| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
地球大気は、陸上植物が生活するには厳しい環境である。大気は光合成に必要な二酸化炭素の供給源であるが、その一方、かなり乾燥しており、蒸散により水の消失や脱水の脅威を招く。
水は植物が生きていく上で、極めて重要な働きをしている。植物は水の消失なくして、二酸化炭素を体内に取り込むことはできない。その二酸化炭素の取り込みと共通した拡散径路を通して、水分を消失するという植物の乾燥の危険を伴う。
二酸化炭素が拡散により葉の中に取り込まれると同時に、葉内の水蒸気は拡散により大気に出ていく。葉内の水分消失を駆動する水の濃度勾配は、二酸化炭素を取り込む濃度勾配よりもずっと大きいので、この問題は一層深刻となる。1分子の二酸化炭素を獲得するために、400分子の水を失っている。この大きな水の消失は、植物の形態と機能の進化に大きく影響した。
水の消失を少なくしながら、できるだけ多くの二酸化炭素を取り込むという矛盾した要求に応えるため、植物は葉からの水分消失を制御しながら、大気に失われた水を、根を使い土壌から吸収し、植物体を通して取り込むことで適応した。
水の吸収やその輸送と、大気への水分消失とのバランスが少しでも崩れると、水不足により多くの細胞活動は、重大な機能不全を引き起こす。
植物細胞と動物細胞との間で、水分生理に大きな影響を及ぼす主な相違点は、植物細胞は細胞壁を持ち、動物細胞は持たないことである。細胞壁は、植物細胞内に膨圧turgor pressure(植物の細胞は、細胞壁を持つために容易に変形できない。そのため、低張状態におくなどして細胞内に水が入ると、細胞壁に内側からかかる圧力が生じる。この圧を膨圧という。)と呼ばれる高い静水圧hydrostatic pressure(静止している液体の中の、すべての方向に均質に働く圧力)を形成させる。細胞が低張液(塩や糖などの溶質の濃度が細胞内液より低い溶液)に接すると、浸透圧によって水が浸入し、細胞容積の増大が起こり、浸透した水によって細胞壁に加わるのが膨圧で、植物細胞には細胞壁があるため、容積の増大は一定の限界で停止する。吸水量よりも蒸散量が多いときはこの圧力が下がり、植物はしおれる。
膨圧は細胞拡大・気孔開口・篩部輸送や、膜を介した様々な物質輸送など、多くの生理的過程に極めて重要である。また、膨圧は木化(ニグニン化)していない植物組織の硬さや力学的安定にも寄与している。
目次へ
水は、植物が成長し機能するために必要な資源の中で、最も豊富であるが、農業生産が阻害される最大要因にもなる。農業生産のために灌漑をするのは、水が農作物にとって重要な資源となるからだ。植物は非常に大量の水を必要とするため、水利用は、生産性においても制限要因となり、降水量によって植生様式に際立った違いが生じる。
根で吸収された水の殆ど(~97%)は、植物体を通して運ばれ、葉の表面から蒸散transpirationされる。根で吸収された水のごく一部が、成長(~2%)、または、光合成や他の代謝過程の生化学反応に消費される(~1%)。
気孔は、若い茎や葉などにあり、特に葉の裏面に多い。特に陸上植物の多くは、その裏側に、水草の葉であれば表側に見られる。
植物の光合成は、太陽の光エネルギーを受けて、二酸化炭素からデンプンなどの光合成産物を合成する。しかし、最も効率がよい条件下でも、光エネルギーの約12%が光合成産物として固定されるだけで、残りは熱エネルギーとなるため、葉の温度を上昇させることになる。そのため、植物の蒸散作用が、葉の温度調節を果たす重要な役割を担う。気孔を通して水を蒸散させ気化するときに、周りから気化熱を奪って温度を調節する。
気孔は光合成が盛んに行われる晴天の時に開いて、葉から水を蒸散させ、根による水の汲み上げと、それに伴う無機養分の取り込みと、同時に光合成に必要な二酸化炭素の吸収、光合成により産出される酸素の放出など、その開閉は、周りの環境条件に応じて厳密に制御されている。
植物は気孔を通して、物質が濃度の高いところから低いところへ移動する拡散の原理によって、水分の放出と二酸化炭素の取り込みを同時に行っている。その仕事をこなすためか、気孔は小さいほど数が多くなる。植物種にもよるが、平均しますと1cm2あたり10,000個以上の気孔を持っており、ファーブルの「植物記」によれば、カーネーションでは、葉の裏側1cm2 あたり約 5,500個の気孔があり、ライラックは、2万3,000個と観察されている。その細い針の先よりも、格段に小さな気孔が、日々重要な役割を果たしている。
「植物記」では、「ヤドリギは、葉の表面1cm2に30個の気孔しか持たない。おそらく、いろんな木に寄生するため、自分の労力に頼る備えもなく、遠慮もなく他の木の樹液を飲むためである」と記す。
一方、雨が降らず晴天の日が続き、植物周辺の水分が不足気味になると、植物に対する成長阻害をストレスとして応答するホルモン・アブシジン(植物内で合成される)が、気孔を閉じ、植物体からの水分損失を防ぐ。
気孔が光に反応して開くことは、1,898年、進化論で有名な C. Darwin の息子である F. Darwin により観察されている。その後、390nm~500nm の波長の青色光が、気孔の開口に特に有効であることが分かった。しかも、気孔が開いた状態の孔辺細胞には、閉じた状態の数倍濃度のカリウムイオンが蓄積しており、カリウムイオンの蓄積により、浸透圧が上昇して水が取り込まれ、孔辺細胞の体積が増加していることが分かった。
孔辺細胞は内側に厚い細胞壁、外側に薄い細胞壁を持っており、孔辺細胞の体積が増加すると外側の薄い細胞壁が押され、孔辺細胞間の孔が開く、つまり気孔が開口する。
水草の葉も、太陽光が当れば、温度が上昇する。しかしながら、根からの水が気孔を通して水中に放出されても、葉の気化熱を奪うことにならない。水中植物の葉は、大気よりも遥かに高い熱伝導度の環境下にあり、いつも「水冷」常態にあり、温度調節を必要としない。そのため気孔が表側にあっても、葉の温度を上昇させず光合成を行うことができる。
蒸散作用のみならず、気孔の機能として、空気・水の交換と、無機養分の移動など重要な働きがあるが、空気交換・水・無機養分の吸収も、葉の細胞と周りの水とで直接進行ができる器官が多いため、陸上植物ほど気孔の蒸散作用を必要としないと考えられている。
植物は、夜間にさえ量は極端に少ないながらも吐出する。朝方、ススキの先の水滴や、キャベツの葉の窪みに丸くなった水滴は、夜間の発汗が冷気により凝結したものだ。
陸上植物は、土中の無機養分が微量であるため、生殖地で汲み上げ作業を途中でやめられない。昼夜を問わず、根は湿気のある大地から汲み上げた大量の水に溶け込んだ中から、僅かな無機養分を選別して吸収しなければならない。
土中の栄養分は、細胞に貯えられ、植物体の必要度に応じて活用されるが、そのために大量に汲み上げられた水の多くが厄介者になる。葉の瑞々しさを保ち、水道を洗ったりするためにも、水は欠かせない。水は、絶えず根から汲み上げられる。そのため余分な水は、気孔から日陰や夜間を問わず蒸散を続ける。
それで、美しい朝露の朝を迎えることになる。
目次へ
生命は、軽い元素から出来ている。地球生命の誕生から40億年、生命の基本は、水素(原子番号1)・炭素(原子番号6)・窒素(原子番号7)・酸素(原子番号8)・ナトリウム(原子番号11)・マグネシウム(原子番号12)・リン(原子番号15)・硫黄(原子番号16)・塩素(原子番号17)・カリウム(原子番号19)・カルシウム(原子番号20)などの原子の質量が小さい元素から成り立っているが、それは地球がまだ固まっていなかった頃、重い元素が地球中心部に沈んでしまい、地表近くには軽い元素しか残らなかった、という事実に由来する。
物質は元素の組み合わせでできている。炭素・酸素・窒素・水素など、元素は化学的手段では分解することも、他の元素に変換することもできない。元素としての化学的性質を保つ最小の粒子が原子である。単一の元素からなる場合を除き、物質の特性は、物質が含む原子の種類と、それらが分子を作る際の結合の仕方によって決まる。そのため物質が作る生物を理解するには、分子の中で原子を結び付けている化学結合のでき方を理解しなければならない。
原子(atom)の中心には密度が高い、正電荷(positive electric charge)を帯びた原子核(atomic nucleus)がある。少し離れてこれを取り巻く電子(electron)の雲は負電荷(negative electric charge)を持ち、原子核とは静電引力(electrostatic attraction;static=静的な, 静止の;反対の電荷を持つ原子を互いに引き付ける力)によって繋ぎ止められている。密度が高い原子核が、原子の質量の大半を占める。これよりはるかに軽い負電荷を帯びた電子が、量子力学の法則に従って原子核の周囲の空間を占める。電子を分布の範囲を示す軌道内に連続した雲として描くのは、ある電子が瞬間ごとにどこにあるのか、正確に予想する方法がないからである。
原子核は、正電荷を持つ陽子と電気的に中性の中性子の2種類の粒子からなり、核内の陽子の数が原子番号となる。水素は原子核に陽子が1個だけであるので原子番号は1であり、元素の中で一番軽い。炭素は6個の陽子があるので、原子番号は6である。陽子が持つ電荷は総べて等しく、電子が持つ電荷とは、+と-で符号が反対になる。1個の原子の中の陽子と電子の数は同じで、原子番号とも等しい。そのため原子の正味の電荷はゼロとなり、全体では電気的に中性になる。この原子番号こそが、元素の化学的振舞いの決め手となる。
中性子は、陽子とほぼ等しい質量を持ち、原子核の構造を安定させる働きをする。核内の中性子の数が変動すれば、核は放射性崩壊を起こすこともあるが、中性子の数によって原子の化学的性質が変わることはない。それで同じ元素であっても、陽子の数が同じであるが、中性子の数が異なる、化学的に違いはないが、物理的に区別できる何種類かの原子が存在するようになる。これらを同位体と呼ぶ。たいていの元素には、複数の天然同位体があり、その中には、不安定で放射能を持つものもある。
水は細胞重量の約70%を占める。細胞内の反応の殆どが水溶液中で起こる。地球上の生物は海で誕生した。生体内で起こる化学反応は水の特性を活用している。2個の水分子間で水素結合することで、典型的な哺乳類の体温でも、液体として存在でき、それが生命体を保持する数々の特性を担っている。
水は様々な物質の溶媒として働き、また植物体内を容易に移動できる特質を持っている。これらの性質は、主に水分子の水素結合と極性構造から生じている。
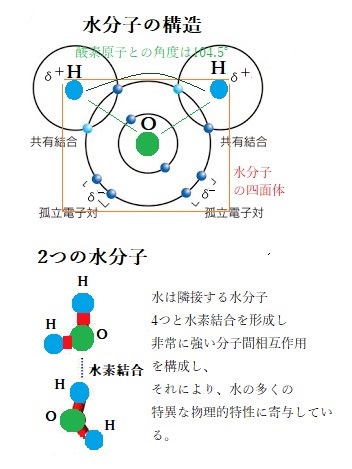 酸素原子は外殻に6つの電子を、水素原子は1つの電子を持っている。水分子(H2O)は、2個のH原子がO原子に共有結合している。水分子の構造は、O(原子番号8)はH(原子番号1)よりもはるかに強く、酸素原子の高い電気陰性度electronegativity(negativity;nɛ̀ɡətɪ́vəti陰性)により、2つの水素原子と共有結合すると、それぞれを合わせた2つの電子を強くひきつけるため、2つの電子は不均等に分布し、各々の水素原子は、部分的な正電荷を帯びる。
酸素原子は外殻に6つの電子を、水素原子は1つの電子を持っている。水分子(H2O)は、2個のH原子がO原子に共有結合している。水分子の構造は、O(原子番号8)はH(原子番号1)よりもはるかに強く、酸素原子の高い電気陰性度electronegativity(negativity;nɛ̀ɡətɪ́vəti陰性)により、2つの水素原子と共有結合すると、それぞれを合わせた2つの電子を強くひきつけるため、2つの電子は不均等に分布し、各々の水素原子は、部分的な正電荷を帯びる。水素の電子を、水素の原子核より酸素の原子核に引き付けるので、2つのH-O結合は極性が非常に大きくなる。そのため水分子は電子の分布に偏りが生じ、2個のH原子は正電荷(δ+)を帯び、反対にO原子は負電荷(δ-)を帯びる。
水分子は、陽子と電子を同数持つので、全体としては中性である。ただ酸素の原子核が水素原子の電子を引っ張るため、水素の原子核は僅かに正電荷を帯び、酸素原子では電子密度が増し負電荷を帯びた領域ができ、水素の2個の電子と酸素の隣り合う2対の電子(4個の電子)が、それぞれ正電荷を帯びる領域と負電荷を帯びる領域と非対称に分布するため極性を持つ。
酸素原子の2つの孤立電子対は、部分的な負電荷を帯びるため、酸素原子側は負に帯電する。異なる2原子から成る化学結合である酸素原子と水素原子間では、酸素原子のほうが水素原子より電気陰性度が大きので、水素原子の2つの電子は酸素原子のほうに引寄せられ、酸素原子と水素原子の結合は極性分子を構成する。その程度は、両原子の電気陰性度の差が大きいほど著しい。ただ、水分子としては、その部分的な帯電が等価であるため、「正味」の電荷を持たない。
三次元的構造である水分子の正電荷領域(H原子)の1つが別の水分子(O原子)の負電荷領域に近づくと、両者の間には電気的な引力が働き、水素結合(hydrogen bond)と呼ばれる弱い結合ができる。
水素結合の強さは、共有結合に比して微弱と言える。その20分の1程度に過ぎず、熱エネルギーによる分子運動によって簡単に切れてしまう。そのため結合の寿命はごく短いが、弱い結合も集まれば見過ごせない効果を発揮する。各水分子が2個のH原子を介して、他の水分子2個と水素結合を作るので、絶えず切れたり繋がったりするが、水素結合は、酸素+水素+酸素と3個の原子が直線に並んだ時が最も強く、水分子における部分的な正・負の電荷(δ+とδ-)は、他の水分子との間に分子間の水素結合のネットワークを作る。
水が室温で液体である時、沸点が高く表面張力が大きいのは、水分子は水素結合により一時的に水分子同士を、三次元的に立体的な格子状に結び付けるからだ。この立体構造により水は凝縮し、表面張力・比熱(物質1gの温度をセ氏1℃上げるのに必要な熱量)・気化熱がいずれも高くなるなど、水素結合なしには、生命体は存在し得ない程、水の重要な特性となる。
水分子は四面体の形状で、その四面体の2頂点に正電荷を持つ水素原子が位置し、残りの2頂点には、負電荷を持つ孤立電子対(原子の外殻電子軌道に属する2つの対となる4個の電子が、他の原子と共有されていない非共有電子対を言う。)が位置する。こうして水分子は、2つの正極と2つの負極を持つ。これらの局所的な正・負の電荷が、水分子間の静電引力を生み出し、水素結合を行う。
「水素結合」は、酸素のような電気陰性度の高い原子が、水素原子と共有結合するような効果的な静電的な結合が形成される場合に使われる。サイズが小さい水素原子が、より密度が高い正電荷を作り出すためには、静電的な結合がより効果的である。
水素結合では、水の多くの特異な物理的特性を形成する。特に水分子の立体構造から、隣接する4つの水素結合により、非常に強い分子間相互作用が機能的に働く。その結合は、水と電気陰性度の高い酸素原子や窒素原子などの間でも形成される。しかも、これらの原子が、水素原子と共有結合している場合に形成されている。
目次へ
水は他の類似の溶媒よりも多くの種類の物質を大量に溶かす。この溶媒としての優れた特性は、一つには水分子のサイズが小さいことによる。また高い水素結合能と極性により水は極めて優れた溶媒になり、とりわけ、イオン性物質と極性基である水酸基やアミノ酸基を含む糖やタンパク質をよく溶かす。
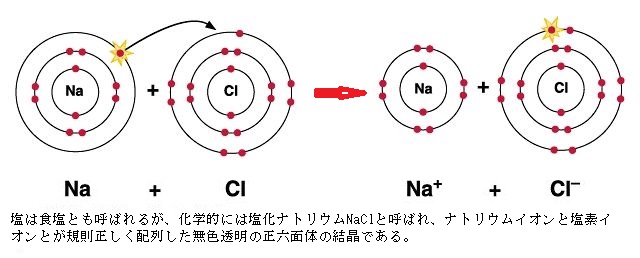 イオン結合(ionic bond)は、電子を共有結合するよりも、他の原子に電子を移すか、電子を受け取る方が、最外殻を満たしやすい原子間で形成される。ナトリウム(Na;原子番号11)原子は、最外殻にあたる第3電子殻に電子が1個あるだけであるから、その1個を放出できれば第2電子殻が最外殻になるので、最外殻が満たされる。塩素(Cl;chlorine)は、最外殻にあたる第3電子殻に電子が7個ある。電子を1個得られれば最外殻は満たされる。
イオン結合(ionic bond)は、電子を共有結合するよりも、他の原子に電子を移すか、電子を受け取る方が、最外殻を満たしやすい原子間で形成される。ナトリウム(Na;原子番号11)原子は、最外殻にあたる第3電子殻に電子が1個あるだけであるから、その1個を放出できれば第2電子殻が最外殻になるので、最外殻が満たされる。塩素(Cl;chlorine)は、最外殻にあたる第3電子殻に電子が7個ある。電子を1個得られれば最外殻は満たされる。Na原子がCl原子と出合うと、電子が1個、Na原子からCl原子に飛び移り、双方の最外殻が満たされる。
ナトリウムは、非常に反応性の高い軟らかい金属で、日本の工業分野では、特に化合物中のナトリウム成分を曹達(ソーダ)と呼び、「毒物及び劇物取締法」により劇物に指定されている。それが緑色の有毒気体である塩素とイオン結合して、美しい無色透明な結晶となり塩化ナトリウム(NaCl)という化合物・塩ができる。それが、地球上の大半の生物にとって、生命維持に欠かせない重要な物質となる。
Na原子からCl原子に電子が飛び移ると、互いの原子は電荷を帯びた2個のイオン(ion; άɪən)が生じる。マイナスの電子を1個失ったNa原子は、正電荷を持つ陽子より負電荷の電子が一個減るため、原子は1価の正電荷を持つ(Na+)。電子を1個余分に獲得したCl原子は、陽子より電子が1個多くなり、1価の負電荷を持つ(Cl-)。Na+とCl-は、反対の電荷を持つため静電引力で互いに引き合うようになる。
イオン結合したイオンは、塩化ナトリウムの結晶ができる。結晶には、ナトリウムイオンと塩化物イオンが三次元的に正確にぎっしり詰まり、反対の電荷はきっちりとつり合っている。1mm角の結晶には、天文学的な数のNa+とCl-がそれぞれ約2×1019(千京)個正確に並んでいる。イオン結合したイオンは、分子ではなく塩(えん)と呼ばれる。
水分子とイオン間または水分子と極性物質間の水素結合は、溶液中で帯電した物質同士の静電的相互作用を効果的に減らし、それら物質の溶解度を増加させる。同様に、水分子とタンパク質や核酸などの巨大分子との間の水素結合は、巨大分子間の相互作用を減らし溶液を溶けやすくする。
水分子間の水素結合により、水は高い比熱容量と高い気化潜熱の両方を持つ。
比熱容量specific heat capacityとは、圧力または体積が一定の条件で、一定量の物質の温度を上げるために必要なエネルギーである。温度は分子の運動エネルギーの尺度になる。水温が上がると水分子はより速く、より大きく振動する。水素結合は、加えられた熱からエネルギーの一部を吸収するゴムバンドのように作用し、熱運動に使われるエネルギーを減らす。つまり、他の液体に比べ、水は温度を上げるために比較的多くの熱を必要とする。このような水の性質は、植物にとって温度変化を緩和するのに重要である。
| 物質 | 温度 | 比熱(J/g・K) |
| 水 | 0 | 4.217 |
| 氷 | -1 | 2.100 |
| アルミニウム | 0 | 0.880 |
| 鉄 | 0 | 0.435 |
| 銅 | 0 | 0.379 |
| 黄銅 | 0 | 0.387 |
| 銀 | 0 | 0.235 |
| 水銀 | 0 | 0.140 |
| 鉛 | 0 | 0.129 |
| 木材 | 20 | 1.250 |
| ポリエチレン | 20 | 2.230 |
| ガラス | 10~50 | 0.670 |
気化潜熱latent heat of vaporization(latent;潜在している)とは、液相から分子を分離し、気相に移動させる、蒸散の際に必要なエネルギーである。それは、液体が気体に変わるときに周りから奪う熱のことである。また、冷却水が沸騰蒸発するときには大きな潜熱を奪う。潜熱は物質や温度によって変化する。逆に、水が氷になるときには熱を放出する。これを凝固潜熱という。
潜熱latent heatとは、物質の相が変化するときに必要とされる熱エネルギーの総量である。物質が固体から液体、もしくは液体から気体に相転移するときには吸熱が起こり、逆の相転移のときには発熱が起こる。気化潜熱は、温度が上昇するにつれて減少し、沸点100℃で最小になる。25℃の水では、気化に必要なエネルギーは44kJmol-1で、これは液体の中では知られている最高の値である。
この熱エネルギーの大部分は、水分子間の水素結合を切るために使われる。潜熱は、蒸発する水分子の温度を変えずに、水の蒸発した表面の温度を下げるエネルギーである。例えば、水が蒸発するとき、つまり液相から気相に変化するとき温度が変わらないままで熱を吸収して蒸気となる。これを気化熱または潜熱という。同様に固相から液相に変化するときも熱を吸収する。これに対して相の変化が逆の場合、温度変化はなく熱を発散して相変化するがこれもまた潜熱という。液相から固相に変化して溶湯が凝固する場合、やはり熱を発散するこれを凝固潜熱といい、単位は(J/kg)である。水のこの高い気化潜熱は、太陽からの入射熱によって上昇しがちな葉温を、蒸散によって抑えるのに役立つ。
目次へ
液体はすべてその表面をできるだけ小さくしようとする傾向を持つ。空気―水界面にある水分子は、水素結合によって隣り合う水分子と引き付け合っている。この相互作用は、隣接する気相とのどのような相互作用と比べても、より強いものである。これは、液体の表面に沿って一種の張力が働くためであり、液体の表面をある線によって2つの部分に分けると、2つの部分は互いに引合うが、この力は線に垂直であり、単位長さあたりの力を表面張力surface tensionの大きさとする。もし空気―水界面が曲面なら、表面張力は、境界面に垂直な力が生じる。表面張力は熱力学的に考えると、2つの互いに混ざり合わない液体の境界面にも存在する。このような一般的な場合では、界面張力という。水は同じ温度で比較すると、他の液体に比べて非常に高い表面張力を持つ。液体の表面張力は、一般に温度の上昇とともに減少する。
液体の20℃での表面張力(N/m)は、1%ゼラチン;0.0083、エタノール;0.0228、フェノール;0.0409、水は0.0728と大きい。
この結果、最も安定した低いエネルギー状態で、空気―水界面の表面積を最小にすることになる。空気―水界面の面積を増やそうとすれば水素結合を切らなければならず、そのための エネルギーの供給が必要になる。この空気―液水界面の表面積を増やすのに必要なエネルギーは、表面張力として知られている。
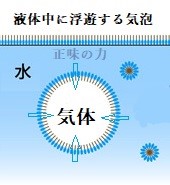 液体内に浮遊する気泡は、表面積が最小になるように球形になる。その表面張力は、気体―液水界面の接線に沿って働き、結果的に生じる正味の力は、気体の中心へ内向きとなり気泡を圧縮する。
液体内に浮遊する気泡は、表面積が最小になるように球形になる。その表面張力は、気体―液水界面の接線に沿って働き、結果的に生じる正味の力は、気体の中心へ内向きとなり気泡を圧縮する。葉内の水の蒸発部位で生じる表面張力と付着は、植物の維管束を通して水を引き上げる力を生じる。植物の葉の気孔から光合成に必要な二酸化炭素を吸収し、できた酸素を排出する。同時に、気孔から水が絶えず蒸散する作用により、導管内の水柱を引き上げようと働く。
水分子間の水素結合は、凝集cohesionという水の静電的に引き合う凝集力となる。導管のような毛細管内では、200気圧位の力があると言う。それと関連する付着adhesionも、水素結合形成により、細胞膜やガラス表面のような固相表面に水を引き付ける。水が固相表面に引き付けられる力と水同士の凝集力は、接触角contact angleを計測することで定量できる。接触角は、空気―水界面の形と表面張力が、液体中の圧力に与える影響を表している。
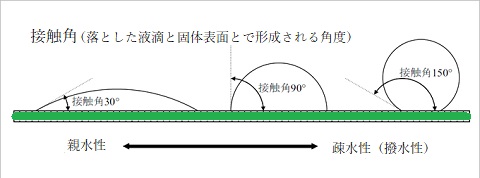 固相表面の液滴の形状は、液体が個体に吸引する力と液体自身に吸引する力との相対的な差で決まる。固相表面と気体―液体界面との間の角度として定義される接触角(θ)は、この相互作用の説明に使われる。濡れがよい(親水性)表面では、接触角は90°未満となる。きれいなガラスや一次細胞壁など、より親水性が高くなれば接触角は0°近くになる。葉の上の露のように、濡れにくい(疎水性)表面では、接触角は90°を超える。
固相表面の液滴の形状は、液体が個体に吸引する力と液体自身に吸引する力との相対的な差で決まる。固相表面と気体―液体界面との間の角度として定義される接触角(θ)は、この相互作用の説明に使われる。濡れがよい(親水性)表面では、接触角は90°未満となる。きれいなガラスや一次細胞壁など、より親水性が高くなれば接触角は0°近くになる。葉の上の露のように、濡れにくい(疎水性)表面では、接触角は90°を超える。凝集や付着と表面張力は、毛管現象を引き起こす。濡れた内壁を持つ毛細管を水中に垂直に立てると、平衡状態では毛細管内の水位は、管の外側の水位より高くなる。ガラス毛細管内での水位は、管の外側の水位より高くなる。ガラス毛細管での、水位の上昇は、極性を持つガラス管内への水の誘引(付着)、及び空気―水界面の表面張力によって起こる。
凝集や付着と表面張力は、一体となって水分子を引っ張り上げ、上に向かう力が水柱の重さにつりあうまでガラス管内を上昇する。管が細ければ細いほど、水柱は高く上昇する。
根で吸収された水は導管に入り、根圧で少しは押し上げられる。幼植物では、毛細管現象だけでも導管の中の水は問題なく上部の葉まで連続して到達するが、成長して大木になると毛細管現象だけでは間に合わない。しかし、水の凝集力は毛細管である導管内ではより強力に働き、大木でも根から頂上まで、連続した水柱にできる。しかも、昼間は上部にある葉の気孔の蒸散により、水は絶えず引き上げられ、導管内を切れることなく上昇する。
目次へ
水素結合は、水に高い引っ張り強度tensile strengthをもたらす。水の引っ張り強度とは、1 本の連続した水柱を引っ張るとき、水柱が分断される直前に水にかかる最大の力と定義され、単位面積当たりの力として表される。これは、毛細管中の水を引っ張り上げるのに必須となる性質である。水の引っ張り強度は、シリングに水を入れて考えればいい。プランジャーを押し込むと、水は圧縮されて正の静水圧hydrostatic pressureを生じる。水や油の中での圧力は殆ど静水圧と考えてよい。液体の中で均等に働く圧力は、その面に垂直に働き、面に沿う方向のずれは生じていないようだ。これは気体―液体界面の表面張力に起因する内向き力と同じ方向に作用する。このためシリンジ中の小さな気泡は、圧力が増すにつれ小さくなる。
逆にプランジャーを引っ張れば、水分子が引き離されることに抵抗して張力、あるいは負の静水圧が発生する。分子が互いに誘引する力を持つときにのみ、陰圧がかかる。液体による気泡への外向きの力が、気体―液体界面の表面張力に起因する内向きの力を超えると、気泡は大きくなる。
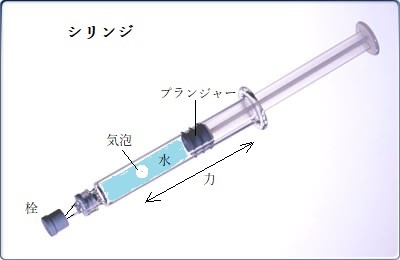 液体であるため、個々では弱い水分子間の水素結合であるが、水分子の立体構造から、隣接する4つの水素結合の連結により、非常に強い分子間相互作用が働き、水分子間で多元的な張力となり、巨大な被子植物相を育てた。
液体であるため、個々では弱い水分子間の水素結合であるが、水分子の立体構造から、隣接する4つの水素結合の連結により、非常に強い分子間相互作用が働き、水分子間で多元的な張力となり、巨大な被子植物相を育てた。これが気体分子間となれば、その相互作用は弾性衝突時に限られるため負の圧力は生じない。弾性衝突elastic collisionとは、衝突の前後で、2物体の力学的エネルギーが保存されている衝突を言う。物体の変形、熱の発生、内部エネルギーの変化などによりエネルギーを失う非弾性衝突に対し、完全弾性衝突とも言う。
シリング内の水柱は、非常に小さな気泡であるため、大きな張力には耐えられない。プランジャーを引っ張り続ければ、気泡は膨張し、シリングをなおも引っ張れば引っ張り抵抗力は低下する。
水分子の結合が引き裂かれ水柱が分断されるのに、プランジャーをどのくらいの強さで引く必要があるか?水は20MPa(メガパスカル)以上の張力にも耐えられることが研究で分かった。
車のタイヤは約0.2MPaまで空気を入れるという。家庭の水道管の水圧は、多くの場合、0.2~0.3MPaと言う。水深10mの水圧は、0.1MPaである。
シリング内の水柱には、非常に小さな気泡があるため、そのような大きな張力には耐えられない。プランジャーを引っ張ると気泡は膨張し、水柱の引っ張り抵抗力は低下する。
周囲の液体の張力による気泡の膨張は、キャビテーションcavitationとして知られている。キャビテーションは、木部の水輸送に致命的な影響を及ぼす。コーラのビンに見られる発泡は、栓を抜くことによってビンの中の圧力が下がったことによって起こる。この減圧による発泡現象を一般にキャビテーション (cavitation)と呼ぶ。
植物は木部のキャビテーションの影響を、いくつかの方法で最小限にする。木部内の水輸送経路は、互いに連絡しているので、原理的には、1つの気泡が広がってすべての仮道管を空気で満たしてしまう可能性がある。
実際には、膨張する気泡が壁孔膜の小さな孔を容易に通れないので、遠くまでは広がならない。木部の毛細管は互いに繋がっているので、1つの気泡で完全に水の流れが止まることはない。実際に、水は遮断された道管に隣接する水に満たされた道管を通って迂回する。木部中の仮道管と道管の長さに限りがあり、水の流れの抵抗を増すが、それがキャビテーションの影響を制限する手段となる。
気泡を木部から除去することもできる。植物の中には、木部に陽圧(根圧)を生ずるものもある。そのような陽圧は気泡を収縮させて水に溶け込ませる。最近の研究では、木部の水が張力を受けている際にも、キャビテーションが修復されていることが分かった。その修復の構造は未解明であるが、活発な研究課題になっている。
多くの植物は、二次成長をする。毎年、新しい木部を形成する。それにより、植物はキャビテーションによる水輸送の欠損を回復する。
水や無機塩類を根から運ぶ木部を構成する細胞群を管状要素と言う。全ての維管束植物に見られる仮道管(tracheid)と、主に被子植物に見られる比較的短い導管要素(vessel element)の2種類がある。いずれも成熟した段階で死細胞となる。導管要素は両端で繋がり、最大0.7mmほどの幅を持つ円柱状の導管(vessel)を作る。
一次細胞壁のみを持つ原生木部細胞が管状要素へ成熟する過程で、セルロースが螺旋状に沈着し、リグニングにより強化された二次細胞壁ができる。
細胞伸長が停止すると、細胞の長軸の上端と下端の細胞壁に大きな穿孔が作られる。細胞の側面では、壁孔以外の部分で二次細胞壁の肥厚化が進む。もともとは、隣り合った細胞間での情報交換を仲介し、タンパク質・核酸などの様々な巨大分子の径路として働いてきた原形質連絡(plasmodesma)が密集していた領域に形成された壁孔は、次第に隣り合った細胞を結ぶ細胞壁中の通路となる。
原形質連絡は、多細胞植物体の細胞間の細胞壁を貫く細い細胞質の膜となり、これにより細胞間の連絡通路と言われる。
植物細胞は、原形質連絡を介して、互いに繋がった連続体を形成している。この連続体をシンプラスト(symplast)と呼ぶ。細胞質と通路の中心を通るデスモ小管と呼ばれる小胞体由来の膜が管を満たし、隣接する細胞間の小胞体と繋ぐ。シンプラストを通した細胞から細胞への分子の移動を可能にする。
太さ40~50㎚前後で、両細胞の細胞膜がこれを通じてつながって細い管をつくり、その中を細胞質が通っている。通路の隙間の大きさは、その内側表面に並んだ球状タンパク質によって制御されている。ウイルスと同じ大きさの粒子までは通すことができる。物質とくに高分子物質の流通に役だっていると考えられる。仮道管と導管要素は、プログラム細胞死により細胞死を起こす。これにより硬い二次細胞壁からなる、かつ周囲の細胞と壁孔と連絡した管の束が残される。
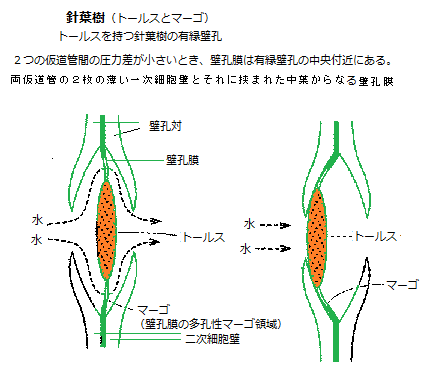 針葉樹では、仮道管の壁孔膜は中心が顕著に肥厚しており、トールス(torus)と呼ばれる。その周りには、多くの隙間を持ってトールスと上下の細胞壁を繋ぐセルロース微繊維構造のマルゴ(margo)がある。
針葉樹では、仮道管の壁孔膜は中心が顕著に肥厚しており、トールス(torus)と呼ばれる。その周りには、多くの隙間を持ってトールスと上下の細胞壁を繋ぐセルロース微繊維構造のマルゴ(margo)がある。壁孔膜の大きな直径と多くの隙間を持つマルゴは、多孔性で比較的柔軟で水を効率よく通すことができる。厚く肥厚したトールスは、バブルのように働き、壁孔対の空間の中央にあれば、壁孔は開孔している。
隣接した仮導管でキャビテーションが起きると、壁孔膜を挟んで圧力差が生じて、壁孔膜が水の入っている仮導管の側へ素早く移動する。その円形あるいは楕円形の厚くなった部分が壁孔に移動して、入り口を塞ぎ気泡の侵入を防ぐのだ。こうした有縁壁孔の構造は、通水の効率と、危険な気泡が隣の仮道管に広がるのを防ぐという安全性を同時に実現している。
ごく少数の例外はあるが、他の全ての植物の仮道管や導管要素の壁孔膜には、トールスがない。針葉樹以外の壁孔膜の水に満たされた孔は、極めて小さいので、それが気泡の移動の障壁となる。このような両タイプの壁孔膜が、気泡の拡散を防ぐ。木部内の気泡による水流の遮断を「塞栓」と呼ぶ。
細い管の中に液体が流れるためには、途切れない液体の流れが不可欠である。万が一、管に気泡が入り空気塞栓ができても、壁孔を通して隣の管に流し変えて塞栓部分を迂回できる。
植物は、蒸散により水を失い続けるため、脱水を防ぐ仕組みを進化させた。 植物は、木部キャビテーションの影響を、いくつかの方法で最小限に抑えている。木部中の水輸送通路は、互いに連絡されているので、原理的には1つの気泡の広がりで、全ての仮道管が満たされる可能性がある。だが、膨張する気泡は、壁孔膜の微細な孔を通過できないので広まらない。 仮道管は、長く伸びた中空の紡錘形で、高度に木化した細胞壁を持つ死んだ細胞である。垂直に重なり合う配置である。細胞壁は、細胞どうしを接着している細胞間層に、薄い一次壁と厚い二次壁が堆積してできており、壁孔はこの二次壁が欠如した部分、一次細胞壁のみにある非常に小さな領域である。植物細胞の細胞壁の一部で、隣り合う細胞同士の水分などの通路となる壁の薄い部分をいう。水は、この細胞壁の側面にある多くの壁孔を通って、仮道管同士の間を流れる。1つの仮道管の壁孔は、多くの場合隣り合う仮道管の壁孔と向かい合うように配置されている。これを壁孔対(pit pair)と呼ぶ。壁孔対は、仮道管間の移動抵抗の少ない水の通路を構成する。隣り合う壁孔対に挟まれた、水を通す多孔性の層は、両仮道管の2枚の薄い一次細胞壁とそれに挟まれた中葉からなり、壁孔膜(pit membrane)と呼ばれている。 また、木部の毛細管は互いに繋がっているので、1つの気泡で完全に水の流れが止まることはない。しかも、水は遮断された道管と隣接する、水で満たされている道管を使って迂回できる。木部の仮道管と道管の長さに限界があるため、水の流れに抵抗を与えるが、逆に、それが、キャビテーションの影響を回避させることに繋がっている。
木部から、気泡を除去することもできる。植物の中には、木部に陽圧(根圧)が生ずるものもある。陽圧は気泡を収縮させて、その気体を水に溶け込ませる。また多くの植物は、二次成長を行い、毎年新しい木部を形成するが、道管を新しくすることによって、キャビテーションによる水輸送機能の欠点を回避している。
最近の研究では、木部の水が張力を受けている際も、キャビテーションが修復されていることが観察されている。 葉から大気へ流出する過程で、水は木部から葉肉(ようにく)細胞の細胞壁の中に引き込まれる。細胞壁から蒸発して葉の空気間隙に入る。その水蒸気は、気孔を通って葉から出ていく。
目次へ