ拡散しない。この選択的透過を持つ障壁を横切る水の正味の移動を浸透と呼ぶ。
拡散では、溶質は溶液全体に均一に分布するが、浸透では、溶質が移動できる範囲は、膜により制限されている。膜が通した溶媒の拡散により溶質を希釈する。実際に、水の流入を防げなければ、すべての水は膜内の溶質側に流れる。
純水が入ったビーカーに細胞を入れると、細胞には選択的透過性を持つ膜があるので、細胞壁を持たない動物細胞では、選択的透過性を持つ膜により浸透して、やがて破裂し拡散する。植物細胞では、強靭な細胞壁があり、水が細胞に流入すると細胞壁に圧力が生じ、この圧力によって水の流入が抑制され、細胞の体積の増大は物理的に制限される。
植物の細胞壁は、その膨圧により大きな抵抗となって、細胞内の静水圧を上昇させる。浸透osmosisは、ギリシャ語の「ωθώντας(othóntas)」を語源にし、pushingと同義である。osmosisとは、植物細胞内の溶質の増大による細胞壁の陽圧を表している。
目次へ
植物を含むすべての生物は、成長や生殖はもとより、高度に組織化された構造を維持するための修復などに、絶えず自由エネルギーを必要とする。組織内の生化学反応や溶質の蓄積、さらに長距離輸送のような過程は、すべて自由エネルギーにより駆動される。
植物による水の吸収は、根の根毛などの若い細胞が主として水を吸収し、これが根の導管にまで移動し、根と茎の導管を通り、葉の葉脈を経て葉の個々の細胞に移動し、結局、その~97%が気孔から蒸散され空気中に放出される。水が植物体内を移動できるのは、上記のそれぞれの段階を踏まえて、土壌~根毛の細胞、根の細胞~根の道管・髄、茎の道管、葉柄、葉の道管~葉の細胞、気孔~大気の間に、水ポテンシャルが生じることにより、それぞれの段階で、水が移動する原動力となっているからだ。
水の化学ポテンシャルは、水の自由エネルギー状態を言う。その化学ポテンシャルchemical potentialは、物質の自由エネルギーの定量的表現である。熱力学では、自由エネルギーは仕事(力×距離)をするための能力potentialを表す。ちなみに、物体の運動は、物体の速度が大きいほど、また、物体の質量が大きいほど、激しいといえる。よって、物体の運動の激しさを表すときの量を運動量といい、物体の運動量は、速度×質量で表す。運動量を変化させるには物体に力を加えればいい。このとき、加える力が大きいほど運動量は大きく変化する。その際、力を加えている時間が長ければ長いほど運動量は大きく変化する。よって、運動量を変化させる効果のある量を考えるとき、力×時間で表し、この量を力積という。
化学ポテンシャルの単位は、モル(物質量の単位;12gの炭素12の中には、炭素原子が6.02×1023個存在する。この6.02×1023個の粒子の集団を1モルという。)あたりのエネルギー(Jnol-1)で表示される。ただ、化学ポテンシャルは、相対値に過ぎない。化学ポテンシャルとは、ある状態における物質の化学ポテンシャルと、同じ物質の標準状態における化学ポテンシャルの差を表す。水の化学ポテンシャルも、水に関する自由エネルギーの差で表される。水は、化学ポテンシャルが高い所から、低い所へ、エネルギーの入力を加えることなく自発的に流れる。植物の水ポテンシャルに影響を与える主な要素は、「濃度」・「圧力」そして「重力」である。溶液の水ポテンシャルの構成要素は、「溶質」・「圧力」そして「重力」で、水の自由エネルギーに対する寄与を示す。
水ポテンシャルを定義するために最も頻繁に用いられる基準状態は、大気の温度と標準大気圧下での純水である。基準となる高さは、植物全体を用いた研究では、その植物の基部に、細胞レベルの研究では、調べている組織に設定されることが多い。
目次へ
溶質ポテンシャルsolute potentialあるいは浸透ポテンシャルosmotic potentialと呼ばれるのは、水ポテンシャルに対する溶質の効果を表す。溶質ポテンシャルでは、溶質自体の固有の性質には、通常、左右されない。むしろ、溶質は水で薄まることなり自由エネルギーを減少させる。則ち、溶質と水の混合は、エントロピーを増加させるため、自由エネルギーを減少させることになる。言い換えれば、基準である純水と比べて溶液中の溶質が水ポテンシャルを減少させている。加えて、温度も水ポテンシャルに影響する。
熱力学第二法則は、「何かの現象が起こるとき、エントロピーentropyは必ず増大する。また、勝手に減少する事は無い」という。「エントロピー」とは、熱力学では、物体や熱の混合度合いのことで、つまり熱力学第二法則は、「熱は高温から低温に移動し、その逆は起こらないという法則」を言う。
「エントロピー」における法則は、「溶質は溶けている溶媒全体に拡散する」現象にも表れる。しかし、細胞には選択的透過性を持つ膜があるため、細胞内の溶質濃度が、細胞外の溶液よりも高ければ、外部の水は細胞内へ拡散するが、細胞内の溶質は細胞外へ拡散しない。この選択的透過性を持つ膜を通過する水の正味の移動を「浸透」と呼ぶ。
化学ポテンシャルにより、選択的透過性を持つ膜が、自由に拡散する溶質を、破裂するまで受け入れ程、細胞は膨張し続けるだろうか。細胞壁を持たない動物細胞では起こりえるが、水を絶え間なく吸い上げる根がない動物細胞では、通常、生じない現象である。
極めて強い細胞壁を持つ植物細胞では、水が細胞内に流入し細胞壁に圧力がかかれば、細胞壁は大いに変形する。それに抵抗をして、水の流入を細胞壁の強靭な圧力によって、細胞の体積の増大を制限する。これが細胞内の「静水圧」の上昇に繋がる「根圧」である。
実質的には、多細胞系からなる根の組織は、まるで浸透膜であるかのように、溶質の蓄積によって木部中に陽圧を生じさせる。植物体内の水の上方への移動は、水の凝集力を介して行われる蒸散により引き上げられ、しかも根圧の押し上げも加わる。根圧による水の移動は、蒸散が行われていない場合でも見られる。
気体と違い、液体や固体が凝集して一定の体積をもつのは、分子・原子またはイオン間に引力が働くからで、この力を凝集力という。しかし、凝集力だけに頼れば、固体や液体はその体積を縮めるだけである。そのため、短い距離間では反発力が作用している。一般に,凝集力は反発力に比べると遠くまで作用する。凝集力には、分子間の静電引力やファンデルワールス力(2個の原子が結合した場合に必ず起こる、電荷の揺らぎによって生じる弱い結合性相互作用による電気的引力)、水分子同士の水素結合やイオン間で働く電荷移動力などが働く。
根圧は、土壌の水ポテンシャルが高く、蒸散速度が小さいときに、最も生じやすい。蒸散速度が増すにつれ、水は植物を通して輸送されて素早く大気へ失われるので、イオンの取り込みにより、溶質濃度の高まりに伴い発生する陽圧は、木部中では生じない。
根圧を生じるような植物は、葉の縁に水滴を付けることが多い。それが排水という現象で、木部の陽圧は、葉の縁の葉脈の末端に繋がる排水組織と呼ばれる特殊化した孔を通して木部液を排出する。
朝、草の葉の先端に見られる「露滴」は、実際は「水孔」と呼ばれる排水組織から排出された液である。この現象を「溢泌」と言う。「溢泌」は蒸散の少ない状況下などで、植物内の過剰水分の排出に働いていると考えられている。そのため「溢泌」は、夜のように蒸散が低下し、相対湿度が高いとき最もよく見られる。
溢泌液は、植物の道管液から排出されるため、様々なイオン類が含まれている。「水孔」周辺の細胞には、その植物にとって貴重な栄養素を、溢泌液の中からもう一度、植物体内に取り戻す仕組みが存在している。
根圧は、高いイオン蓄積速度によって生じる不可避な結果を反映している。夜の木部内での陽圧は、気泡の溶解を助け、それによりキャビテーション(cavitation)の有害な作用が打ち消される。Cavitationとは、空洞化、空洞形成の意味であり、「道管内の小さな気泡が大きく膨張することにより起こる水柱の張力の崩壊」を言う。
目次へ
圧ポテンシャルpressure potentialは、水の自由エネルギーに対する静水圧の効果を表す。陽圧は水ポテンシャルを増加させ、陰圧はそれを減少させる。陽圧と陰圧のいずれも植物内で生じる。細胞内の正の静水圧は、「膨圧」とも呼ばれる。木部道管内で、幾度も発生する負の静水圧が、張力tensionである。張力は植物内の水の長距離輸送には欠かせないほど重要である。
液体であるため、個々では弱い水分子間の水素結合であるが、水分子の立体構造から、隣接する4つの水素結合の連結により、非常に強い分子間相互作用が働き、水分子間で多元的な張力生み、巨大な被子植物相を育てた。
水は20MPa(メガパスカル)以上の張力にも耐えられることが研究で分かった。
車のタイヤは約0.2MPaまで空気を入れるという。家庭の水道管の水圧は、多くの場合、0.2~0.3MPaと言う。水深10mの水圧は、0.1MPaである。
目次へ
重力は、大きさが等しい反対方向の力と拮抗していなければ、水を下方向へ流す。それを重力ポテンシャルgravitational potentialと呼び、それは、基準状態の水からの高さと水の密度、そして重力加速度によって計算される。
しかし、隣り合う植物細胞間の重力ポテンシャルの差は、浸透ポテンシャルや、細胞内の正の静水圧「膨圧」と木部道管内の負の静水圧「張力」による圧ポテンシャルと比べると無視されるほど小さく、細胞レベルの水輸送における水ポテンシャルでは、溶質ポテンシャルと圧ポテンシャルの和で計算されるほど簡略化される。
目次へ
植物細胞の成長や光合成、さらには穀物生産も含めて、すべてが水ポテンシャルとその構成要素に強く依存している。植物細胞内の水の自由エネルギーは、通常、周囲の温度や大気圧が同一であれば、その高さが同じである純水の自由エネルギーより小さい。そのため、細胞周囲の溶液の水ポテンシャルが変化すると、水は浸透により細胞内へ出入りする。
水は大気と接しているので、水の圧ポテンシャルは大気圧と同じである。細胞膜を介した水の流れは、受動的な過程であり、水は物理的な力に応じて低い水ポテンシャルや自由エネルギーに向かって移動するのである。
水ポテンシャルの勾配に逆らって、半透明膜を介した水輸送が起こる唯一の例は、溶質移動と共役する場合である。様々な膜タンパク質による糖やアミノ酸、および他の小分子の輸送は、輸送される溶質分子あたり260個に及ぶ水分子を、膜を介して引っ張り込む。
このような水輸送は、水ポテンシャル勾配に逆らった方向、即ちより高い水ポテンシャルの方向にも起こりえる。それは、溶質の自由エネルギー減少が、水の自由エネルギー増加を上回るからである。この時の正味の自由エネルギー変化は負のままである。通常、この方法で輸送される水の量は、水ポテンシャル勾配の低い方向へ流れる受動的な水の移動と比べてかなり小さい。
水ポテンシャルとその構成要素は、成長状態と植物内の位置で変わる。よく潅水された植物葉の水ポテンシャル(ψ;ギリシャ文字のプサイ)は、草本植物で-0.2~-1.0MPa、また木本及び灌木植物で-0.2~-2.5MPaの範囲にある。乾燥地域に生育する植物葉では、水ポテンシャルはずっと低く、極限状態では-10MPa以下にまで下がる。
水ポテンシャルの値が成長条件や植物の種類に依存しているように、溶質ポテンシャルの値もまた大きく変化する。典型的な植物細胞の溶質ポテンシャルは-1.2~-0.8MPaの値であるが、よく潅水された菜園の植物細胞であるレタス・キュウリの芽生え・豆の葉などでは、-0.5MPaくらいまで高くなる。つまり、細胞内の溶質濃度は水の含有量が高まるため低くなる。木本植物では、溶質ポテンシャルは低い傾向にある。つまり、細胞内の溶質濃度が高い傾向にある。多くの場合、真昼には膨圧の低下なしに、水ポテンシャルはさらに低下する。
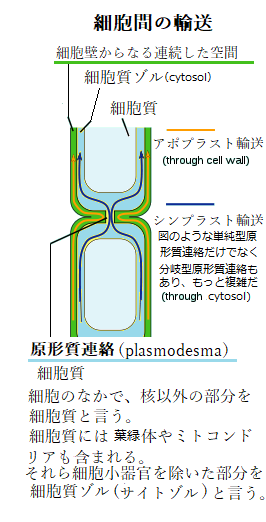 細胞内の溶質ポテンシャルは、大きな負の値を持つにも関わらず、細胞外のアポプラストapoplast溶液、即ち細胞壁や木部は殆ど溶質を含んでいない。
細胞内の溶質ポテンシャルは、大きな負の値を持つにも関わらず、細胞外のアポプラストapoplast溶液、即ち細胞壁や木部は殆ど溶質を含んでいない。アポプラストは、植物組織において細胞膜よりも外側の細胞壁や細胞間の空間の総体、及び死んだ細胞の内腔、例えば木部道管や道管繊維などが連続した系で、その径路で、水が根の皮層を横切る時、膜を横断する必要がなく、連続して移動できる領域を言う。言わば、水溶液(アポプラスト液)で満たされた空間の総体で、アポプラスト輸送apoplast transportとは、アポプラストと呼ばれる「細胞壁からなる連続した空間」を通る分子の移動のことである。隣接した細胞群の互いに繋がった細胞壁を通す物質の移動になり、分子は、細胞膜を横切ることなく植物体内を移動できる。通常、植物組織の全体積の5~20%が細胞壁である。植物体内におけるアポプラストは、水や溶質の輸送路であり生化学反応の場でもある。特にその細胞壁は単なる物質の移動経路としてのみならず、物質代謝の場としても機能している。
アポプラストは、植食性昆虫や動物による食害に、全身的に抵抗する。その物理的損傷に応答して、植物組織の障壁を分解する加水分解酵素が植物側から放出される。このような酵素による生成物の殆どは、アポプラストに存在しており、damage associated molecular pattern(DAMP;損傷関連分子パターン)と呼ばれる内生のエリシターelicitor(自然免疫を活性化させる自己生成する誘導因子)として機能して、広範囲の生物に対する抵抗性を誘導する。
成長中の果実や生育地が高塩環境であれば、アポプラストの溶質濃度が、高いにもかかわらず、アポプラストの溶質ポテンシャルは、-0.1~-0MPaの範囲にあることが多い。
シンプラストsymplastとは、アポプラストと対をなす輸送路である。植物細胞の細胞膜内で、隣接した細胞同士を繋ぐ、直径40~50nmの管により高頻度で連結されている連続体を言う。細胞質ゾル(サイトゾル)同士は、分岐した原形質連絡や、枝分かれのない原形質連絡によって、植物細胞間では、複数にわたる繋がりが、それぞれ連続体を形成している。
シンプラストにより、細胞間の低分子輸送をはじめ、溶質の移動が、拡散によって容易に行われると考えられている。それをシンプラスト輸送symplastic transportと呼ぶ。その原形質連絡は、タンパク質や核酸など、様々な巨大分子の輸送経路となり、隣接する細胞間の情報交換を仲介している。 アポプラスト輸送とシンプラスト輸送は、いずれも維管束系で重要な役割を担っている。
よく潅水された植物細胞内の圧ポテンシャルは、溶質ポテンシャルに依存して0.1~最大3MPaの値に達する。そのような組織の細胞内膨圧が0に向かって低下すると、植物はしおれる。多量の水が細胞から失われると、細胞壁は機械的に変形し、結果的に細胞は損傷される。
目次へ