| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達
| 目次 |
| 1)細胞のシグナル伝達 |
| 2)植物のシグナル伝達 |
| 3)概日リズム |
| 4)フィトクロムとクリプトクロムが時計を同調させる |
| 5)コルチゾールcortisol受容体 |
個々の酵素は、特定の分子の化学反応を触媒する。その酵素が触媒として働く反応の連続で代謝経路ができる。その触媒作用によって、細胞は、生存・成長・増殖に必要な全ての化学反応の総和である代謝を厳密に調節できるので、これこそが生命化学の中核となる。
細胞内で起こる化学反応には、反対に向かう2つの流れがある。一つは、食物を分解する径路「異化」であり、細胞が使いやすい形態のエネルギーや、細胞の構成材料として欠かせない小分子の一部を提供する。
もう一つは、異化作用によって蓄えられたエネルギーを用いて、細胞構築用の分子を生合成する「同化」径路である。両者を合わせて細胞の代謝という。
生物が周辺環境の変化に対応するように、細胞も周囲の状況変化に対応する。細菌ですら、栄養物を見つけ、明暗を感知し、毒物や捕食者を識別して回避する。
細胞も何らかの「社会生活」を営むには、他の細胞との連携が不可欠である。例えば酵母細胞は、接合の準備が整うと接合因子を分泌する。この化学物質による、呼びかけを感知して異性の酵母細胞(異なる接合型の細胞)は、細胞分裂を中止し、シグナルを出して細胞に向けて突起を出して呼応する。
出会うと、抱き合って細胞同士が融合する。この過程を「接合」と呼ぶ。雌雄の体が溶け合って融合した細胞は遺伝子を混ぜ合わせて、新しい遺伝子の組み合わせをもつ次世代の細胞を作る。こうして母細胞の中に4つの子供細胞ができる。
酵母の異性細胞の融合から、遺伝子の新しい組み合わせを持つ個体を生み出すことから、高等生物の有性生殖に基本は同じである。酵母は無性生殖と、有性生殖の二つの異なる方式で子孫を増やしていく。
酵母の一倍体にはa細胞(MATa)とα細胞(MATα)という2種類の接合型(性)が存在し、接合因子(Mating Factor)を分泌する。言い換えれば、酵母には、オスとメスにあたるプラス型とマイナス型があり、それぞれに特有の接合因子であるフェロモン分子を細胞外に発散する。酵母の接合因子をα-Factorとa-Factorと呼ぶ。α-Factorはa細胞で、a-Factorはα細胞で作用する。
それを感じた異性の細胞は刺激されて、その方向へ突起を出して接合への準備態勢を作る。プラス型とマイナス型の酵母は、それぞれ相手のフェロモンを見分けて結合できる「受容体」というタンパク質が細胞の表面にある。
つまりフェロモン分子は受容体に認識されて、細胞を発情させることができる。細胞外シグナル分子であるフェロモンを特異的に識別して応答する受容体タンパクは、鍵と鍵穴の関係のように、厳密な相性があり、異なるカギでは錠前は開かない。
多細胞生物では、もっと複雑で、細胞が互いに協調するには、細胞外から受け取る多くの様々なシグナルを解釈しなければならない。例えば動物の胚発生では、胚の細胞間でシグナルを交換し合い、それぞれの役割や体組織での位置を決め、生存と分裂のみならず死をも決める。
その後の生活でも、動物は、膨大な数のシグナルの共働により、成長・生理機能・行動などを調節する。植物でも、細胞は常に互いに連絡し合い、その細胞間の相互作用を調節し、根・茎・葉などの様々な組織を構成していく。やがて広範で複雑なシグナル伝達のネットワークによる相互作用を活用して行動を調節していく。
植物は、移動できない。好ましくない状況下でも、耐え抜くために、常に環境に応じた調節を行っている。そのために、植物は水分や栄養分の利用を最適化するシステムや、光の量や質及び方向性を検知するシステム、更に生物学的・非生物学的な脅威を防御するシステムを発達させた。
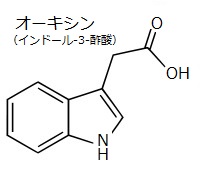 チャールズ・ダーウイン(Charles Darwin)と息子のフランシス・ダーウィンは、草花が日光を浴びると、光を感じて茎や根が生長の方向を変えるという先駆的な研究を行った。ダーウィン親子が、光に反応する幼葉鞘の屈折時におけるシグナル伝達の観察で、一方向から照射された光は、幼葉鞘の先端で受容されるが、実際に屈折するのは、それよりも下の領域であることを見出した。この発見でダーウィン親子は、1880年に、幼葉鞘のある領域から別の領域に移動可能なシグナルが存在し、屈曲を起こしていると結論した。
チャールズ・ダーウイン(Charles Darwin)と息子のフランシス・ダーウィンは、草花が日光を浴びると、光を感じて茎や根が生長の方向を変えるという先駆的な研究を行った。ダーウィン親子が、光に反応する幼葉鞘の屈折時におけるシグナル伝達の観察で、一方向から照射された光は、幼葉鞘の先端で受容されるが、実際に屈折するのは、それよりも下の領域であることを見出した。この発見でダーウィン親子は、1880年に、幼葉鞘のある領域から別の領域に移動可能なシグナルが存在し、屈曲を起こしていると結論した。1931年に、この移動性のシグナルが同定され、ギリシャ語で成長を意味する「オーキシン」と名付けられた。これが最初に発見された植物ホルモンである。
一般に、植物の反応を引き起こす環境因子をシグナルと呼び、そのシグナルに生化学的反応をする実体を受容体と言う。受容体はタンパク質であるが、光受容体の場合は、タンパク質に結合した色素である。
受容体が自分に特有な特異的なシグナルを検知すると、シグナルを増幅し細胞応答を引き起こすため、そのシグナルを伝達するため変換する必要がある。受容体は他のタンパク質の活性を変化させたり、細胞内のシグナル分子であるセカンドメッセンジャーsecond messengerを用いたりしてこれを行う。これらの分子は、遺伝子の転写などの細胞内プロセスを変化させる。
多くの場合、最初の反応は、ホルモンなどのシグナルとなる分子の生産であるが、それが作用部位に輸送され過程で主要な生理反応を引き起こす。その植物におけるシグナル伝達の間に生じる様々なイベントや中間段階が明らかになってきている。それら中間産物がシグナル伝達径路signal transduction pathwayを構成する。
シグナル伝達径路は、遺伝子の転写を阻害する抑制因子(リプレッサーrepressorタンパク質)の不活性化・分解・除去によって機能しているとは驚くべき発見であった。植物全体の反応を調和させるシグナル伝達のメカニズムとともに、セカンドメッセンジャーを介したシグナルの増幅が重要である。
目次へ
植物の環境応答のタイミングと同様、シグナル伝達機構も、非常に速いものから、極めて遅いものまで様々ある。ハエトリソウなどの食虫植物が、変形した葉で昆虫を捕獲する際に、その葉の捕虫器の上で昆虫が動くと、刺針が接触刺激を受けて活性化され、葉はミリ秒の瞬息で閉鎖される。
モウセンゴケの葉は、昆虫を触毛と呼ばれる長く突き出た有柄腺(ゆうへいせん)によって生産される粘性の高い粘液で捕獲し、巻き上げて獲物を確保し、葉に張りついたような赤い無柄腺(むへいせん)で消化を始める。
オジギソウは、接触後、直ちに葉は垂れ下がり、小葉は重なり合って閉じる。その後、徐々に元に戻る。オジギソウやネムノキは、無駄な蒸散を防ぐため、光合成をしない夜には葉を閉じる。
発芽した芽生えは重力を感じ、茎は上方へ根は下方へ成長する。植物を宇宙空間の微少重力環境で育成すると、根は色々な方向に伸びると言う。
このような迅速な反応機構には、電気化学的な応答がシグナルの伝達に関与している。その都度、遺伝子の転写やタンパク質の翻訳を介していては遅すぎる。
昆虫の食害を受ける植物は、数時間で昆虫の天敵を誘引する揮発性物質を放出する。栄養分の状況に応じた根の分岐、光の状態に対応する陽葉と陰葉の成長、茎頂が捕食された場合の腋芽(えきが)成長の活性化など、これらの事象では、多くの場合、新たな転写と翻訳を必要としている。より長期的な環境変化には、個々の植物は、数か月から数年に及ぶタイムスケールで応答するが、むしろ、植物は、遺伝子の発生プログラムを変化させ、一生を通して植物の体制を形作っているようだ。
植物の葉には陽葉と陰葉があり、弱光下では、陰葉の方が光合成速度は高いが、強光下では、陽葉の光合成速度は、陰葉より格段に高くなる。そのため強光下の植物の葉は陽葉であり、弱光下の地面に近いところにある植物では陰葉であることが多い。また強光下の方が陽葉で、弱光下の環境では陰葉であるといった個体差だけでなく、同一個体で2種類の葉を持つものもある。
最初から、強光下では陽葉、弱光下では陰葉とするのは、難しいようで、まずは、陽葉のみを先に全体に茂らせ、落葉し、次の葉を茂らすときに、光の当たらないという情報をもとに徐々に陰葉へシフトしていったのではないかと考えられる。また世代が代わるごとに、過去の情報が細胞に受け継がれ陰葉を茂らすなど、個体全体の葉を調節し、光合成速度を最大化にするため効率よく光を利用している。光環境を感知する要は、前年の葉の環境が当年の葉の性質を決めていることにある。
弱光下では、陰葉の光合成速は陽葉を超えるが、強光下で生育した陽葉でも、例えば天候が曇りの時など、弱光下に置かれることは当然である。実際には弱光下での光合成速度は陽葉よりも陰葉の方が速いことから、陽葉は強光下での速い光合成速度と引き換えに何かを犠牲にし、逆に陰葉は、弱光下での光合成速度向上のために、強光下での光合成速度を犠牲にしていると考えられている。
弱光下での光合成速度を増すためにはクロロフィル含有量を増すことが必要であり、また、最大光合成速度に達するためには、電子伝達系のタンパク質・ATP合成酵素・カルビンーベンソン回路(大気CO2を有機物へ取り込む代謝回路)の酵素を増やすことが必要である。葉が保有することのできるクロロフィル量、タンパク質量は限られているため、陽葉と陰葉は、置かれた環境に合わせて一方を増やし、もう一方を減らしているとみられる。陽葉はその体積を増やすことでクロロフィル含有量を増やしたとしても、光合成系タンパク質の含有量がそのままであれば、光合成速度は高まらない。それで、弱光下では、陽葉は陰葉の光合成速度を超えることが不可能になる。
環境シグナルに応答する植物の細胞自律的反応(cell autonomous response)では、シグナルの受容と反応が同一細胞で起こる。非細胞自律的反応(non-cell autonomous response)では、シグナルの受容と反応が、離れた細胞や組織、あるいは器官で起こる。
細胞自律的なシグナル伝達には、気孔の開閉がその例である。青色光は、青色光受容体タンパク質であるフォトトロビンを介して膜のイオン輸送体を活性化し、孔辺細胞を膨張させる。同じ器官における非細胞自律的なシグナル伝達では、例えば成熟した葉が強い太陽光にさらされた場合、新たに気孔を形成する。これらが、ある器官から別の器官へ、シグナルの伝達がなされる必要性の証明となる。
非細胞自律的反応の典型例が、シュート頂分裂組織が花の形成を決定する花成惹起(floral evocation)に見られる。植物の生活環で最も重要なのが、「いつ花を咲かせるか」ということである。この決定を遅らせれば、篩管を通して転流に回すことができる炭水化物を増し、その成熟過程で、種子により多くの貯蔵栄養分が蓄えられる。また、より多くの種子も作ることができる。
その一方で、花成の遅れは、食害と環境変化のリスクを伴う。また、繁殖前に他の植物に繁殖領域を奪われかねない。そのため植物は、多岐にわたる繁殖適応を進化させてきた。一年草と多年草の生活環もそれであろう。
ノボロギクはヨーロッパ原産の一年草で、今では世界中の温帯に分布している。日本には明治初頭に帰化した。葉はシュンギクに似た形である。花は真冬にも咲き、一年中見られる。このノボロギクは、発芽後数週間で花を咲かせる。
シロイヌナズナの花成応答は、ある環境刺激によって促進されるものの、そのような環境シグナルがない場合でも起こる。このシロイヌナズナの花成応答は、条件的であると言う。
環境要因のうち、春化vernalizationと光周期photoperiod(日長)が最も重要なものである。多くの植物では、冬に経験する長期間の低温が花成の促進には重大で、春化と呼ぶ。光周期とは、昼と夜の長さに対する応答であり、光質や外気温、非生物的なストレスなどの他のシグナルも植物の発生を促進する重要な外的信号刺激となる。
植物や動物は、通常、日々の明と暗のサイクルにさらされている。共にそうした変化に関連した周期的な振る舞いを示す。葉や花弁による昼の姿勢と夜の姿勢、気孔の開閉、菌類(ミズタマカビ・アカパンカビ)の成長、蛹から羽化の時間帯、光合成や呼吸といった代謝過程など、その日周変動があげられる。
生物の24時間の明・暗サイクルから、恒常的な暗あるいは明条件に移した場合、そのようなリズムの多くは、少なくとも数週間は持続する。そのような一様な条件下にあるリズムの周期は24時間に近く、そのため、ラテン語の「概ねcirca」と「日diem」に由来する「概日リズムcircadian rhythm」と呼ばれた。 その後に起こる明あるいは暗条件の変化にも、数週間は持続することから、こうした概日リズムは、光の有無に対する直接的な応答ではありえず、内的な時計の分子機構に依存していると見られる。これらは、内生振動子endogenous oscillatorと呼ばれることが多い。 植物の様々な生活活性は、ほぼ24時間の周期(概日)の振動を示す。この内生リズムは、外界からの毎日の明暗周期に同調する振動子の働きによる。
目次へ
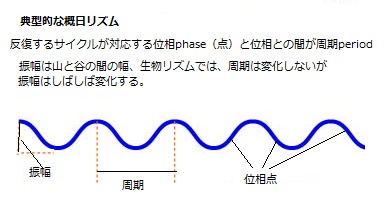 (位相phaseとは、時間経過とともに最大値、最小値がある律動的周期現象において、全過程の周期との関係が認識できる周期上の任意の位置を指す。)
(位相phaseとは、時間経過とともに最大値、最小値がある律動的周期現象において、全過程の周期との関係が認識できる周期上の任意の位置を指す。)概日リズムの生理的応答は、内生振動子の特定の時間点と結びついているため、応答は一日の中の決まった時刻に起こることになる。しかし、単一の振動子は、位相が異なる複数の概日リズムと連結されているが、大きく北半球と南半球の違いがあり、それに伴う季節の移り変わりに合わせて、日々の明期(昼の長さ)と暗期(夜の長さ)を調節しながら、生理応答を定刻通り行っている。
概日リズムとは、約24時間の周期で生命現象が繰り返されることをと言う。
ハムスターは夜行性なので、暗い時間帯に活動し明るい時間帯に休息する。1日の長さを24時間に保ったまま、照明の点灯と消灯の時刻を人工的に変えてやると、生物時計は活動と休息の時間を照明の点灯と消灯の時間帯に合わせようとする。これを同調という。その同調のきっかけとなる事象を同調因子という。
その同調因子が働かない暗中の部屋で何日も過ごすと、約24時間の周期で活動と休息を繰り返すようになる。これは、生物時計の独自の周期が行動に表れるからで、これを自由走行と言う。このような生物時計の周期の約24時間は、正確な24時間ではない。概日リズム(circadian rhythm)とは、概(おおむね)日(じつ、一日)のリズムの意である。
概日リズムの研究者は、まずは生物の恒常的な暗条件で、概日リズムを自由走行させる。その自由走行リズム(周期的変動)の位相点で、通常は1時間以内の短い光パルスPulse(電磁波)を与え、それに対する反応を調べる。それが、はじめ12時間明、12時間暗というサイクルに同調していたものが、恒常的な明あるいは暗状態と自由走行リズムに移されると、自由走行リズムの位相で、以前の同調化サイクルにおける明期に対応する位相を、主観的な昼subjective dayと呼び、暗期に対応する位相を主観的な夜subjective nightと呼ぶ。
恒常条件の環境下では、約24時間周期で昼の状態と夜の状態が交互に起こる概日リズムが現れる。その昼の時間帯を主観的な昼、夜の状態を主観的夜という。昼(夜)行性動物で、明(暗)期開始と同時に活動を始める種類では、恒常条件下の概日時間circadian timeは24時間で、それが概日リズムの1サイクルとなる。活動開始時を起点として半分に分け、前半(概日時間の12時間)を主観的な昼(夜)とし、残りを主観的な夜(昼)とする。
この条件下で、光パルスが主観的な夜の最初の数時間の間に与えられた場合、リズムの位相に遅れが生じる。生物はその光のパズルを夜に先立つ昼の終わりと解釈するからである。連続暗条件に移って数時間の間に与えられた光パルスに対して応答する典型的な位相シフトである。この遅れに位相はシフトトするが、その概日リズムは維持される。
一方、主観的な夜の終わり近くに与えられた光パルスを、生物は次に来る昼の始まりと解釈し、リズムの位相をシフトする。
これにより、季節の進行により昼夜の長さが変わると、概日リズムは、それに合わせて適切な時を刻み続ける。概日時計は、こうした位相をシフトさせる応答によって、概日リズムを、様々な長さの明期と暗期からなる、約24時間のサイクルに同調させる。
位相シフト応答によって、植物が日長の季節変化に合わせて、概日リズムを調節させることが明らかになった。
ヒトばかりかマウスなどの小動物でも、老化が進むと、概日時計の性能も衰えると、概日リズムの中でも、睡眠覚醒リズムの「振幅」が小さくなる。そのため、老年期に入ると、夜間は寝つきが悪く、夜尿症のせいばかりでなく何度も目が覚め、昼間は幾たびも睡魔に襲われる。24時間周期の概日時計に狂いが生じている。
目次へ
光シグナルが位相のシフトを引き起こす分子機構は、未だ解明されていないが、シロイヌナズナでの研究から、特定波長の弱い光で、位相のシフトを引き起こせるのは、この光応答が、光合成反応とは異なる、特異な光受容体によって仲介されているとことまでは解明されている。
シロイヌナズナは5つのフィトクロム(phytochrome)を持ち、その内のフィトクロムCを除く全てが概日時計の同調に関わっている。個々のフィトクロムは、赤色光(せきしょくこう)・遠赤色光・青色光(あおいろこう)に特異的な光受容体として作用する。
植物は、フィトクロムとともに、クリプトクロム(CRY)を介して光を受容して、CRY 1とCRY2と呼ぶタンパク質が、青色光による概日時計の同調に関わる。クリプトクロムは、胚軸伸長の抑制・子葉展開の促進・細胞膜の脱分極・葉柄伸長の抑制・アントシアニンの蓄積・概日時計の同調などの幅広く青色光応答を制御する青色光受容体である。
クリプトクロムが時計の同調に関わることは、昆虫や哺乳類でも同様である。クリプトクロムは、赤色光による通常の同調にも必要とされている。しかし、クリプトクロムは、赤色光を吸収しないため、CRY 1とCRY2は、概日時計の同調過程で、フィトクロムの情報伝達において仲介役として働いているようだ。シロイヌナズナでは、フィトクロムとクリプトクトムが、共に概日時計に関わることを示している。
植物では、光により活性化されたCRY2が、鍵となる花成遺伝子FLOWERING LOCUST(FT)の発現を直接的に速めることで、青色光に応答して花成の促進に関わっていることが示されている。
樹木では、花をつけ始めるまでに20年、あるいはそれ以上も稀ではない。笹は、短いものでは10年足らずで開花結実するが、おおよそ50年と見られている。マダケ(真竹)の開花時期は初夏であるが、その周期は約120年で、ほぼ間違いないとされている。
自律的な調節系と環境を感知する調節系の両方が進化したことにより、植物は花成を正確に調節し、それを繁殖に導く最適な時期に種子を形成する。特定の種子の多くの個体群は、花成を同調させて外交配(他家受粉;異なった形質をもつ血縁関係の遠い個体同士の交配)を有利に仕組んでいる。環境からのシグナルに応答した花成は、特に水と温度を重要視し、その好条件の下で確実に種子が発芽できるよう計算されている。
目次へ
 細菌では、一群の遺伝子の発現を協調させるのに、1個のプロモーターが制御する1つのオペロン(同一オペレーターによって同時に調節される一組の構造遺伝子)に複数の遺伝子をまとめる仕組みがある。
細菌では、一群の遺伝子の発現を協調させるのに、1個のプロモーターが制御する1つのオペロン(同一オペレーターによって同時に調節される一組の構造遺伝子)に複数の遺伝子をまとめる仕組みがある。真核細胞では、すべてをまとめる遺伝子がなく、遺伝子は個々に転写され、個々に転写調節因子共働体が働いている。その場合、遺伝子発現における全体の協調はどうなっているのだろうか?
実は、遺伝子の発現が組み合わせによって調節されていても、1個の転写調節因子が、遺伝子のオン・オフの決定権を握っている。
個々の遺伝子の活性あるいは抑制に必要な組み合わせを、この因子が最後に完成させていた。異なる遺伝子でも調節に必要な組み合わせの最後に同じタンパク質が使われていることもある。
異なる遺伝子であっても、同じ転写調節因子を識別する調節DNAが含まれてさえすれば、この因子によって、ひとまとめに発現をオン・オフにできる。
原核生物や真核生物のいずれも、その細胞は、個々にオン・オフするだけでなく、異なる遺伝子の発現が協調されなければならない。そのため、真核細胞が「分裂せよ」というシグナルを受信すると、それまで発現しなかった多数の遺伝子が活性化し、細胞分裂を誘発する。しかし、各遺伝子の調節配列に、異なった遺伝子活性化因子が結合するだけでは、それらの結合タンパク質の転写後の活性化効率は悪いので、もう1つの別の転写調節因子、即ち活性化したコルチゾール受容体複合体が、各遺伝子すべての調節配列に結合する。
その別の転写調節因子である1個のタンパク質が、異なる遺伝子の発現を協調させている。実は、遺伝子の発現には、今まで発現していなかった多数の遺伝子の組み合わが一斉に働かなければならない。そのため、1個の転写調節因子が、ある遺伝子群のオン・オフの決定権を握り、それぞれの遺伝子の活性化とその抑制に必要な組み合わせを、この因子が最後に完成させる。
異なる遺伝子でも、調節に必要な組み合わせのために、最後に同じタンパク質を使っていれば、その転写調節因子を識別する調節DNAさえ含まれていれば、この因子によって、同時にまとめて発現をオン・オフにできようになる。
ヒトでは、コルチゾール受容体タンパクにより、この調節が行われている。この転写調節因子が、まずコルチゾールと複合体を形成しないと調節部位に結合できない。
肝細胞は、コルチゾールに応答して様々な遺伝子の発現を上昇させる。
遺伝子は、すべてコルチゾール受容体複合体が、個々の遺伝子内の調節配列に結合するため、制御されるコルチゾール濃度が再び低下すると、すべての遺伝子の発現は正常レベルに戻る。
1個の転写調節因子で、多数の遺伝子の発現を調節して協調させている。 限られた数の転写調節因子を使って多数の遺伝子のスイッチを切り替え、遺伝子を発現する仕組みは、肝細胞などの機能の調節に働くだけでなく、真核生物の細胞が胚発生(多細胞生物が、受精卵から成体になるまで)を経て特定の細胞に分化する過程でも重要な役割を果たしている。発生過程では、少数の転写調節因子の組み合わせで、数多くの細胞が誕生している。 実験の段階では、培養した肝細胞に3種類の神経特異的な転写調節因子を人為的に組み込みニューロンへと変換させている。
どのような転写調節因子を組み合わせるかの研究が積み重ねられれば、やがて、各種細胞が作られるようになるだろう。
ヒトでも、コルチゾール受容体タンパクに、このような調節が見られる。これも転写調節因子だが、まずコルチゾール受容体複合体を形成しなければ、調節部位に結合できない。
多細胞生物の専門化した細胞は、細胞外のシグナルに応じて、遺伝子の発現パターンを変える。例えば、肝細胞が、ステロイドホルモン(コレステロールの誘導体の一種)の1つコルチゾールに出合うと、いくつかのタンパク質の生産量が著しく増え、様々な遺伝子の発現を上昇させる。コルチゾールは、飢餓や激しい運動や長期のストレスの際に副腎で作られ分泌されるストレスホルモンの1つで、アミノ酸などの低分子からグルコースを作るよう肝細胞に合図すると、コルチゾールが副腎から分泌される。また、アミノ酸などの低分子からグルコースを作るよう肝細胞に合図すると、例えば、アミノ酸の一種のチロシン(細胞でのタンパク質を生合成する20のアミノ酸のうちの1つ)を、グルコースに変換するチロシンアミノ基転移酵素などの酵素が増加する。肝細胞は、コルチゾールに応答して、様々な遺伝子を発現する。これらの遺伝子すべても、コルチゾール受容体複合体が、個々の遺伝子の調節配列に結合することにより制御されている。
コルチゾールが無くなると、これらのタンパク質の生産は正常に戻る。
受容体タンパクはコルチゾールが結合すると活性化して、DNAの特定の調節配列に結合できるようになり、特定の標的遺伝子の転写を活性化する。細胞の種類が違えば、抑制因子としても働く。
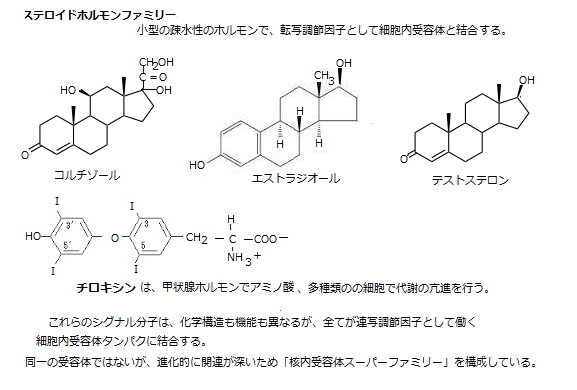
コルチゾール・エストラジオール・テストステロンなどいくつかのステロイド系ホルモン(シグナル分子)は、コレストロールの誘導体で、そのホルモン系の受容体は、細胞質に存在するが、アミノ酸の一種チロキンの誘導体であるチロキシン(甲状腺ホルモン)の受容体は、ホルモンがやってくる前から核内のDNAに結合した状態で待機している。
コルチゾールの生産部位は副腎で、殆どの組織でタンパク質・炭水化物・脂質の代謝に影響する。エストラジオールの生産部位は卵巣で、女性の二次性徴の誘導と維持、テストステロンの生産部位は精巣にあり、男性の二次性徴の誘導と維持に働く。
チロキシンの生産部位は甲状腺にあり、多種類の細胞での代謝とその亢進を担う。
ステロイドホルモンは一般に生殖腺や副腎においてコレステロールから合成される。それらのホルモン分子の構造は脂質であり、そのため細胞膜に達すると、容易に内部に通過し細胞核へ到達し、核内にある受容体タンパクと結合できる。
コルチゾールは、ストレスに応答して副腎で作られるホルモンの一つであるから、細胞膜を透過して細胞質にある細胞内受容体タンパクに結合する。この受容体―ホルモン複合体は、核膜孔を通って核内に運び込まれる。受容体タンパクは、コルチゾールと結合すると、構造を変化させ活性化し、DNAの特定の調節配列に結合できるようになる。これにより核内の転写調節因子(核内受容体)として働き、特定の標的遺伝子の転写を活性化し、あるいは阻害する。活性化すれば標的遺伝子の転写を開始する。
コルチゾールなどいくつかのステロイドホルモンの受容体は、細胞質に含まれているが、他のステロイドホルモンや甲状腺ホルモンの受容体は、シグナル分子であるホルモンが、細胞膜や核膜孔を透過するまえから、核内のDNAに結合した状態にある。
シグナル分子としてのホルモンの種類ごとに、核内受容体が異なり、その受容体ごとにDNA上のどの組み合わせの調節部位に働くかも異なる。同一ホルモンであっても、細胞の種類が異なれば、異なる組み合わせの遺伝子群を調節するので、標的細胞が異なれば生起する生理反応も異なる。
核内受容体とそれを活性化するホルモンは、ヒトの生理反応に重要な役割があり、テストステロン(男性ホルモン)の受容体を欠失していれば、それに関わるシグナル伝達系を失い重大な結果が生起する。ヒトのテストステロンは、胎生期には外性器を作るが、脳の発達にも影響する。テストステロンは、思春期には、男性の二次性徴の発現を促す。ごく稀に男性に、遺伝子的変異によりテストステロン受容体を欠く個体が現れる。これによりテストストロンができても細胞が応答できず、性が逆転して女性として成長することになる。これは、男性ホルモンや女性ホルモンが作れない場合に生じる、性や脳の発達経過である。テストステロンの受容体が、性的発達に決定的な役割を果たしている証明である。その受容体は、1種類の細胞に1通りのみに働くのではなく、男女を区別するあらゆる形・形態・器官などを生み出す過程で、多様な種類の多数の細胞が必要であることをもの語っている。
スポーツなどで、その投与がドーピング問題として取り上げられる「ステロイド」は、ステロイドホルモンと同様、あるいはそれより強力なホルモン作用を持つ人工的に合成されたステロイドである。
遺伝子はすべて、コルチゾール受容体複合体が、個々の遺伝子内の調節配列に結合することによって制御される。コルチゾール濃度が低下すれば、すべての遺伝子の発現は安静状態に戻る。この一個のコルチゾールという転写調節因子で、多数の遺伝子の発現が協調させられている。
細胞の種類が違うと、コルチゾールに対する反応も違う。肝細胞はステロイドホルモンの1つコルチゾールに出合うと、アミノ酸からグルコースを作るよう合図されるので、チロシンをグルコースに変換するチロシンアミノ基転移酵素が増加する。逆に、脂肪細胞ではチロシンアミノ基転移酵素の生産を減少させる。またコルチゾールに全く反応しない細胞もある。細胞の種類が異なれば、同じ細胞外シグナルに対する応答の仕方も違うことも多く、これが細胞の専門化の一因となっている。それにより、細胞それぞれが独特の特徴を作り出しいく。
限られた数の転写調節因子を使って多数の遺伝子のスイッチを切り替える仕組みは、通常の細胞機能の調節に役立つばかりか、真核生物の細胞が胚発生を経て、特定の細胞へ分化いていく過程でも重要な役割を担っている。その顕著の例が、筋細胞の発生である。
哺乳動物の典型的な骨格筋細胞は他の細胞と大きく異なり、細胞質ゾル(サイトゾル;cytosol;細胞質基質)にとどまる筋収縮装置を構成する2種類の収縮タンパク質であるアクチンとミオシンや、受容体タンパク、筋細胞を神経刺激に対して感受性にする細胞膜のイオンチャンネルタンパク(膜貫通タンパク質の一種で、受動的にイオンを透過させるタンパク質の総称)など、特徴があるタンパク質を数多く持っている。
目次へ