| Top 車山高原 車山高原お知らせ 車山ブログ
|
 前方の車山の南山麓に湧き出る、大規模な伏流水・「大清水湧水」を活用して、米沢では、稲作や新鮮な野菜・花卉の栽培が盛んです。晴天率が非常に高く、日照時間が長いという環境の恵みもあります。 前方の車山の南山麓に湧き出る、大規模な伏流水・「大清水湧水」を活用して、米沢では、稲作や新鮮な野菜・花卉の栽培が盛んです。晴天率が非常に高く、日照時間が長いという環境の恵みもあります。光を十分に吸収した葉緑素が、光合成を行うことにより、デンプンを作ります。 米沢のように、晴天が続き、夜涼しい気候で、稲が育つと、日中に作られた養分は安定的にうまみ成分となって実に蓄積され、美味しい「お米」になります。 昼と夜の「寒暖差」が、特に重要なのです。昼間暑い間に養分を作り、夜寒くなると、その養分を実に回します。 寒過ぎると植物は枯れてしまいます。その危機を察知し、子孫を絶やさないために、植物は種子を充実させようとします。それで、昼暖かく、夜寒い地方の「お米」が美味しいのです。 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体
| 目次 |
| 1)イネにおける出穂期 |
| 2)クロマチンの構造と機能 |
| 3)レトロトランスポゾンとDNAトランスポゾン |
| 4)エピジェネティクスな修飾 |
| 5)ユビキチン化 |
1)イネにおける出穂期
イネにおいて出穂期は、品種の地域適応性に直接影響する重要な品質形成期である。このため、多くの研究者らが、出穂期を支配する遺伝機構の解明というテーマで、様々な研究を行ってきた。その成果は歴然で、出穂期の遺伝機構に関する重要な知見が数多く蓄積されてきた。いずれも既存品種を主要な分析対象としており、従来の遺伝子分析法、即ち形質の異なる品種・系統間で交雑を行い、その後代F2およびF3において分離分析を行い、分離に関与する遺伝子座locus( ˈləʊkəs;各遺伝子が染色体に占める位置)を検出・同定するという方法が用いられている。
遺伝子座間相互作用を解析するために、 1品種より人為的に誘発した出穂期突然変異系統を、既存品種とともに分析材料に用いている。イネは、 1年性の短日植物short-day plantsで、短日条件下でのみ花成する植物であるため、限界暗期critical dark periodにより、継続した暗期を含む光周期が与えられると、花芽分化を開始・促進する。
光周期による花芽形成の制御においては、明期の長さではなく、中断されない暗期の長さの方が重要である。植物に一定時間より長い中断されない暗期を与えたときに、花芽形成が促進される。その継続暗期を限界暗期という。
長日植物と短日植物いずれも、光周期刺激は葉により、そのシグナルを受容する。例えば、オナモミ属では、植物体の他の部分が長日条件に晒されていても、たった一枚の葉を短日処理するだけで花を形成する。日長に応答した葉は、シュート頂へ花成の移行に調節するようシグナルを送る。
キク科オナモミ属の植物は世界に広く分布する一年草である。日本でも各地の荒地で普通に見かける。茎はよく分枝し、下向きの短い刺が生え、この棘を動物の体に引っ掛けて種子を伝播させる。葉を揉んでつけると虫刺されに効くところから「ナモミ(生揉み)」の名前が付いたとも言われている。
日長により制御され、それにより葉で始まるシュート頂への花成形成までのシグナル伝達過程全体を、光周性誘導photoperiodic inductionと言う。光周性誘導は、ドイツを代表する文豪ゲーテが、自然科学研究者として著した「植物変態論」に、「葉という基本的な器官が変形して花となった」と書いている。
光周性誘導は、植物体から切り離された葉でも起こる。短日植物のシソは、切り離されて短日条件に晒された葉は、長日条件に置かれ、短日の誘導を受けていない植物に接ぎ木すると、その植物に花成を引き起こす。この実験により、光周性誘導が葉だけに起こる事象であることの証明となった。
また、自然条件では、昼と夜の長さを合わせて明暗の24時間サイクルになる。短日植物は、日照時間が短くなると花が咲く植物と言われたが、多くの実験的研究により、イネ・キク・アサガオ・コスモスなどは、必ずしも暗期のほうが明期よりも時間的に長いことは要せず、種ごとに決った一定時間以上の暗期の長さが、光周期を与えて花芽を形成する。例えば、短日植物は、限界値よりも長い明期に晒されても、その後に夜が十分に長ければ、花成を誘導できている。一方、短い昼と短い夜の組み合わせでは、短日植物は花成しない。短日植物の光周性計時は、中断されない暗期の持続時間を計るものであった。
光周性においては、光は光合成のエネルギー源としてではなく、シグナルとして利用されていた。光情報はフィトクロムやクリプトクロムといった光受容体を介して、概日時計に入力され、概日時計を用いた計時機構により日長が測定されいる。オナモミでは、8.5時間を超える暗期、大豆では10時間を超えなければ花芽を形成できない。アサガオの限界暗期は、9時間、シロガラシの限界暗期は12時間、通常、生育地の緯度が高くなるほど限界暗期は短くなる。
イネのアケボノ品種の限界暗期は約10時間である。この限界暗期では出穂までに56日かかるが、30分延長して10時間半の暗期にすると、出穂までの日数は34日、1時間延長して11時間にすると30日に短縮できる。アケボノ品種の主産県は岡山で、南部の肥沃地に適し、倒伏しにくく、かつ、安定多収であると言う。
なお、長日植物は、短日条件でも、それに続く夜の時間が短ければ、花成することが明らかになった。そのため、長い昼と長い夜という条件は、花成の誘導には繋がらない。
目次へ
2)クロマチンの構造と機能
最近の花成研究によれば、花芽形成には多くのシグナル伝達経路が関与し、シロイヌナズナの花成には、長日条件促進経路 (光周期依存促進経路)・春化処理抑制経路・ジベレリン信号促進経路や、成長プログラム制御促進経路が分子遺伝学的に同定された。 それらの遺伝子と併せて、flowering locust (FT)は、花成信号統御遺伝子と呼ばれる。それらの花成信号統御遺伝子が同定されたことで、茎頂分裂組織の形態変化に先立って、花芽誘導の状態を知ることができるようになった。シロイヌナ ズナFT遺 伝子とイネ遺伝子群の花成信号統御遺伝子の一つであるFT遺 伝子は、比較的小さなタンパク質をコードしている。
FT遺伝子は、地上部での発現が開花時期に先んじて上昇することから、葉で作られる花成シグナルの生成に関与していると考えられる。しかし、葉からの花成シグナルを受けて、葉で生成される花成シグナルに関与しる遺伝子に留まることなく、花芽誘導に伴い発現すると見られている。
興味深いことに、イネゲノム中に15を越えるFTと相同性homology(形態や遺伝子が共通の祖先に由来する)を持つ遺伝子があり、その内の約10個がFTの遺伝子重複である。遺伝子重複とは、遺伝子を含むDNAのある領域が重複する現象のことである。遺伝子重複が起こる原因としては、遺伝的組換えの異常、レトロトランスポゾンの転移、染色体全体の重複などがある。
 |
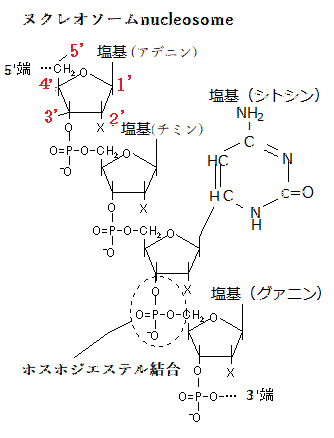
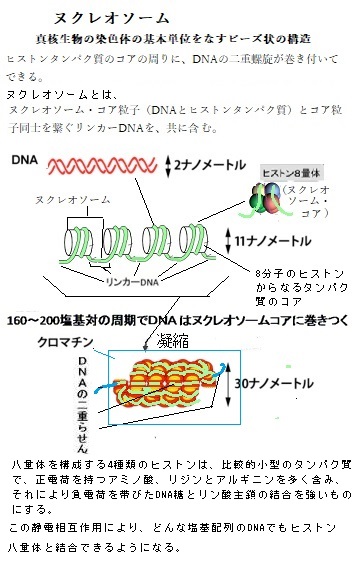 核ゲノムは、クロマチンchromatinのかたちに折り畳まれている。クロマチンとは、8つのヒストンタンパク質分子(ヒストンは、アルギニンやリシンなど塩基性アミノ酸に富む塩基性タンパク質)からなる筒状構造に、それに巻き付く核ゲノムの二重らせん構造のDNAとその他のタンパク質が形成するヌクレオソームnucleosomeが、染色体DNAの全長にわたり糸を通したビーズが並ぶような構造となっている。それが染色体の基本構成単位となる。クロマチンとは、そのビーズ状の構造を、更にコンパクトに凝縮収納した構造体である。ゲノムDNAは、そのクロマチン構造を形成して核内に収納されている。ヒトの一個の細胞に存在する染色体DNAをまっすぐ伸ばすと、その長さは約2mにも達する。それに対して、細胞の直径は約10
μm (1 mmの100分の一)であり、その中でクロマチンは直径約30 nm(nanometre ;1 µmの1,000分の一;10−9m)で収まる。
核ゲノムは、クロマチンchromatinのかたちに折り畳まれている。クロマチンとは、8つのヒストンタンパク質分子(ヒストンは、アルギニンやリシンなど塩基性アミノ酸に富む塩基性タンパク質)からなる筒状構造に、それに巻き付く核ゲノムの二重らせん構造のDNAとその他のタンパク質が形成するヌクレオソームnucleosomeが、染色体DNAの全長にわたり糸を通したビーズが並ぶような構造となっている。それが染色体の基本構成単位となる。クロマチンとは、そのビーズ状の構造を、更にコンパクトに凝縮収納した構造体である。ゲノムDNAは、そのクロマチン構造を形成して核内に収納されている。ヒトの一個の細胞に存在する染色体DNAをまっすぐ伸ばすと、その長さは約2mにも達する。それに対して、細胞の直径は約10
μm (1 mmの100分の一)であり、その中でクロマチンは直径約30 nm(nanometre ;1 µmの1,000分の一;10−9m)で収まる。DNA の高メチル化methylation(ˌmɛθəˈleɪʃən)や抑制型ヒストン修飾の増加によりクロマチン構造が凝集すると、遺伝子発現の活性が抑制される。
2種類のクロマチンがあり、ユークロマチンeuchromatin(真正染色質)とヘテロクロマチンheterochromatin(異質染色質)と呼ばれる。ヘテロ (hetero) とは、「異なる」という意味を持つギリシャ語由来の接頭語である。ヘテロクロマチンは、通常、ユークロマチンよりは堅く折り畳められいるため、凝集度が高く、より密度が高い。ユークロマチンの方は、凝集度が低いためか、遺伝子発現が活発で、遺伝子領域の割合が高い。その凝集の少ないユークロマチンで、遺伝子の多くは活発に転写されている。その一方ヘテロクロマチン領域にある遺伝子は、殆どの組織では不活性か遺伝子発現が抑制されている。その転写活性のないヘテロクロマチンは、DNAの約10%を占めている。
殆どのヘテロクロマチンは、核膜周辺に濃縮され、テロメアやセントロメアなど少数の遺伝子しか含まない染色体領域と関係している。残りのDNAは、凝縮せず分散し、遺伝子の発現が活発なユークロマチンを形成している。とは言っても、いつでも転写が活発なユークロマチンは僅か10%程度で、残りはユークロマチンとヘテロクロマチンとの中間体として凝縮している。それぞれの染色体は、核質内の決まった場所に独立して局在し、そのため染色体ごとに異なる制約を受ける。
そのヘテロクロマチンの反復配列で主要な一つにトランスポゾンtransposonがある。あるいは、転移因子 (transposable element) とも呼ばれ、いずれもジャンプする遺伝子と知られている。転移因子は、ゲノム上を「動く(=転移)」ことのできるDNA因子である。これは、いくつかのトランスポゾンが、ゲノムの中の新たな場所に、自分の配列のコピーを挿入する能力があることを示す。 トランスポゾンには、複製して新たな場所へ挿入される様式の違いにより、DNAトランスポゾンとレトロトランスポゾンretrotransposonの二種類に分類されている。
目次へ
3)レトロトランスポゾンとDNAトランスポゾン
レトロトランスポゾンの「転移」は、すなわち「増幅」である。ヒトゲノムに存在する転移因子の大部分はレトロトランスポゾンである。
レトロトランスポゾンは、自分のコピーであるRNAを作り、次にそれがDNAへ逆転写され、ゲノムのどこか他の場所に再挿入される。通常は、元の場所から抜けるのではなく、自分のコピーを余分に作るため、活性のあるレトロトランスポゾンは、ゲノムの中で増殖する傾向がある。レトロトランスポゾンに由来するゲノムの量は、種間で大きく異なり、トウヒでは、レトロトランスポゾンがゲノム配列の58%を占めると言う。食虫植物のタヌキモ(日本に生息する種は、湿地に生息するミミカキグサ類と、水生のタヌキモ類)では、せいぜいゲノムの2.5%を占めているに過ぎない。
また、トウモロコシのゲノム配列の50%以上を占めるなど、植物種によっては重要なゲノム構成要素となり、植物種間のゲノムサイズの違いを生じる主要因となっている。
培養によって活性化されるイネのレトロトランスポゾンは、培養中に多様な遺伝子にランダムに転移し、高頻度で突然変異を誘発することが明らかになった。この性質を利用しイネの遺伝子単離が可能となった。トランスポゾンは植物における遺伝子単離の有効な手段として注目されている。これまでトウモロコシやキンギョソウなど、ごく限られた植物でしかその存在が知られていなかったが、最近、イネから培養によって活性化されるレトロトランスポゾンTos17が単離された。そのレトロトランスポゾンTos17を用いて、小粒遺伝子sg1のmRNA などを鋳型として逆転写酵素を用いて合成して、相補的な塩基配列を持つ 1 本鎖のcDNA(complementary DNA;相補的 DNA)を単離して増やす。この遺伝子は粒形を制御し、また光や植物ホルモンの影響下で根の形態形成にも関与している。
DNAトランスポゾンDNAtransposonは、カット・アンド・ペーストによってある場所から別の場所に動き、この反応はトランスポゾンがコードする酵素が行う。この酵素はトランスポゼースtransposaseと呼ぶ。切り出されたトランスポゾンは、ゲノムの他の場所へ挿入される。このため、殆どの場合、トランスポゾンのコピー数は変化しない。
遺伝子内への転移は、突然変異を引き起こす場合がある。トランスポゾンがコード領域に挿入されると、その遺伝子は不活性化され、遺伝子の近くに挿入されただけでも、その遺伝子の発現に変化が生じかねない。トランスポゾンが遺伝子発現の調節に関わる領域を破壊して転写を阻害したり、あるいは、トランスポゾンは、しばしばプロモーターを持つので、転写を増加させたりするかもしれない。
トランスポゾンが突然変異を引き起こすことは、宿主ゲノムの進化に重要な役割を果たすかもしれない。低い頻度での突然変異誘発によって、個体は次世代に伝わる新たな多様性を持つことになる。しかし、転移の頻度が高く、個体に多くの突然変異を引き起こすと、そのうちのいくつかは有害で、その個体の適応度を下げることになるだろう。
トランスポゾンは、挿入だけでなく、欠失・逆位・染色体切断などのゲノム再編成をひき起こす。これらの再編成は、近傍にある遺伝子の発現に影響を与えるばかりではなく、特にDNA型トランスポゾンは挿入と脱離により色素生合成に関わる遺伝子などの発現を変化させ、絞りや斑入り模様形成の原因となる。近年では、トランスポゾンをランダムに転移させた植物体の大規模なプールを作製し、得られた遺伝子破壊株を利用して遺伝子単離を行う手法が確立し、植物の研究に盛んに利用されている。
目次へ
4)エピジェネティクスな修飾
 植物やその他の生物では、トランスポゾンの活性をDNAやヒストンのメチル化で制御できるようである。これと同じ過程で、ゲノム中のヘテロクロマチン領域の転写を抑えるにも使われている。ゲノム全DNA配列が明らかにされるにしたがい、ヘテロクロマチン領域には、高度にメチル化されたトランスポゾンが多数あることがわかってきた。トランスポゾンのメチル化がヘテロクロマチン形成の原因になったのであろうか?あるいは、トランスポゾンが偶然ヘテロクロマチン領域に挿入されるためにメチル化されたのであろうか?
植物やその他の生物では、トランスポゾンの活性をDNAやヒストンのメチル化で制御できるようである。これと同じ過程で、ゲノム中のヘテロクロマチン領域の転写を抑えるにも使われている。ゲノム全DNA配列が明らかにされるにしたがい、ヘテロクロマチン領域には、高度にメチル化されたトランスポゾンが多数あることがわかってきた。トランスポゾンのメチル化がヘテロクロマチン形成の原因になったのであろうか?あるいは、トランスポゾンが偶然ヘテロクロマチン領域に挿入されるためにメチル化されたのであろうか?トランスポゾンのDNAメチル化を維持できない変異体では、眠っていたトランスポゾンが活性化されて突然変異の頻度が上がることがわかった。このようなトランスポゾンの活性化は、その子孫の適応度を顕著に下げるかもしれ ない。したがって、DNAのメチル化とヘテロクロマチン形成は、ゲノムの安定には重要な働きをしている。
エピジェネティックな修飾epigenetics modificationという言葉は、個体発生に関する説の1つである「エピジェネシス(後成説)」と、「ジェネティクス(遺伝学)」を起源としている。Modification( mὰdəfɪkéɪʃən)は、 (部分的)変更・修正・緩和・調節・(部分的)改造・修飾・限定の意味、「エピ」はギリシャ語で「優位」や「上に」という意味の接頭語である。
「エピジェネティクス」は「遺伝子の上にさらに修飾が加わったもの」「さらに優位になったもの」などという意味を持つ。ジェネティクスでは、DNAを構成するA(アデニン)・T(チミン)・G(グアニン)・C(シトシン)という4種類の塩基の並び方、すなわち塩基配列を遺伝情報の基本とする。一方エピジェネティクスでは、DNAの塩基配列は変えずに、あとから加わった修飾が遺伝子の活性を決める。
転写が始まるには、RNAポリメラーゼや他の結合タンパク質が、DNAに近付ける場合だけである。DNAに接近できるのは、ヒストンによるDNAの折り畳みが「ゆるめられる」必要があり、その過程は、DNAとヒストンによる化学的修飾による。その修飾は、遺伝子の塩基配列を変えずに遺伝子の働きを変えるため「エピジェネティックな修飾」と呼ばれる。DNA の低メチル化や活性化型ヒストン修飾の増加により、クロマチン構造が緩むと遺伝子発現が活性化される。
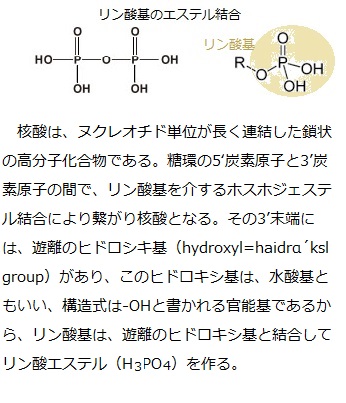
DNAの修飾は、シトシン残基のメチル化である。哺乳動物では、4つの塩基の組み合わせの中で、シトシンの次にグアニンが続くCGの配列のシトシンにメチル基(-CH3)が付加され、5メチルシトシンになることをDNAメチル化という。哺乳類では3種類のDNAメチル化酵素と3種類のDNA脱メチル化酵素が見つかっている。
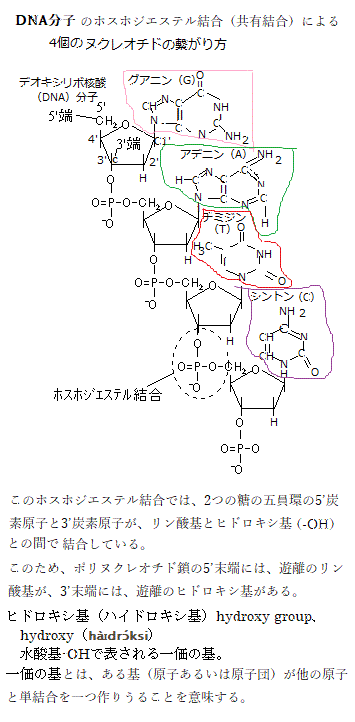
 DNA配列のなかには、CG配列が集まって密に存在する領域(CpG island)があり 、 CpGの「p」の文字は、シトシンとグアニンの間のホスホジエステル結合phosphodiester
bond(炭素原子の間がリン酸を介した2つのエステル結合によって強く共有結合している。phospho;リン酸phosphoric acid、diester;1つの分子の中に2つのエステル結合原子団を含む有機化合物)を表している。哺乳類の遺伝子のうち40%近くが、プロモーター内部もしくはその近傍にCpG islandを含んでいるとされる。CpG island
のDNAメチル化は、遺伝子が働きはじめる過程で深く関与することが知られている。 植物では、CGとCHGとCHH(H = C,T,A;G以外のいずれかの塩基)の配列でシトシンがメチル化される。複数のDNAメチル化酵素が、シントンのメチル化を触媒し、DNAの脱メチル化は、グリコシラーゼglycosylaseが行なう。グリコシラーゼは、メチル化されたシントンをメチル科されていないシントンと置き換える。
DNA配列のなかには、CG配列が集まって密に存在する領域(CpG island)があり 、 CpGの「p」の文字は、シトシンとグアニンの間のホスホジエステル結合phosphodiester
bond(炭素原子の間がリン酸を介した2つのエステル結合によって強く共有結合している。phospho;リン酸phosphoric acid、diester;1つの分子の中に2つのエステル結合原子団を含む有機化合物)を表している。哺乳類の遺伝子のうち40%近くが、プロモーター内部もしくはその近傍にCpG islandを含んでいるとされる。CpG island
のDNAメチル化は、遺伝子が働きはじめる過程で深く関与することが知られている。 植物では、CGとCHGとCHH(H = C,T,A;G以外のいずれかの塩基)の配列でシトシンがメチル化される。複数のDNAメチル化酵素が、シントンのメチル化を触媒し、DNAの脱メチル化は、グリコシラーゼglycosylaseが行なう。グリコシラーゼは、メチル化されたシントンをメチル科されていないシントンと置き換える。遺伝子の活性に影響するエピジェネティクスな修飾は、タンパク質ファミリーであるヒストンでも起こる。ゲノムDNAはヒストンというタンパク質に巻き付いた状態で核内に収納され、ともにヌクレオソームを構成する。この構造をクロマチンという。それぞれのヒストンには、ヒストンのコア領域に含まれないN末端・C末端側の領域に、ヒストンテイルhistonetailを持ち、それがヒストンのアミノ酸鎖の最初の部分を構成し、ヌクレオソームの外に出ている。ヒストンの修飾は、これらのテイルで起こる。これらの修飾は、ヌクレオソームの立体構造に影響し、それによってその領域の遺伝子の活性に影響する。
アセチル化、メチル化、リン酸化、モノユビキチン化など様々な翻訳後修飾を受けている、これらの修飾はクロマチン構造を変化させ、エピジェネティックな遺伝子発現制御に関わっていると考えられている。
目次へ
5)ユビキチン化
 ヒストンがメチル化・アセチル化・リン酸化やユビキチン化などの修飾を受けると、クロマチン構造が変化し、DNAと転写因子などの核内因子とが接近のしやすくなり、遺伝子の発現制御が可能となる。
ヒストンがメチル化・アセチル化・リン酸化やユビキチン化などの修飾を受けると、クロマチン構造が変化し、DNAと転写因子などの核内因子とが接近のしやすくなり、遺伝子の発現制御が可能となる。ユビキチンubiquitinは、ATP加水分解で放出されるエネルギーを用いて、ユビキチンリガーゼという3つの酵素により、タンパク質に共有結合する低分子のポリペプチドである。ユビキチン化は、巨大なタンパク質分解複合体であるプロテアソームproteasomeに認識される部位として働くタンパク質修飾の一種で、ユビキチン化されたタンパク質はよく分解される。
タンパク質の分解を行う巨大な酵素複合体であるプロテアソームは、真核生物の細胞質及び核内のいずにも分布している。今では、クライオ電子顕微鏡による構造解析の向上・進展により、プロテアソームはユビキチン化タンパク質を確実に分解するために隅々までうまくデザインされた分子マシーンmolecular machineであることが分かってきた。その結果、細胞内タンパク質の恒常性を維持するが、細胞周期の進行やシグナル伝達など様々な生命現象において必須の役割を果たしている。
近年、クライオ電子顕微鏡cryo-electron microscopyによる構造解析が驚異的に進展し、プロテアソームはユビキチン化タンパク質を確実に分解するための分子マシーンであることが分かってきた。
スウェーデン王立科学アカデミーは、2,017年のノーベル化学賞を、スイスのジャック・デュボシェ氏、英国のリチャード・ヘンダーソン氏、米国のヨアヒム・フランク氏に贈ると発表した。授賞理由は「溶液中の生体分子の構造を高い解像度で観察できるクライオ電子顕微鏡(低温電子顕微鏡)の発明」であった。
サンプルを含む水を急速に冷却してガラス化させる方法を開発し、合わせて電子顕微鏡の改良が進み、真空中でも元の構造を維持したままで生体分子を観察することを可能にした。クライオ電子顕微鏡で観察する手法で得られる二次元デジタル画像から、生体分子複合体の三次元立体構造に、計算機上で再構築することにより、生体分子複合体構造の解析が明解なった。そのため、生体分子の3次元構造が容易に生成できるようになった。
分子マシーンは、並進や回転などの制限された動きを誘発する分子や分子集合体を意味する。生体内には、様々な働きをする分子マシーンがあり、その多くはATPがADPとPi(リン酸)に分解する時に出す高エネルギー物質との結合を引き金に駆動する。
分子モーターとも呼ばれる、生体内のタンパク質が、方向性のある動きによってなんらかの機能を発現する。タンパク質はアミノ酸が連なった「ひも」状のポリマーpolymer(重合体)で、タンパク質が機能するためには、アミノ酸配列が、最も安定した状態になる「立体構造」に折れたたまれる必要がある。その折れたたみをフォールディングfoldingと呼ぶ。分子シャペロンmolecalar shaperonというタンパク質ファミリーは、リボソームから産まれたばかりのタンパク質がきちんとフォールディングするのを助けている。
タンパク質をはじめとする生体高分子で混み合っている細胞内において、シャペロンはフォールディング途上の不安定な中間体や熱で変性したタンパク質が、周りに同様の不安定な「ひも状」にあるタンパク質と、相互に絡まり合う(凝集)状態にならないように、フォールディング化を助けている。代表的なシャペロンには、シャペロニンchaperoninと呼ばれるファミリーがあり、ATPを使って多くのタンパク質のフォールディングを助けている。分子シャペロニンは、ATPの結合と加水分解に応答してその筒状構造を開いたり、閉じたりして、変性タンパク質の取込み、およびリフォールディングが完了したタンパク質の吐出を行う。
タンパク質のフォールディングの異常が、アルツハイマー病・パーキンソン病・ハンチントン病(常染色体優性遺伝によって発病する神経変性疾患、舞踏運動といわれる異常運動と、認知症や人格変化が特徴)・狂牛病(プリオン)など哺乳類の神経変性疾患に繋がり、そこにシャペロンが深く関与していることが知られてきている。
近年ユビキチン修飾が、タンパク質分解以外にも、DNA修復やシグナル伝達など、様々な生命現象に関わることが明らかとなった。ヒストンが、ユビキチン化などにより翻訳後修飾を受け、その翻訳後修飾が、遺伝子転写や遺伝子修復など、様々な現象に関与していることが明らかにされた。ヒストンを構成されるコアヒストンの一つH2Aは、1,975 年に最初のユビキチン化タンパク質として同定され、それ以後の研究により遺伝子転写制御に関与していることが分かった。
遺伝子発現の活性化に関与するメチル化は、特にヒストンH3テイルと言う特定のリジン残基lysine residue(必須アミノ酸、略号は Lys、一文字で省略するとK。Lysine;láisiːn、residue;rézəd(j)ùː;残余・残留物)のメチル化が影響する。そのヒストンの最も外側に出た部分でも、メチル化される。1つのリジンに、1~3つのメチルが付くことができる。メチル基は、ヒストン脱メチル化酵素で除かれる。
ヒストンテイルのもう一つの修飾形式がアセチル化acetylationで、これはヒストンアセチル化酵素が触媒する。通常、アセチル化ヒストンは、頻繁に転写されている遺伝子に見られる。ヒストン脱アセチル化酵素が、アセチル基を除去し、この活性化を元に戻す。
メチル化もアセチル化も、クロマチン複合体の構造を変えて、クロマチンを凝集したり緩めたりする。そのいずれの変化も、複数のタンパク質からなるクロマチン再構成複合体が、修飾されたヒストンに結合した時に起こる。これらの複合体は、クロマチン再構成複合体の中で、ヌクレオソームnucleosomeを5‘から3’へ動かすことでクロマチンを開く。そのエネルギーは、ATP加水分解で放出される。こうしてできるヌクレオソーム間の隙間が広いので、RNAポリメラーゼがそこに結合して転写を開始させる。あるいは、ヒストン修飾で、新たにできに結合部位に制御分子が結合して、転写活性を有効に働かせる。
ヒストンテイルにある40残基ほどあるアミノ酸配列のそれぞれの位置における化学修飾は、漸く解明されたばかりである。メチル化とアセチル化の他にも、リン酸化やユビキチン化などによる他の型のヒストン修飾も、特定の遺伝子の転写活性に影響し、それら特定のヌクレオソームにおけるヒストン修飾の全体像を、ヌクレオソーム構成因子と遺伝子活性の関連を強調して「ヒストンコード」と呼ぶこともある。「ヒストンコード」とは、ヒストンに書き込まれた遺伝子発現制御をコードする「ヒストンの遺伝情報」の意味である。
エピジェネティクスの主要制御機構は、DNAメチル化とヒストン修飾であるが、個体発生や細胞分化の過程をはじめとして、様々な生命現象における重要なのメカニズムである。これらのエピジェネティックな修飾に起因する多くの疾病が解明されてきている。
その中の一つFTL遺伝子は、過剰に発現すると、植物が再生後直ぐに花芽形成を起こし、イネの穂を単一の小花から花序に変えてしまほどの強い花芽形成促進能を持ち、また、花序形成自体にも深く関与している。
多くFTのオーソログ(他種間で同一機能と構造を持つ遺伝子で、同じご先祖さんから出たと推定され、種分岐の際には、共有された遺伝子だったもの)が、イネにあるか定かではないが、広い地域での自然環境下でも、日長条件など繁殖能に関わる、それぞれ異なる発現制御が働いているようだ。しかし、どんな環境下でも、FTオーソログのどれかが発現すれば、花芽形成が始まると見られている。
また、多くのオーソログをゲノムに持つことで、種内の多様な遺伝的バックグラウンドとなり、それが適応能力を向上させているようだ。
イネの生育ステージのうち、花芽形成開始~出穂(生殖生長期間) ~登熟(登熟期間)までの期間の長さは品種・系統間で概ね同じである。違いが生じるのは、播種~花芽形成開始の期間(栄養生長期間)の長さである。品種・系統の出穂期の違いは、栄養生長期間の違いによる。
1品種より人為的に誘発された突然変異系統は、 遺伝的背景が原品種に近く、出穂期に関する遺伝子分析には好適な材料となる。しかも、出穂期は、突然変異によって比較的容易に突然変異系統を得ることができる。突然変異系統の研究により、遺伝的背景の複雑さにとらわれず、分析することが可能となったと言う。遺伝子座でイネの品種の出穂期に関する変異が明らかになり、出穂期を支配する主働遺伝子座の数がそれほど多くないことが分かってきた。
目次へ