| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生
花粉の形成と構造
| 目次 |
| 1)花粉の発生 |
| 2)花粉粒の多層構造 |
| 3)動物の減数分裂 |
| 4)植物の減数分裂 |
1)花粉の発生
植物と動物における有性生殖の最も大きな違いは、植物の生活環では、雌性配偶体female gametophyteと雄性配偶体male gametophyteと呼ばれる2つの完全に独立した単相世代の個体が存在することである。厳密に言えば、花自体は生殖の組織ではない。花は雌雄の配偶体を含み、両配偶体が被子植物の真の生殖の組織を作る。
動物では、胚発生の過程で体細胞系列と生殖系列とが分かれ、この生殖系列の細胞が広い意味で生殖細胞とされる。一方、植物では、胚発生の過程で生殖細胞が隔離されることはない。植物では、ある細胞が配偶子を作ることが約束された段階で生殖系列が決まる。つまり植物の生殖系列は2段階で決まる。
小胞子嚢の詳細な発生過程は、植物種で異なるが、第1段階では、植物の成長が生殖成長に切り替わると、シロイヌナズナでは、成熟した葯は、胞原細胞archesporial cellを内包する。胞原細胞とは、雄性の花粉母細胞(小胞子母細胞)、または雌性の大胞子母細胞に分化する細胞群を言う。
花粉母細胞となる胞原細胞は、4種の体細胞性の細胞層である表皮・内皮・中間層・タペート組織に囲まれている、小胞子母細胞である。その胞原細胞を含む中心部を葯室loculeと呼ぶ。

花粉は葯室で2つの連続した過程により生じる。雄性配偶体である花粉の発達は、小胞子形成と小配偶子形成の2つの過程に分かれる。小胞子形成の過程では、葯室内の胞原細胞が小胞子母細胞に分化する。小胞子母細胞は、花粉母細胞とも呼ばれるように、減数分裂により小胞子を形成する二倍体の細胞である。その花粉母細胞の減数分裂によって、半数体の小胞子からなる花粉四分子を形成する。その葯室を取り囲む一層の分泌細胞からなるタペート組織tapetumは、加水分解酵素であるカロース分解酵素や他の細胞壁を分解する酵素を葯室へ分泌する。これら作用で花粉四分子が4つに分離し、各小胞子へと遊離する。
野生種のシロイヌナズナは、個々の小胞子に分離するが、マメ科ネムノキ亜科のアカシア属のように、さらに多くの花粉粒が集合するので多集粒polyadと呼ばれている。酸性土壌を好み、耐寒性はあるが、高温多湿に弱いギョリュウモドキ(カルーナ)の花粉は、四分子のまま送粉される。
小胞子が4つに分離しようが、花粉四分子や多集粒のまま接着していても、葯室内で小胞子が形成されると、小配偶体の発生過程における小胞子形成は完結する。
 第2段階は、雄性配偶子の形成、即ち小配偶子形成である。やがて、細胞壁合成や巨大液胞の形成に伴う小胞子の膨張が生じる。同時に、小胞子の核が、その細胞壁の一端に移動し(小胞子の極性化)、その状態で極性化した小胞子polarized
microsporeは、半数体の体細胞分裂(花粉有糸第一分裂)を行う。この極端な非対称分裂により、大きさの異なる2つの細胞が生じ、大細胞の方は栄養細胞vegetative cell(花粉管細胞tube
cell)と呼び、小細胞の方は雄原細胞generative cell(雄性生殖細胞とも言う)と言う。当初、雄原細胞の方は、花粉管細胞に取り込まれたように、小胞子の細胞壁に付着したまま、半球状のカルロース壁で、花粉管細胞と仕切られている。そのカルロース壁が崩壊して、完全に雄原細胞は、花粉管細胞の中に含まれることになる。花粉特有の二重構造となり、これを二細胞花粉(二細胞期)と呼ぶ。小細胞の方の雄原細胞は、そのまま花粉管細胞に取り込まれた状態が続く。その位置で雄原細胞は、細長い形状か紡錘形となる。これにより、急速に伸長する花粉管の中で、雄原細胞が活発な原形質流動により、胚嚢にある栄養核まで移動ができるようになるようだ。
第2段階は、雄性配偶子の形成、即ち小配偶子形成である。やがて、細胞壁合成や巨大液胞の形成に伴う小胞子の膨張が生じる。同時に、小胞子の核が、その細胞壁の一端に移動し(小胞子の極性化)、その状態で極性化した小胞子polarized
microsporeは、半数体の体細胞分裂(花粉有糸第一分裂)を行う。この極端な非対称分裂により、大きさの異なる2つの細胞が生じ、大細胞の方は栄養細胞vegetative cell(花粉管細胞tube
cell)と呼び、小細胞の方は雄原細胞generative cell(雄性生殖細胞とも言う)と言う。当初、雄原細胞の方は、花粉管細胞に取り込まれたように、小胞子の細胞壁に付着したまま、半球状のカルロース壁で、花粉管細胞と仕切られている。そのカルロース壁が崩壊して、完全に雄原細胞は、花粉管細胞の中に含まれることになる。花粉特有の二重構造となり、これを二細胞花粉(二細胞期)と呼ぶ。小細胞の方の雄原細胞は、そのまま花粉管細胞に取り込まれた状態が続く。その位置で雄原細胞は、細長い形状か紡錘形となる。これにより、急速に伸長する花粉管の中で、雄原細胞が活発な原形質流動により、胚嚢にある栄養核まで移動ができるようになるようだ。成熟期に入ると花粉は、炭水化物・タンパク質・脂質を蓄積し、その後、活発に起こる花粉管の発芽・伸長に必要な代謝に備えていく。例えば、シロイヌナズナの花粉外被では、タペート組織由来のタンパク質・脂肪酸・ワックスや炭水化物に富んでいる。通常、花粉は葯壁の裂開で放出される。その花粉が柱頭に受粉して花粉管が発芽すると、有糸分裂を行い(花粉有糸第二分裂)、2つの精細胞を形成する(雄原細胞分裂)。しかし多くの植物では、雄原細胞がまだ葯の中にある時に、花粉有糸第二分裂を行う。これを三細胞花粉(三細胞期)と呼ぶ。いずれにしろ、2つに精細胞の形成により、成熟した雄性配偶体が発生して、小配偶子形成が完了する。花粉粒が成熟すると、特殊な細胞壁が形成され、最終的に、2つの精細胞により、被子植物特有の重複受精が行われる。
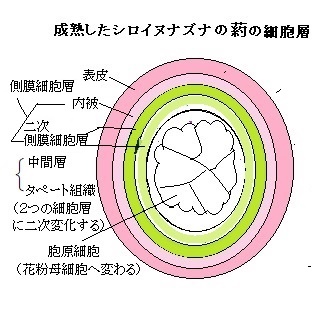 葯室の中に、胞原細胞からなる胞原組織 (ふつう1層の細胞層) が形成される。胞原細胞は並層分裂periclinal division(pèrəkláinl;分裂面が表面と平行方向となる分裂)をして、側膜細胞からなる側膜細胞層 (parietal cell layer) と、胞子形成細胞からなる胞子形成組織に分化する。 側膜細胞層が、内被・中間層・タペート組織に分化する。中間層は通常、短命で、花粉形成の過程で崩壊する。中間層の細胞はデンプンなどに富んでおり、花粉形成に用いられる栄養分を貯蔵しているようだ。側膜細胞層の最内層にあるタペート組織は、発生中の小胞子・花粉粒に接しており、花粉形成の際に栄養と原料の供給を行う。
葯室の中に、胞原細胞からなる胞原組織 (ふつう1層の細胞層) が形成される。胞原細胞は並層分裂periclinal division(pèrəkláinl;分裂面が表面と平行方向となる分裂)をして、側膜細胞からなる側膜細胞層 (parietal cell layer) と、胞子形成細胞からなる胞子形成組織に分化する。 側膜細胞層が、内被・中間層・タペート組織に分化する。中間層は通常、短命で、花粉形成の過程で崩壊する。中間層の細胞はデンプンなどに富んでおり、花粉形成に用いられる栄養分を貯蔵しているようだ。側膜細胞層の最内層にあるタペート組織は、発生中の小胞子・花粉粒に接しており、花粉形成の際に栄養と原料の供給を行う。シロイヌナズナのようにタペート細胞が、葯室周辺に残存する場合もある。あるいは、アメーバ状になり葯室内に進入して、発達中の小胞子と混じり合う植物種もある。タペート細胞tapetumは、花粉細胞壁の形成に関わる葯室を取り囲む分泌細胞層で、分泌機能を担い、最終的にはプログラム細胞死を引き起こして、内容物を葯室内へ放出する。それにより、タペート細胞は、発達中の花粉に酵素・養分・細胞成分を供給する。そのため、タペート細胞の障害は、通常、花粉発生の不全となり稔性が低下する。
シロイヌナズナやイネの花には、6つの雄しんがあり、それぞれの雄しんの上部には葯がある。葯は4つの葯室からなっている。葯室の中で減数分裂が起こり、花粉母細胞から小胞子が作られ、それが成熟して花粉粒が作られる。小胞子期の葯室は 、最も内側の小胞子から、タペート細胞、内被、そして 最も外側の表皮から構成されている。
 今日では、花粉母細胞やタペート細胞において発現している遺伝子が徐々に明らかになってきている。ただ、どのような遺伝子が、どの組織の形成や機能を制御しているのか、それ程までには、厳密に明らかになっていない。
今日では、花粉母細胞やタペート細胞において発現している遺伝子が徐々に明らかになってきている。ただ、どのような遺伝子が、どの組織の形成や機能を制御しているのか、それ程までには、厳密に明らかになっていない。ただ、SPL/NZZ (SPOROCYTELESS/NOZZLE) やTPD1 (TAPETUM DETERMINANT1) 遺伝子は、葯の発生に重要な役割を果たしている、SPL/NZZは花粉母細胞・タペート細胞・中間層・内被の発生を制御している、またTPD1はタペート細胞の分化に重要な役割を果たしている、ことまでは.、明らかになっている。
シロイヌナズナやイネでは、一次胞子形成細胞から花粉母細胞へと、一次側膜細胞からタペート細胞・中間層,・内被へと分化するが、例えば、タペート細胞の細胞壁のセルロースは、花粉母細胞が減数分裂する直前の時期に減少し、また同じ時期に花粉母細胞の細胞壁のセルロースも減少する。spl/nzz変異体の葯では、これらの細胞の発生が進んでも、花粉母細胞とタペート細胞の両方が分化しないため、 葯内のどの細胞もセルロース性細胞壁を失わないことが知られている。またtpd1変異体の葯においてはタペート細胞が分化しないが、花粉母細胞のセルロース性細胞壁は減少することも分かっている。このように、花粉形成の分子機構や、どの遺伝子がどの組織の形成や機能を制御しているのか、その着実な研究が進行している。
 多層構造の花粉粒の細胞壁は、特異な組成・形態をもち、その最外表面は、極めて複雑な層状構造を示し、花から花へ花粉が運ばれる生態学的構造に、進化の過程が読み取れる。そのためか、植物種ごとに特異なパターンが生じている。更に、表面下にある多層の細胞壁層も複雑で、脂質やタンパク質が沈着する内部空間は錯綜としている。
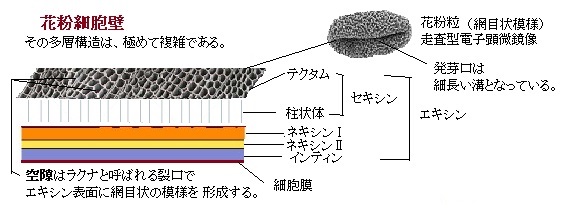
多層構造の花粉粒の細胞壁は、特異な組成・形態をもち、その最外表面は、極めて複雑な層状構造を示し、花から花へ花粉が運ばれる生態学的構造に、進化の過程が読み取れる。そのためか、植物種ごとに特異なパターンが生じている。更に、表面下にある多層の細胞壁層も複雑で、脂質やタンパク質が沈着する内部空間は錯綜としている。典型的な花粉細胞壁(sporoderm)の構造は、外層・内層と立体的要素で層状に構成されている。セキシンsexineは、エキシンの外層で、そのセキシンの底部層を構成するネキシンを加えた部分をエキシンと呼ぶ。セキシンの内層に柱状層があり、テクタム tectum(外表層)は、柱状体の頭部が融合したものを言う。全てが切れ目無く融合して花粉粒を覆っていることも (外表層型) 、一部に孔の開いた状態で取り残されて網目状になるもの (半外表層型) 、柱状体が独立しておりテクタムが形成されないもの(外表層欠失型)があり、特に外表層欠失型を非テクタム性intectateと言う。それに対してテクタムをもつものをテクタム性 (tectate) と言う。
模式図の花粉は、テクタム性で滑らかな表層構造を形成する。空隙はラクナlacunaと呼ばれる裂口で、エキシンの表面に網目状の模様を形成する。
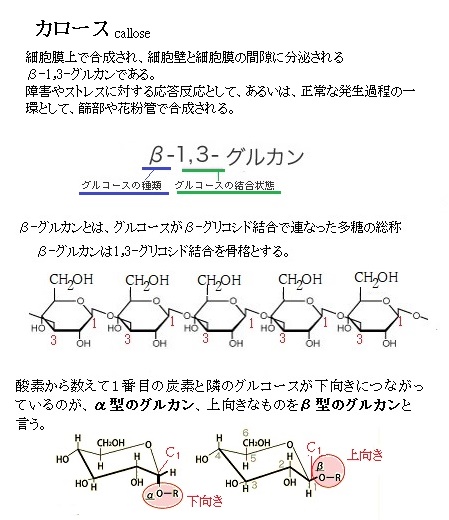
減数分裂後直ぐに、小胞子で花粉粒細胞壁の形成が始まる。カロースcallose(β-1,3-グルカンから成る、グルコースを重合した多糖)を主成分とする一過性の細胞壁は、小胞子で作られる細胞表面に沈着する数層の中で最初に形成される。続いて、カロース壁の内側に、セキシsexine(有刻層)の前駆体であるプライムエキシンprimexine(小胞子の表層に形成されるマトリクス層でその中でエキシンが発達する)とネキシンnexine(無刻層)が順次形成され、最終的にはインティンintine(内壁)が形成される。ということは、小胞子内部から小胞子の細胞表面へ、,外表層から上記の細胞壁層が段々に形成されるため、最も内側の層が最後に沈着される層になる。
インティンは、おもにセルロース・ペクチン質・カロースからなる。熱帯性のショウガ科やクスノキ科ではインティンが発達してエキシンが極めて薄くなっている。これは多湿環境への適応だと考えられている。
プライムエキシンの主成分は多糖である。フェノール酸や脂肪酸から構成されるスポロポレニンsporopollenin重合体の蓄積を誘導する鋳型として働く。スポロポレニンは、ネキシンとセキシンを含む外層構造であるエキシンの構造的主成分である。小胞子がまだ四分子であった時から、小胞子で合成され分泌されたスポロポレニン前駆体をもとに、初期のエキシンが構築される。ネキシンは、エキシンの内層で、化学的な違いから2層に分けられるものが多いが、均一で区分できない種もある。成分は、セキシンと同様、スポロポレニンを主とする。
ところが、最外層のカロース壁が分解されて、小胞子から四分子が遊離すると、スポロポレニン前駆体の大部分は、タペート組織から供給される。インティンintine(内壁)や内層壁は、主にセルロースやペクチンから構成される。
殆どの花粉壁には、発芽口apertureと呼ばれる細長い領域があって、シロイヌナズナの柱状体columellaの頭部が融合しているエキシンのテクタムtectum(蓋・天井の意味)構造では、その部分は薄いか欠失している。シロイヌナズナの花粉粒にある3つの発芽口は、細長い溝で、その細胞壁は薄くなっている。
テクタムは、柱状体の頭部が融合したもので、エキシンの内層であり、底部層の表面に直立した柱状の構造である。花粉壁において柱状体を欠くものを無テクタム性 atectateという。例えば、ニューカレドニア島の多雨林にのみ生育するアンボレラ科は、無テクタム性で、被子植物の中で最も初期に分かれた、わずか数百種のみが現生する基部被子植物である。モクレン科やモクレン目のバンレイシ科も、無テクタム性である。
テクタムには、柱状体の頭部が融合したものや、全てが切れ目無く融合して花粉粒を覆っていることもある。一部には、孔の開いた状態で取り残されて網目状になるものもある。
報酬として花粉だけを送粉者に与える花を花粉花pollen flowerと呼ぶ。チョウやガのように液体しか利用できない動物は、花粉花から報酬を得ることができない。その分、花粉花は送粉者の幅が狭くなる。進化的には、被子植物では花粉を提供する花は原始的であったと考えられており、スイレンやモクレンなど原始的な種では、花粉を報酬とするものが多い。スイレンの花粉はおわん型をしている。まわりにはねっとりした粘着物質が付着している。
コケ植物や維管束植物などで陸上植物であるものは、胞子は乾燥に耐えられるように,多層からなる厚い胞子壁でおおわれ、さらに固い表皮で包まれている場合が多い。胞子壁はスポロポレニンsporopolleninとよばれる物質からできている。「sporo」は、種子・胞子な「連結形」で、花粉の外膜はカロチノイドの重合体であるスポロポレニンという非常に硬い物質で、また内膜はセルロースとペクチン(多糖類)の重合体でできている。
トクサ植物の胞子の表面には胞子壁の肥厚によってできた2本の弾糸がまきつき、周囲の乾湿によってほどけたり、まきついたりするので、胞子の放出に有効である。古来、茎を煮て乾燥したものを研磨の用途に用いた。「とくさ(砥草)」の名はこれに由来している。現代でも高級ツゲグシの歯や漆器の木地加工、木製品の仕上げ工程などに使用されている。
スポロポレニンは、分解されにくい高分子物質で、アルカリにも酸にも不溶で、花粉や小胞子は、土中の酸素の少ない条件化でも、長年月、化石記録として保存される。
その体細胞は、カロチノイド系色素で保護され、外膜にまで直立した柱状層を持つものが多い。そのエキシンの特徴的なパターンにより、古代の気候変動やどの植物種が生殖していていたを推定できる、貴重な資料になっている。
化石で花粉が見つかることが多いのは、これは外膜のスポロポレニンが化石として残るためで、表面の網目や放射状の模様、また形などは花粉ごとに異なる事により植物種が特定できる。奈良の纏向古墳でも大量の花粉がみつかり、ベニバナと特定された。
昆虫で花粉を食料とするものがいる。花粉の成分を調べると、タンパク質が多く、16~30%、脂肪分は3~10%、デンプンを1~7%程度含んでおり、これら昆虫は、脂肪分を多く含む花粉を好むようだ。
昆虫と植物との共生関係は、地面に落ちている花粉を昆虫が食料とした事から始まったようだ。またミツバチの蜂蜜は赤から黄色まで色が異なるが、これは集めた花粉のカロチノイドの分子構造により生じる。
目次へ
配偶体において配偶子の形成へと進む細胞が決定される段階で、雄では胞原細胞が細胞分裂を行うなか、減数分裂へと進む細胞が多数生じる。生殖系列の決定には細胞の系譜よりもその位置が重要であり、この位置情報の重要性が、植物の発生において一般にみられる基本戦略である。
植物の生活環は、二倍体(2N)の胞子体世代と一倍体(1N)の配偶体世代と言う、2つの独立した多細胞世代からなる点で、基本的に動物の生活環とは異なる。植物は、動物と異なり、生活環の中で異なる2つの多細胞世代を交互に繰り返す。これを世代交代alternation of generationsと言う。そのうちの1つの世代は、複相diploid phaseの細胞からなり、それぞれの染色体を2コピーずつ持っている(2Nの染色体を持つ)。もう1つの世代は、それぞれの染色体を1コピーずつ有する単相haploid phaseの細胞(1Nの染色体を持つ)からなる。単相と複相の世代は、相互に循環する依存関係にあり、その依存度は系統ごとに異なっている。
二倍体生物の有性生殖に直接かかわる生殖細胞(germ cell)、即ち配偶子(gamete;gǽmi t)は、殆どの細胞と異なり一倍体であるため、ヒトの場合、各配偶子には23本の染色体が1組だけ.と.なる。しかも、殆どの生物は、雄と雌で異なる新たな遺伝子を含む配偶子を作り上げている。動物の配偶子は、大型で運動性のない卵と、小型であるが活発に運動する精子がある。その違いは、卵が大量の細胞質を抱えているが、精子にはそれが殆どないことによる。この特異な2種類の一倍体である配偶子が、融合して二倍体の細胞の受精卵、即ち接合子(zygote;zάɪgoʊt)となり、やがて新たな遺伝子を受け継ぐ個体を誕生させる。そのため2種類の配偶子は、染色体を正確に半分に分ける特殊な細胞分裂をして作られる。このような細胞分裂を減数分裂と言う。
減数分裂では、1回のDNA複製に続いて2回の細胞分裂が起こる。二倍体細胞が通常の細胞分裂で分裂する場合は、持っている2セットの染色体を前もって2倍にする。これにより、その細胞分裂で新たに誕生する2つの娘細胞それぞれが、完全な母方染色体1セットと父方染色体1セットからなる、ヒトの染色体セットを受け継ぐことになる。
減数分裂では、通常2セット分ある全染色体が、最終的に1セットになり、それが精子と卵からなる配偶子を作る。その前に起こる染色体倍化の現象は、通常の細胞と同様で1回である。生殖系列細胞は、その後独自の過程を経る。 DNAの複製を行なうことなく、連続して2回の細胞分裂が起き、むしろ最終的には、細胞内の染色体数を半減させて4個の一倍体細胞である配偶子を作る。生物40億年の進化の歴史は、苛烈な自然選択に、無駄なく合理的に応えられる生命体の存続しか認めなかったことを物語る。それ以後の種の存続のために不可欠な突然変異が必ず生じる仕組みを、進化として積み重ねる原動力にした。
生殖系列細胞の減数分裂は、通常の細胞分裂を少し変えていく。DNA複製を完全に省略すれば、二倍体細胞は細胞分裂を1回することで、2個の一倍体を簡単に作れるはずだ。しかし生殖系列細胞は、その後の減数分裂に特異な手順を踏ませた。それを理解するためには、ここでも生物40億年の進化の歴史の重みをわきまえる必要がある。
ヒトの減数分裂は、卵巣や精巣にある特定の二倍体生殖系列細胞で起こる。これらの生殖系列細胞は、体細胞と同じく二倍体で、それはそれぞれの染色体が父方と母方に由来のため、2対存在するようになる。生物学では、父方相同染色体、母方相同染色体と呼ぶ。減数分裂の最初の段階で、これらの染色体は、すべて倍加し2倍になった染色体コピー同士は、通常の細胞分裂の場合と同様に、互いに接着している。ところが、次の段階は、減数分裂に特有で、倍加したそれぞれの父方染色体がまず定位置に並び、その後、それに対応する母方の倍加した相同染色体が接着して対を作る。この対合pairingという過程のおかげで、相同染色体は、その後の2回の細胞分裂に際しても正確に分離し、最終的にできる配偶子それぞれに完全な一倍体の染色体セットが入ることができる。
減数分裂で2回連続して起こる細胞分裂、減数分裂第1分裂と減数分裂第2分裂で生じる4個の一倍体細胞それぞれには、完全な染色体セットが分配される。一倍体の娘細胞に父方と母方どちらの相同染色体が振り分けられるかはランダムなので、ここで生じる一倍体細胞からできる配偶子は、それぞれ父母の由来が異なる組み合わせの染色体を持つ。
つまり、減数分裂では、二倍体細胞1個から遺伝的に異なる4個の一倍体細胞ができ、それぞれの染色体は親生殖系列細胞の染色体数の半分になる。これが、通常の細胞分裂では、二倍体細胞1個から遺伝的に同一の二倍体の娘細胞が2個できるのとは異なっている。通常の細胞分裂と減数分裂は、数時間以内に完了するのが普通だが、生殖系列細胞の減数分裂第1分裂は、「倍加した相同染色体の対合」⇒「倍加した相同染色体の対が中期紡錘体に並ぶ」⇒「第1分裂後期に相同染色体が分離」といった前期Ⅰの段階が長いため、数日あるいは数か月、場合によれば数年かかることもある。
ヒトを含む複相の動物は、単相の配偶子gameteの卵子と精子が、複相の細胞の減数分裂meiosis(mɑɪóʊsɪs)により作られる。動物では、生殖細胞 (卵子または精子) 形成時に起る組換えは、核分裂が2回続き、第1分裂で相同染色体(母方および父方から由来した同じ形の一対の染色体を言う)の対合時に起こり、分離して染色体数が半減する。第2分裂は普通の核分裂で、結果としてもとの半数の染色体をもった4個の細胞ができる。
相同染色体の一方の染色分体のDNA鎖が切断酵素により切られ、もう一方の染色分体の相同な配列に入り込み、そこでDNA鎖の乗り換え(繋ぎ換え)を起こして染色体の一部が入れ替わりる。
先述するように、減数分裂第1分裂と減数分裂第2分裂で生じる4個の一倍体細胞それぞれには、完全な染色体セットが分配されるが、相同染色体が互いに、再度、接着する際に、数か所の接着点のうち、染色体の交換が起こった部位・キアズマchiasmaが生じる。しかし、一倍体の娘細胞に父方と母方どちらの相同染色体が振り分けられるかはランダムなので、ここで生じる一倍体細胞からできる配偶子は、それぞれ父母の由来が異なる組み合わせの染色体を持つ。この組換えにより、同一染色体に乗っている遺伝子の組み合わせが変化し、より多くの種類の配偶子を作り出すことが可能になる。その結果、子孫の遺伝子が多様に展開される。こうして、ヒトなどの複相(2N)の動物では、複相の細胞の減数分裂によって、単相(1N・卵や精子)の配偶子が作られる。
目次へ
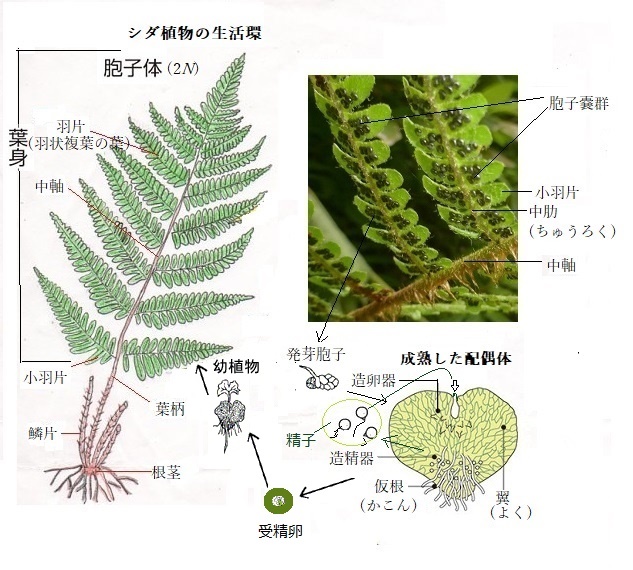 一方、複相の植物は、減数分裂を経て胞子sporeを形成する。そのため複相世代の植物は、胞子体sporophyte(spɔ'ːrəfàit)と呼ばれる。配偶子形成の際の細胞分裂は、体細胞分裂と似ているが、重要な違いがいくつかある。減数分裂の第一分裂では、それぞれの染色体が娘細胞に分かれる前に、相同染色体間でDNAが交換され、これによって遺伝物質の組み換え、即ち染色体交差(キアズマchiasma;kaiǽzmə)が、相同染色体間で複数起きている。これを「減数分裂組み換え」と言う。
一方、複相の植物は、減数分裂を経て胞子sporeを形成する。そのため複相世代の植物は、胞子体sporophyte(spɔ'ːrəfàit)と呼ばれる。配偶子形成の際の細胞分裂は、体細胞分裂と似ているが、重要な違いがいくつかある。減数分裂の第一分裂では、それぞれの染色体が娘細胞に分かれる前に、相同染色体間でDNAが交換され、これによって遺伝物質の組み換え、即ち染色体交差(キアズマchiasma;kaiǽzmə)が、相同染色体間で複数起きている。これを「減数分裂組み換え」と言う。減数分裂の第二分裂では、相同染色体それぞれから姉妹染色体が生じ、元の細胞1つから4つの娘細胞が生まれる。減数分裂は2回の分裂からなるが、DNAの交差複製は1回しか起きていない。つまり、4つの娘細胞は、元の細胞と比べて半分の遺伝子しかもたない。
その意味は、二倍体(2N)の植物体が、結果、半数体(1N)の配偶子を作る。これを減数分裂還元と言う。
2段階の減数分裂により生じた胞子は、有糸分裂mitosisを経て配偶体と呼ばれる単相の多細胞個体となる。有糸分裂では、2つの細胞に別れた中心体から放射する紡錘糸(微小管)は、赤道面に並ぶ動原体kinetochore(kiníːtəkɔ`ːr)を介して染色体と結合し、各染色体は、二つに縦裂される。分裂後期には、縦裂によって分かれた各染色分体が、それぞれの極に向かって移動する。終期には、両極に密集した染色体が、逆の過程を経て娘核となり核膜を形成する。
この染色体の数が母細胞と娘細胞で変化しない有糸分裂により、配偶体gametophyte(gəmíːtəfàit)という多細胞個体となる。
動物の配偶体が減数分裂により作られるのに対して、コケ植物やシダ植物などの配偶子は、単相の配偶体が、体細胞分裂をすることにより形成される。この点が動物とは根本的に異なる。雄性配偶子は、雄ずいの葯の中に作られ、雌性配偶子は胚珠の中に作られる。成熟するにつれ、雌雄配偶体の特殊化した細胞が、有糸分裂により分裂し、配偶子である精細胞や卵細胞を作る。
植物の生活環に一倍体の配偶体世代があるということは、植物の配偶子は減数分裂によるのではなく、体細胞の分裂を介して作られることを意味する。
「ハチが雌花に精子を運ぶ」のではなく、多細胞の配偶体である精細胞を作る花粉を運ぶのである。花粉は、胞子体の受容組織に運ばれて発芽する。花粉は、柱頭から花粉管を伸長させ、胞子体の組織内の雌性配偶体へ侵入し、精細胞を放出する。これが卵との受精である。受精の最終段階では、卵細胞と2つの精細胞の中の1つが生殖融合を行う。
動物では減数分裂により生じた細胞は、配偶子である卵と精子をその流れのまま分化する。植物の方では、減数分裂により生じた細胞(1N)は、体細胞分裂をして多細胞の組織としての配偶体を作り、そのなかで配偶子が分化する。
動物⇒減数分裂→配偶子(卵および精子)の分化→受精
植物⇒減数分裂→多細胞からなる配偶体の形成→配偶子の分化→受精
ひとたび、単相の配偶子同士が融合し、受精fertilization(受胎;肥沃化・多産化の意味がある)となって2Nの受精卵になれば、その後の発生過程は動物と類似しており、2Nの受精卵は、有糸分裂を繰り返して胚を形成して、最終的には植物体へと成長する。すべての植物の生活環は、複相で胞子を形成する胞子体世代sporophyte generationsと単相で配偶子を形成する配偶子世代gametophyte generationsを繰り返す世代交代alternation of generationsをする。受精と減数分裂が、植物の世代交代の節目となる。
受精から減数分裂までの間に起こる体細胞分裂の回数が増大する植物体ほど、胞子体世代のサイズが大きくなる。それにより、作り出される胞子の数も増加する。一度の受精ごとに形成される胞子が増えれば、やがて胞子が置かれる、乾燥などの環境リスクから、種が保存できる機会を大いに向上させられる。植物の進化の過程で、胞子体世代が次第に大きくなるのも、これを裏付ける。
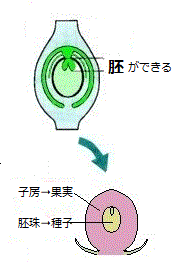 植物の受精戦略は、その過程で不可欠な水と時間が限定される環境条件下で、効率よく繁殖を行うためのものである。砂漠に降った雨に、素早く反応して、植物が一斉に芽吹き、花を咲かせ、水が干上がる前に、種子を作り次の世代へと種を繋げようとする。これが被子植物の繁殖能力を支える生活環である。
植物の受精戦略は、その過程で不可欠な水と時間が限定される環境条件下で、効率よく繁殖を行うためのものである。砂漠に降った雨に、素早く反応して、植物が一斉に芽吹き、花を咲かせ、水が干上がる前に、種子を作り次の世代へと種を繋げようとする。これが被子植物の繁殖能力を支える生活環である。現生種として約760種が知られているイチョウやスギなどの裸子植物の生殖方法は多様である。それはまるで、種子を作る植物における、様々な進化の過程が、その生きた化石により克明に観察される。ところが、約25万種もある被子植物には、ここで説明する重複受精とよばれる仕組みを共有している。重複受精の仕組みを獲得した被子植物は、爆発的な繁栄を誇っている。
被子植物には、特有な様式の配偶子融合が起こる。受精の最終段階で、卵細胞と2つの精細胞の中の1つが生殖融合を行う。2つ目の精細胞は、雌性配偶体内の二倍体の中央細胞と融合して、三倍体の「一次胚乳組織」を作り、それが養分に富む種子の胚乳組織となる。被子植物特有な、2つの精細胞が受精に関わる受精様式を、「重複受精」と呼ぶ。
陸上植物の系統発生によれば、親植物から独立して自由生活する一倍体の配偶子が中心の発生様式から、二倍体の胞子体が、自由活発な生活環へと進化して行く。
多くの被子植物の雄性の配偶体が花粉である。雌性の配偶体を胚嚢(はいのう)と言う。それぞれ、葯と胚珠(はいしゅ,受精前の種子)で作られる。花粉が雌しべの先端の柱頭に付着する受粉により、受精の過程が始まる。
配偶体には、受精に深く関わる、雌性の4細胞、雄性の3細胞、あわせて7つの細胞がある。雌性の配偶子として「卵細胞」と「中央細胞」の2つ、雄性の配偶子として“精細胞”が2つ存在している。卵細胞は片方の精細胞と受精し胚を作る。中央細胞も、独自に受精して胚乳という胚の栄養器官を作る。卵細胞のように次世代に遺伝情報を伝えることはできないが、受精する細胞であることから配偶子のひとつとみなされている。卵細胞と中央細胞の受精を合わせ重複受精と呼ぶ。
一方、生殖細胞のほかには、精細胞を運ぶ雄性の細胞として花粉管細胞(栄養細胞とも、核は栄養核とも呼ばれる)が1つと、その花粉管とのやりとりを担う雌性の細胞として助細胞が2つ存在する。花粉管細胞は、細胞の一部が先端成長をして、内在している精細胞を運ぶ。助細胞は、胚嚢に花粉管を誘引し、その先端を破裂させ精細胞を放出させる。それにより、助細胞は崩壊する。死をもって受精を助ける、献身的な役割を担う細胞である。これら7つの細胞がそれぞれ重要な役目を果たして、雌しべの組織(2N)の力を.借りながら組織的に受精を達成する。
目次へ