| Top 車山高原 車山高原お知らせ 車山ブログ
|
|

DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物
| 目次 |
| 1)キク科とは |
| 2)キク科の頭花 |
| 3)キク科の花の構造 |
| 4)キク科の繁殖力 |
逆に花粉媒介に特定の昆虫や動物を必要としない風媒花などでは、花弁は退化傾向にあり目立たなくなる。特に、栄養生殖や自家受粉、あるいは無配生殖(アポミクシス)で次世代が作られるようになるとなお更である。
キク科の植物では、多数の花が集合して、花としての機能を分担しあっている。進化の一方の頂点にあるキク科から、多数の花を持つ被子植物を中心に、植物は進化してきた。和名のキクは漢字の菊の音読みで、菊の字は鞠とも書かれ、鞠は窮と同じ意味で窮極または最終の意味という。キクの花が、年のいちばん終わりに咲くことから、この字があてられたようだ。
野生のキク属の植物は、通常は多年草で、茎は木質でも冬には枯れるが、低木状となって茎が木化して冬も枯れないものもある。葉は単葉で裂けないものもあるが、浅くあるいは深く羽裂して、いわゆる菊の葉状になる。両面とも毛は少ないが、裏面に白色の綿毛を密生するものもある。
キク科は、南極大陸以外、世界のあらゆる地域に分布している。種子植物の中では、最も種の数が多く、形態的にも変化に富んでいる。成育地は海浜から高山の雪線(せっせん;一年間の降雪量と雪の溶解量が一致する点を結んだ線)まで広い。林縁・道端・草原・海岸の砂浜・砂漠など直射日光を好むようだ。典型的な陽生植物と言える。そのため森や林の日陰では、殆ど見られない。また、キク科は、乾燥に耐えるように進化してきたようだ。キク科の90%以上は多年草で、レタスやシュンギクなど約7%が一年草である。約3%が木本で、高木もある。
日本にもコウヤボウキのような、木本とされているキク科植物がある。高野山で茎を束ねて箒の材料としたのでこの名がある。通常、コウヤボウキを木本とは呼ばない。
キクモンタノア属ツリーデイジーは、 ハワイオアフ島のタンタラスの丘の林縁や低木林中に、4~5cmの白い花を咲かせている。近寄ってみると日本の野菊のような花であり、葉はクワとかコウゾのようだ。木なのでツリーデイジーと呼ばれている。ハワイでの外来の植物で、旺盛な生育によって在来種が駆逐されかねないので、有害植物に指定している。森林破壊があった場所では、遷移初期に侵入する。
ツリーデイジーは、冬に幹の先端に多数の花を咲かせることからクリスマスデイジーと呼び、生育状態からブッシュデイジーとも呼ばれる。メキシコ南部からグアテマラ・コスタリカ・コロンビアなどが原産地である。高さは数mになり、電柱にするほどである。
東南アジアのベルノニア・アルポレアは、十数mになる高木であり、小笠原のワダンノキ(海菜木)は、小笠原諸島の固有種で、葉は肉厚の常緑で、大きいものは樹高3~ 5m、幹の直径は10cm以上もあり、しかも群生する。
キクを仏花として当たり前のように供えるのも、庭先や畦地に長く咲き続け簡単に手折れるし、乾燥に強いため花もちがよいからである。現代のように仏花を、近くのコンビニで簡単に購入するすべもなかった。
目次へ
 キク科の花の大まかな特徴は、密に集まった多数の小花群と、その基部を包み込んでいる、葉状の総包片とで、頭花を構成していることにある。頭花が1つの花にみえるのは、その基部に輪状に取り囲むように集まったもので、1枚ずつの包(包葉;ほうよう)が、がくのように見えるからだ。
キク科の花の大まかな特徴は、密に集まった多数の小花群と、その基部を包み込んでいる、葉状の総包片とで、頭花を構成していることにある。頭花が1つの花にみえるのは、その基部に輪状に取り囲むように集まったもので、1枚ずつの包(包葉;ほうよう)が、がくのように見えるからだ。頭花は、頭状花序(とうじょうかじょ) とも言う。 散形花序(さんけいかじょ)は、セリ科の特徴で、この散形花序の花柄(かへい)がとれて、花軸(かじく)の上が平らになった花床(かしょう)の上にならんだのが頭状花序と呼ぶ。頭状花序は、花のもっとも進化した形といえる。 多数の花が円形に集まって、一つの花の形を作るものである。頭花を以て一つの花だと考えがちであるが、小さな個々の花びらに見えるのが、それぞれが1つ1つの花である。
それぞれに雄しべや雌しべがあり、小さいながらも花の構造を持っているのが分かる。ヒマワリは、1,000個を超える小花(しょうか)を持つ花序である。
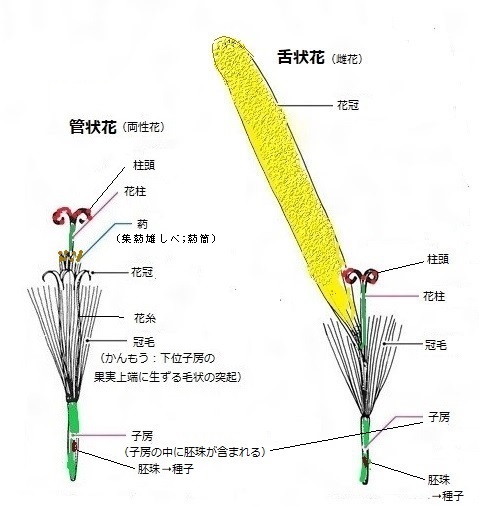
頭花をつくっている小花には、筒状花(とうじょうか)または管状花(かんじょうか)と、舌状花(ぜつじょうか)の2種類がある。筒状花も舌状花も、アサガオのようにラッパ形の花の花弁(一つの花の一枚一枚の花弁)が全部くっついているのを「合弁花」と言う。もともと何枚かあった花弁が隣同士でくっついてしまった状態、例えばアサガオの花弁の先は五角形であるが、もともとは5枚の花弁がくっついたと考えられている。
ところが、キクや・タンポポ・ヒマワリも合弁花で、筒状花の花弁の先端部分だけが5つに裂けているが、先端部以下では、5枚の花弁が合弁して一体化している。ただ、5枚の花弁は、たいへん小さい。舌状花では、その花冠(一つの花の花弁全体)の一方だけが伸び、伸びていない方は、Vネックになっている。
ヒマワリの小花自体、筒状花と舌状花があり、筒状花が中心花(心花)となり、中心花を取り巻く周辺花(辺花)が、筒を切り開いたような舌状花を伸ばし、頭花の外周を巡っている。総苞片の変異も加わり、頭花の多様性は実に豊かである。
キク科の頭花を構成しているのは、中心花となって密に集まった多数の小花と、それを支える下部周囲の総苞片である。
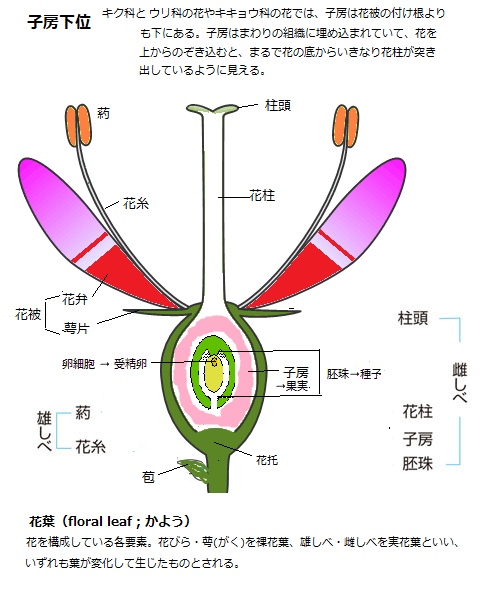
その小花は、すべて合弁花で、その生殖器官の中核となる子房は花被perianth(萼と花冠を総称)の下にあるため、いわゆる子房下位(しぼうかい)と呼ばれる。更に小花自体、心花と辺花に分化し、生物の宿命であるが、その総苞片にも変異が加わるため、頭花の巧妙な進化は、更に多様性が豊かに広がった。そのため、キク科は、種子植物の中では最も種類が多く、しかも最も進化した科となった。
目次へ
 小花は、雄しべと雌しべが揃う両性花と、雄しべが退化した雌花と、雌しべが不稔となった雄花と、雄しべも雌しべんも退化した中性化がある。
小花は、雄しべと雌しべが揃う両性花と、雄しべが退化した雌花と、雌しべが不稔となった雄花と、雄しべも雌しべんも退化した中性化がある。通常、筒状花は両性の心花であり、舌状花は雌花か中性の辺花となっている。
キク科のノアザミの筒状花から飛び出した雄しべには、花粉が付いている。周辺花として咲き進んだ筒状花では、さらに長く細い糸状の花弁が多数出ている。これが雌花である。ノハラアザミは、極めて細いピンクの花弁が多数伸びている。それは糸状花(しじょうか)と呼ばれる雄しべを欠いた雌花である。
小花の構造は、筒状花で見ると、中心に先端が2裂した雌しべが1本あり、その周囲5本の雄しべが取り巻いている。雄しべは縦に長い葯と細い花糸からなり、葯は合着(ごうちゃく)して筒状になっている。これを集葯雄しべ(しゅうやくー)と呼び、その筒状を葯筒(やくとう)と言う。
頭花は小さいものは径1cm、大きいものは3~5cm、中心に管状花、周囲に舌状花があり、両方とも結実するが管状花のほうが稔実率が高い。
開花前の小花が咲く順序は、常に外側の辺花から心花の中央部へと、求心的で、最初に、葯筒の内側の葯室が裂けて花粉が出される。開花と同時に、花筒の下部にあった雌しべ伸長する。その際、雌しべの先端にあるブラシ状の毛で、花粉を葯筒上部に押し出す。その押し出された花粉は、訪花昆虫により他の花の雌しべに運ばれる。
これは自家受粉を避けるのに都合がよい。これらの小花の構造と花粉放出の構造は、キク科の重要な特徴であり、キク科の進化の証明でもある。
キク科ツワブキ属の常緑多年草ツワブキの頭花は、舌状花と筒状花の2種類の小花で構成されている。舌状花は雌花で頭花の外周に1列に並び、筒状花は両性花で中央に多数集まっている。舌状花は雄しべを欠いた雌花で、花冠は筒を切り開いたような形状となっている。花冠の先端部はタンポポのような5枚の花弁が合着した痕跡はない。
雌しべの柱頭は通常2裂しているが、まれに完全に3裂しているものも見る。開花期の初期段階では、周囲の舌状花のみが雌しべを見せているが、舌状花に取り囲まれる筒状花はほとんど開いていない。筒状花は両性花で、花冠の先端が5裂し、下部は合着している。中心に先端が2裂した雌しべが1本あり、それを5本の雄しべが取り巻いている。筒状花は、外周側から次第に開花している。合着して筒状となった、褐色がかった雄しべの葯筒の先から、葯室が裂けて花粉が出ると、これら雄しべの下方から閉じた2裂の雌しべの柱頭が出てくる。
雌しべが外へ押し出してくる。伸び出た柱頭部分で、丸味のある先端部は無毛で、その直下は特に長い毛が見られる。閉じた柱頭の外側の上向きの毛が、ブラシとなって花粉を捉えて押し出し、閉じた柱頭の先端の毛が花粉にまみれる。
やがて、中心部を占める一部の筒状花以外は、殆ど開花する。この段階では、中央寄りの筒状花は、まだ雌しべが出ていない。やがて、雌しべに押し出されて、ふっくらと盛り上がった花粉の様子が見られる。葯筒の先端から姿を見せた雌しべの柱頭が開き、雌しべの柱頭が完全に2裂すると、他の筒状花からの花粉を受粉する状態になる。
キク科の特徴の一つで、その果実は多くの被子植物に見られる痩果(そうか)で、イネの種子 (籾) などのように小さな乾いた果実で、果皮は硬くて裂開せず、中は1種子のものもある。
鱗片状(りんぺんー)や芒状(のぎー)、棍棒状(こんぼうー)、または鱗片状と剛毛状を二重に持つものなど様々で、冠毛(かんもう)を持たないものもあるが、多くは冠毛を持つが、あってもごく短い。冠毛は、キク科植物のタンポポ・アザミやスイカズラ科のツルカノコソウなどの下位子房の果実上端に生ずる毛状の突起で、萼が変形したもの、果実が熟したあと、乾燥して放射状に開き、風を受けて飛び、種子を散布するのに役立つ。 タンポポでは、ごく軽い綿毛のような冠毛が、種子を遠く運ぶが、アザミなどの冠毛は、痩果から簡単に脱落して風に乗る。
目次へ
 日本の代表的な花であるキクは、中国の唐時代に品種改良が盛んになり、そのキクが日本に伝来したようだ。
日本の代表的な花であるキクは、中国の唐時代に品種改良が盛んになり、そのキクが日本に伝来したようだ。菅原道真が、宇多天皇の命を受けて、詩文などの古典の中から抜粋し分類再編集し、寛平4 (892) 年、天皇に奉ったといわれる『類聚国史(るいじゅこくし)』に、宮中で催された宴の時に、桓武天皇が詠んだ歌が載っている。
「このごろの しぐれの雨に 菊の花 散りぞしぬべき あたらその香を」
キク科は世界に広く分布し、双子葉植物では最も進化・分化したため、1,100~1,200属に分類されるが、2万3,000を超える種があるため、キク亜科とタンポポ亜科の2亜科に分けられる。
それでも、属の数が多いので13連(動物では「族」)に分けられる。
その分類の特徴としては、
①乳管の有無
②頭花の形
③葯室下部付属体の形
④柱頭の形
⑤冠毛の有無と形
などである。
 タンポポ亜科は、タンポポ連だけである。そのいずれの植物体に共通するのが、乳管が発達し、葉や茎をちぎると白い乳液が出ることである。
タンポポ亜科は、タンポポ連だけである。そのいずれの植物体に共通するのが、乳管が発達し、葉や茎をちぎると白い乳液が出ることである。植物の乳管とは、乳液を分泌する細胞が集まってできている管で、タンポポのみならず、他のキク科のレタス・ニガナ・ノゲシ・アキノノゲシなどや、キキョウ科(キキョウ・キキョウソウ・ホタルブクロ・ツリガネニンジン)などに見られる。
乳液は、植物体内を血管のように発達した乳管と呼ばれる特別な管状構造の中を流れる。例えば、乳管には、直列に融合した細胞列(有節乳管)や、細胞融合した1つの長い細胞(無節乳管)からなるものなどがある。この管状構造を切断すると、気体にさらされると凝固する乳白色の液を滲出する。その乳液には、タンパク質・糖質・脂質・アルカロイド・テルペノイドなどが含まれており、植物種によってその主成分は違う。この液体は、ラテックス(latex)と呼ばれることが多い。樹脂と比べ、その構成成分は複雑で、毒性または忌避性の二次代謝産物を含むことも少なくない。ラテックス生産植物には、その乳液が弾性ゴム(生ゴム)、つまり天然ゴムの原料となるインドゴムノキが有名である。自然環境下では、インドゴムノキが傷を付けられると、それと同時に、膨大なラテックスは放出される。その分子量は100万までになり、植食性昆虫や動物のみならず、病原菌を寄せつかなくするか、包み込んで防御をする。
サポヂラの乳液がチューインガムの基材チクル、またケシの未熟果実の乳液がモルヒネの原料となっている。ケシの乳液に含まれるラテックスには、高濃度の麻酔剤、特にモルヒネとコデインを含む。これを摂取すると、草食動物の神経系で、オビオイド受容体と結合し鎮痛効果を発揮する。
キョウチクトウは、かなりの量のカルデノリドを含む、透明なラテックスを非有節乳管から放出する。この有毒なステロイドの活性は、強い経口毒性があり、高濃度で心臓停止を引き起こす可能性がある。腐葉土にしても1年間は毒性が残るため、腐葉土にする際にも注意を要する。枝を箸代わりに利用し、中毒した例がある。
カルデノリドは、脊椎動物の脳で、嘔吐を誘発する神経中枢を活性化させる。広食性の植食性昆虫では、この物質に反応して忌避するか、あるいは痙攣死するかである。
多様なテルペノイドには、植食性昆虫や動物による食害に、直ちに反応して放出される樹脂酸も含まれている。一度放出されると、それらは有毒となり、植食性昆虫を攻撃し、昆虫の口の部分を接着させる粘着剤となったりする。極端な事例としては、その樹脂が昆虫や病原菌全体を巻き込み、効果的に死滅させている。
また、タンポポ亜科の小花は、すべて両性の舌状花であり、性や形態において心花と辺花の分化が見られない。 キク亜科は、12連からなり、乳管は発達していない。すべての小花が両性の舌状花ではなく、しばしば辺花に舌状花が見られるだけで、そのいずれも雌花か中性花である。
タンポポ連(ニガナ属・クンショウギク属など70属)・オナモミ連(ブタクサ属など8属)・メナモミ連(201属)・シオン連(ヒメジョオン姫女菀など113属)・キク連(ノコギリソウ属・ヨモギ属・キク属・シュンギク属・カミツレなど82属)・ハゴロモギク連(ガザニアなど16属)・オグルマ連(ハハコグサ属・ヤマハハコ属・エゾノチチコグサ属・ムギワラギク属など183属)・サワギク連(コウモリソウ属;モミジガサ・ツワブキ属・フキ属・ギヌラ属;スイゼンジナなど94属)・キンセンカ連(キンセンカ属・オステオスペルマム属・カレンデュラ属など8属)・ヒヨドリバナ連(カッコウアザミ属、フジバカマなどのヒヨドリバナ属、ステビアなど62属)・ショウジョウハグマ連(ルリギク属・ショウジョウハグマ属など62属)・アザミ連(ノハラアザミ・トネアザミ・ハマアザミなどのアザミ属、ゴボウ属、オケラ属、ヒレアザミ属など)・コウヤボウキ連(ガーベラ、センボンヤリ属など69属)の13連である。
限定された領域内に生息する1属1種から、1個体あたり47,000以上の種子を生産し発芽率も高い、しかも種子の寿命が35年と長いなど、進化の最高水準に達する繁殖能力を持つヒメジョオンは、全世界を席巻する種である。
1回の繁殖で、大量の種子や果実を生じ、侵入した先々で、在来の植物との競合を避けるため、乾燥や強い日差しにも耐えられる生活環をもっている。この典型的な陽生植物は、強い光を有利に利用する代謝系を持ち、全体が毛に覆われ日中の乾燥への適応にも優れている。しかも、冬季だけロゼットの状態で越年し、数か月、あるいは数週間という短さで次世代を作れる。
ヒメムカシヨモギ(姫昔蓬)の大株1個体が生産する痩果の総数は、約81万9,000個で、この個体が発芽し結実するまで180日と言う。多産で知られるスベリヒユの24万4,000個をはるかに超えるものである。
キク科の植物は、人間や貨物・航空機・自動車などの移動に伴って容易に運ばれる散布体となりうる種子や果実を持ち、そのため散布体が急速に拡散し、しかも農作物の種子に混入しやすい。その繁殖においても、特定の昆虫に花粉の媒介を依存していない。栄養生殖や自家受粉、あるいは無配生殖apogamyで次世代が作られる。
キク科の花は、基本的に虫媒花であるが、花の構造はそれほど特殊化しておらず、特定の昆虫だけに依存することはない。このため、新天地に帰化し繁殖するのに有利に働く。キク科のブタクサなどは風媒花である。ヨーロッパ原産のセイヨウタンポポは、受粉を必要としない無配生殖によって雌しべだけで結実する場合もある。
キクイモやハルジオンでは、地下部分だけで繁殖する栄養生殖も行う。 ヒメジョオンやヒメムカシヨモギは、北アメリカ原産で、明治維新のころ渡来した。通常、秋に芽生えてロゼットを形成し、そのまま越冬して、生育が好条件となる翌年の初夏から夏に花茎を伸ばして開花する。生育期間は1年未満であるので一年生草本である。
日本のキク科植物は、72属370種余りが知られる。属では、キク科全体の6%、種では僅かに1.6%だけ、日本に生息する。
目次へ