| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物
| 目次 |
| 1)種子植物の分類 |
| 2)花葉floral leafと苞葉bract leaf |
| 3)進化する種子植物 |
| 4)植物種の分類 |
| 5)植物種と倍数性の関係 |
| 6)シダ植物の生殖 |
 種子植物は、植物界の一門で、花が咲き、種子を生じる一群をいう。裸子植物と被子植物とに分けられ、種子植物が獲得した最も顕著な形態的特徴が花であったため、かつては顕花植物と言われた。
種子植物は、植物界の一門で、花が咲き、種子を生じる一群をいう。裸子植物と被子植物とに分けられ、種子植物が獲得した最も顕著な形態的特徴が花であったため、かつては顕花植物と言われた。農業上、主要な食料資源であり、木材や医薬品の用途など、古来より極めて有用な植物である。
陸上の生態系でも、優占的な生物であるため、3億年ほど前に現れた種子植物が、これまで植物科学の主な研究対象になってきた。
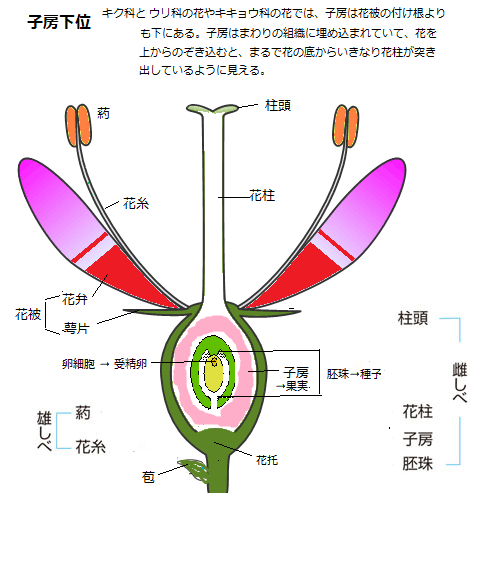
被子植物の対語である裸子植物は、子房がなく胚珠が心皮(雌ずいを形成する葉を心皮という)に包まれることなく、表面に裸出する一群の植物の総称である。
陸上植物は、進化の過程で葉(花葉floral leaf)を変形させて、受精して種子となる胚珠を包む器となる雌しべを作り、その雌しべを生殖器官の中核とした。
雌しべと雄しべを持つ植物が、被子植物であり、真の花の登場は、恐竜が地上を闊歩していた、今から1億数千万年前と推定される。
真に花と呼べるものは、被子植物しか持たない。花弁はその中の特殊な器官であるから、花弁のある植物は全て被子植物と言える。被子植物の種のおよそ7割は、花に雄しべと雌しべの両方がある。
顕花植物には、裸子植物と被子植物があり、裸子植物の生殖器官も花と見立てる見解もある。
種子は、シダ植物における大胞子嚢(胞子が、その内部に生じる袋状の生殖器官)と、それを覆う構造から生じたもので、本来は胞子形成を行う葉の裏面に付着していた。
裸子植物では、胞子葉(胞子を形成する葉)と区別がつきにくいものもあるため、被子植物の花だけを花と呼ぶ考えもある。英語で Flowering plant と言った場合、被子植物のみを指す。 裸子植物には花弁がないことは確かである。
「花は被子植物が作る生殖器官」という定義がある。その定義からすると、裸子植物には花がないことになる。それでは、裸子植物の花粉や胚珠を作る生殖器官をなんと呼ぶのかというと、これが実は専門用語しかない。そのため裸子植物でも、「花」あるいは「花にあたるもの」と書くことが多い。
目次へ
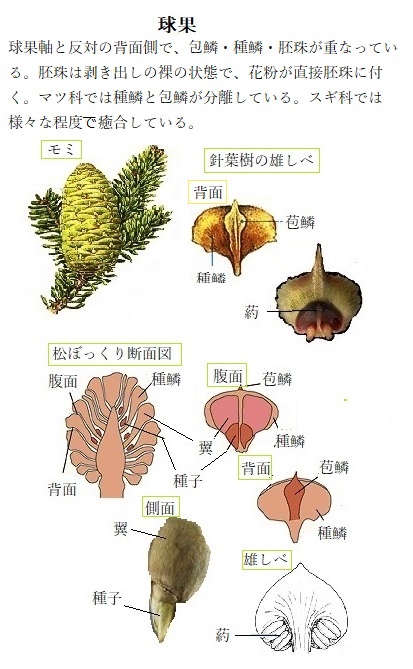 植物の後胚発生post‐embryonic development(embryonic;èmbriάnɪk胚の)段階での幼若相と成熟栄養相の根本的な違いは、成熟栄養相の段階にある裸子植物には、球果coneという、生殖のための構造を形成する組織を保有する点にある。
植物の後胚発生post‐embryonic development(embryonic;èmbriάnɪk胚の)段階での幼若相と成熟栄養相の根本的な違いは、成熟栄養相の段階にある裸子植物には、球果coneという、生殖のための構造を形成する組織を保有する点にある。裸子植物のマツ・スギ・モミ・トウヒ・ヒノキなどの果実が球果である。これらの植物は雌雄異花で、雌花は球花(きゅうか)と呼ばれる、1本の軸から多岐に伸びる雌しべの基部に2つの胚珠をつけたものが球状になった、いわば球果をつける。
受精後、雌しべは発達し、軸上に多数の鱗片葉を作る。球果の実態は、胞子をつけるように分化した胞子葉の集まりで、松かさに代表されるように楕円体の球果となる。その鱗片葉の付け根には、通常、翼を持った種子が2個ずつ付着している。鱗片葉は、種子をつける種鱗seed scaleと、それを支える苞鱗 bract scaleで形成される。植物の種により様々な形をしている。種鱗は花期に胚珠をつける一種の大胞子葉となる。
総苞は、包(包葉)、または単純化した茎葉が花序の基部に輪状に取り囲むように集まったものを言う。1枚ずつの包または葉を総包片もという。一つの総包は一つの花序に対応する。
苞葉bract leafともいい、集合体としては苞bractと呼ばれる。花を抱く葉でも普通葉と変わらない場合には苞葉とはよばない。 球果類において、胚珠をつけた種鱗 seed scale は、苞鱗 bract scale の腋についているので、苞鱗も特殊化した苞葉と見ることができる。
球果類の球花 (球果) を構成する苞鱗と種鱗が、様々な程度で癒合している。苞鱗は被子植物における苞葉 (小苞) にあたり、種鱗は胚珠をつけるので花葉 (心皮) に相当する。
胞子葉は、通常の葉と異なり、胞子を形成する葉のことで、シダ植物では、成熟した葉が胞子葉となる。裸子植物の雌の生殖器官であり、被子植物では雌しべにあたる。
マツ・モミ・トウヒやアメリカ合衆国西海岸の海岸山脈に自生する最も巨大な木のセコイアなどの針葉樹など、現生の裸子植物の多くの種子は、雌花が球果状(松かさ状)となる。現在ではイチョウ類(裸子植物イチョウ科)・ソテツ類(裸子植物ソテツ科)や古くから生薬の麻黄として用いられたマオウなどのグネツム類を除いた多数の種が球果植物に分類され、独立したグループとして扱われることもある。ヤシは、被子植物のうち、1枚の子葉を持つ単子葉植物である。
ただし、裸子植物の生殖器官が花だとしても、花弁は見当たらない。胚珠を抱えた器官や、花粉を抱えた器官がそれぞれ雌しべや雄しべに相当するとしても、その外側に特別な器官があれば萼片や花弁ということになるが、それも見当たらない。
目次へ
 「花弁のある植物は全て被子植物に分類される」。被子植物が作る生殖器官は「花」であり、花弁は花しか作らない。花の中のどの部分を花弁と呼ぶのかは、花という構造体があった場合、その花被を構成する一番外側に並んでいる器官を萼片と定義することになっている。
「花弁のある植物は全て被子植物に分類される」。被子植物が作る生殖器官は「花」であり、花弁は花しか作らない。花の中のどの部分を花弁と呼ぶのかは、花という構造体があった場合、その花被を構成する一番外側に並んでいる器官を萼片と定義することになっている。種子植物の花は、一つの枝の各葉が分化し、生殖器官となったもので、心皮は、雌しべを構成する葉(よう)的器官、つまり、雌しべ(雌ずい) を形成する葉を心皮という。この語は、ドイツの詩人ゲーテが『植物変態論』において述べている。
胚珠は心皮の向軸面(茎頂分裂組織に向いている側)の縁に沿ってつく場合と面全体につく場合とがある。 心皮は、雌しべを構成するために特殊な分化をした葉で、子房・花柱・柱頭に分化する。シダ植物でいえば大胞子葉にあたるものが進化的変形である。
3億年ほど前に現れた種子植物が裸子植物で、針葉樹・ソテツ類・イチョウや、非常に多様に裸子植物として進化したグネツム類を含む。現在約1,000種が知られている。
裸子植物で最大のグループが針葉樹で、商業価値の高いマツ・モミ・トウヒ・ヒノキ・セコイアなどがこれに属する。セコイアは、ヒノキ科セコイア属の常緑針葉樹、高さ100m近くにもなる世界有数の大高木である。セコイア属は、このセコイアだけの1属1種である。
被子植物の適応進化は、そのごく初期の段階から、様々な動物、特に昆虫と密接な共進関係にあった。被子植物の発生当初の約1億8,300万年前から、頻発する過酷な環境変化に翻弄され続け、それに辛うじて対応できる手段が、他家受粉による多種多様な遺伝子組み換えと、種子を出来るだけ広範囲に、しかも的確に散布することにあった。
初期の被子植物は、ゲンゴロウ・コガネムシ・ホタル・カミキリムシ・ハムシ・テントウムシなどの甲虫(こうちゅう)が送粉していたようだ。中にはハエやガによる送粉もあったらしい。初期段階では、被子植物の花は、特段、特徴的な形態にこだわらなかったようだ。
それでもごく初期の頃から存在する被子植物の訪花昆虫は、蜜や糖・タンパク質などの栄養価に誘われて来る様々な動物の進化、特に昆虫の進化と密接に関わってきた。とりわけ、被子植物の種子は、果実に包まれ、その散布形式は様々な共進化と適応化による結晶と言える。
種子植物の中で最も種数の多いのが被子植物で、現在約37万種が同定されている。分類学者によるコンピュータモデリングにより、更に1万7,000の未記載種の存在が予測されている。被子植物の中でも、特に初期に進化したと思われる痕跡を留める種が2つある。1つが現在のセンリョウに似た小さな緑色の花を持つ種類と、もう1つがモクレンの花に似た種で、花を構成する各要素が多数、螺旋状に並び、花も比較的目立つ形となっている。
化石や地質などの研究によれば、被子植物は、1億8,300万年前に現れたモクレン類が最初のようだ。ちょうど恐竜が王者として君臨していた中生代であった。1億6,900万年前のある過去の時期に、原始的双子葉植物から単子葉植物が進化したと考えられている。それは変異で生じた原始的単子葉植物が、現代にまで生き残ってこれたのは、自然が選択してきた結果で、「必然性」とは違った「環境適応」といった次元ではなかったか。
単子葉植物は、双子葉植物とともに被子植物を大きく2分する植物群で、種子が発芽する際に1枚の葉 (子葉) を出す。双子葉植物では2枚または2枚以上の子葉を持つ。葉脈の通り方・維管束の並び方・根の形などの特徴がキーになり、両者は、はっきりした生活観の違いとなって分かれている。
被子植物であれば、葉脈の通り方を見ればそれが双子葉類か単子葉類かすぐ分かる。双子葉類の葉脈は、葉身の基部から数本の葉脈の太い支脈が掌状に出ている(掌状脈)。単子葉類の葉脈が葉の長軸方向にほぼ平行に走っている(平行脈)。前者はスミレ・アブラナ・サクラ・アサガオなど、後者は、植物の科の中で最も多くの種が知られているラン科、単子葉植物の中で2番目に多くの種が記載されているイネ科とユリ科・ギボウシ属も含まれる。イネ科はまた、カヤツリグサ科と共に、風媒による受粉へ二次的に進化した。
単子葉植物は、双子葉植物に比べ、形態に共通点が多い。細く立った葉は、周囲の様々な方向からの輻射光を受けるのに有利となり、上に葉を伸ばせるため草原で優占しやすい。材の構造が双子葉植物などのそれと、かなり異なっており、維管束が散在する「不斉中心柱」をつくる。また、根についても主根がはっきりしない、ひげ根ばかりのものが多い。花びらの数は、3と6枚(3の倍数)が多い。
双子葉植物では、胚の幼根がそのまま発育した根となり、裸子植物にも見られるが、まっすぐに地中に伸び、周囲に多くの側根を出す。また維管束が環状に並ぶ、花びらの数4、5枚(4か5の倍数)が多い。双子葉類を花びらの特徴によって、さらに分けることができる。花びらが一続きになる合弁花類では、キキョウ・アサガオ・オニタビラコ・カボチャ・ジシバリ・タンポポの筒状花・ツツジなどがある。一見、タンポポや野菊の花びらに見える筒状花周辺の花弁は、舌状花でいずれも1枚がひとつの花である。通常、舌状花は雄しべを欠いた雌花で、よく観察すると、基部は筒状になり、花びらに見えた舌状花は、筒状の片側が極端に張り出している形状だと分かる。
花びらが分かれている離弁花類には、バラ・スミレ・イヌガラシ・アブラナ・エンドウ・サクラ・ナズナ・ホウセンカ・ミヤコグサなどがある。
目次へ
 菊の分類で、センボンギクとノコンギク、シロヨメナとタマバシロヨマナ、ノジギクとアシズリノジギクなど、野菊のフィールドで必ず突き当たるのが、これらの間にどちらとも言えない中間型の存在である。
菊の分類で、センボンギクとノコンギク、シロヨメナとタマバシロヨマナ、ノジギクとアシズリノジギクなど、野菊のフィールドで必ず突き当たるのが、これらの間にどちらとも言えない中間型の存在である。それらにどの程度の違いがあれば種となり、どの程度似ていれば変種なるのか?それを突き詰めていくと「種とは何か」という大命題に幾度も突き当たる。
動物の場合、種の境界は、はっきりしている。同じ繁殖集団に属していれば、同種と断定してほぼ間違いはない。動物のように繁殖集団による種の概念を機械的に当てはめると、リュウノウギクとイワインチンが同じ種になってしまう。その一方では、シロヨメナでは、3~4種に分類されなければならない。
植物の場合は、例えば、キク属の場合、多くの種の間で簡単に雑種ができる。あるものは稔性も良好で、ニシガハマギクやサンインギクなど、雑種群落が長年にわたって存続している。
シロヨメナでは、外見は殆ど違わないが、染色体数の違うグループがいくつか存在し、相互の生殖隔離が働いていると見られるものもある。シロヨメナは、いわゆる「野菊」の仲間で、林縁などの半日陰になるような場所に自生する。しかも、ほぼ山地に限られる。山の木陰で小さな白い野菊を見たら、その殆どはシロヨメナと言われている。
シオン属では、シロヨメナほど分布が広く、個体数の多い種はない。草丈50cm前後、初秋から秋の初めまで、茎頂で花柄を分けて、小さい白色のキク型の頭花をつける。舌状花は殆どが白色だが、淡紫色を帯びることもある。
植物の「種とは何か」?生物学の大命題の一つでありながら、その回答は難しい。動物の種であれば、繁殖可能な同一集団という生態でほぼ間違いはない。
植物の種のあり様はもっと多様で複雑、染色体に倍数性があり、外見では識別できない。いくつもの繁殖集団に分かれている場合もあれば、頻繁に別の種と交雑する集団もある。要するに、種というグループを形づくっている要素は、そのグループごとに様々であり一様ではない。
最近では、遺伝子レベルの研究が盛んだ。遺伝子を調べれば、外見の違いでは分からないものも、遺伝子を比較すれば、核ゲノムの配列など決定的な違いとしてあらわれる。単子葉植物であるキヌガサソウのゲノムは、知られている中では最大で、約150,000Mbp(百万塩基対単位)もある。
同じ種でも遺伝子は、個体すべてにおいて同一ではない。親子でも遺伝子は違う。どこまで違えば別種なのかは必ずしも明らかではない。
逆に言えば、同じ先祖から分化した種同士あれば、どこかに種を分かつ親子が存在したはずだ。それなのに、親子で別の種となったケースは確認されていない。おそらくは、ある集団から少しだけ違った個体が生まれ、それが何万世代を経過する過程で別の種に分化したのであろう。その渦中の最中であれば、中間型が多く識別が難しいグループが錯綜することになる。それも常態化しているのではないか?
「有性生殖による遺伝子交換」そのものが、真核生物に特有で、しかも種の保存には欠かせない。さらに、真核細胞では、有性生殖にあたる接合だけではなく、特に植物細胞には、核ゲノム以外に2つの細胞質ゲノムがあるため、より錯綜する。1つはミトコンドリアのゲノムmitochondrial genomeで、それは動物細胞にもある。もう一つが色素体のゲノムplastid genomeである。色素体plastidの内、特に光合成を行うように分化したのが葉緑体chloroplastと呼ぶ。細胞質ゲノムは、いずれも細菌細胞を別の細胞が取り込んだ生物進化の名残である。その2つの遺伝子のゲノムの交換は、相当に遠縁でも遺伝子の交換が行われている。
生物的な接合は、かなり遠縁同士でも行われているのが実態である。外形的な変化に乏しいが、遺伝的には極めて多様な植物もある。種の定義・概念すら、現在、22以上あり、研究が進むほどに増加している。
スウェーデンの博物学者リンネが創始した植物の分類学は、植物の生物学的な実態を、外形から予測ができるため、名前から認識を整理するのに役立つ。ところが、純粋に生物学的な立場に固執すれば、種を分類して名前を付けること自体、正しい認識の妨げにもなりかねない。また「種とは何か?」と言う命題に答えられないまま分類することもできない。
「種という概念は、人間が作り出したもので、自然界には、種は存在しない」と断言すれば、極めて不便な結果となる。認識学としての植物分類学は、「人間の目で識別できる」ことを重視する。それは現実的で、極めて有効だ。そもそも日常的な生活環境で植物に出合えば、まずは植物名と、それが属する「科」を思い起こす。それにより、興味をもった草花のおおよその生態が想定できるからだ。
どの植物にも、「国際植物命名規約」によれば、亜種と変種、および品種があるが、どういう基準で種類分類群を設定するのか、それを定めてはいない。それは、「種とは何か」と言う根本的な命題に未だ回答できないためである。しかし、一般的には、「亜種」は分布域が隔離されている地方的な変異に、「変種」は分布が接している近接地の変異に、「品種」は白花や班入りなど遺伝的にまで固定されていない変異を表現することが多い。
植物には、種の数だけ種のあり様があるが、それぞれの個体差に拘り過ぎれば、リンネ以来の近代的な分類学が無に帰するだけで、そこからは何も生まれない。
目次へ

生物学上、環境が違えば、環境に応じて遺伝子が発現されるため、形態に多少の差異は当然生じる。それに拘り過ぎれば、「種とは何か」と問いかければ、「種の数だけある」という答えになってしまう。
学名にしても、その植物を研究に使う学者のとらえかたを表している。極論すれば、植物学者の数だけ学名が存在すると言えよう。そのため、シロヨメナの倍数体は種とは認められないのである。しかし、可能な限り生物学的な種と、認識学上の種を一致させる課題は残ったままである。
キク科のシオン属やキク属は、染色体の倍数性が明瞭で、染色体数によりその系統を推測しやすい。倍数性とは、生物あるいはその生活環の一時期において、生存に必要な最小限の染色体を何セット持つかを示す概念である。
核の中に、生物が生きるために必要な最小限の遺伝子を持つ染色体を1セット持つ。これをゲノムという。例えば、ヒトは23対(=46本:相同の染色体2本で1対)の染色体をもち、この染色体のうち1対でもかけると、生存できないか、あるいは重大な障害を引き起こす。そのため、ヒトは23対の染色体からなるゲノムを1つもつ、ということになる。
植物は、動物と異なり、系統により異なる生活環を持つ。動物のように減数分裂により、単細胞の単相(1N)の配偶子を直接作り出すのではなく、減数分裂を経て多細胞の配偶体を形成する。配偶子(精子と卵)は、配偶体の有糸分裂で形成される。精子と卵が受精してできた2 N(複相)の接合子は、成熟した胞子体へと成長した後、減数分裂を経て再び配偶体を形成する。
種子植物の配偶体における精子と卵の形成や受精の際の動態は、その種によって様々である。被子植物が行う重複受精では、2つの精子が作られ、その1つが卵と受精し、もう1つは雌性配偶体中の2つの核と融合し、被子植物の種子の貯蔵組織である核相が3 N(染色体のセットが3つ)の胚乳を作る。被子植物では、より核相の高い胚乳もある。 このようなゲノムの数が変わることにより新しい種ができるような進化を「倍数性進化」 と呼ぶ。
被子植物では、約7割の種が、倍数性種とも言われ、生物進化、特に被子植物の進化に非常に重要な役割を果たしてきた。裸子植物は、重複授精を行わない。裸子植物の種子では、1Nの配偶体が貯蔵組織として働く。
配偶体世代を作り出す細胞を胞子と定義するならば、種子植物の種子とは、まったく別の物と言える。シダ植物では、卵子と精子の融合体が、すぐに次世代へと発達する。裸子植物や被子植物になると、受精卵は、それから分裂し胚発生を始める。ある段階に達すると胚発生を中止して種子を形成するようになる。
種子植物が、種子を作るということは、植物の遺伝的な発現である。植物体の細胞核は母方由来と父方由来の染色体を、それぞれ1対ずつ持つ(2N)。成長の基本である細胞分裂では、1対の染色体が倍加し、それらが正確に2等分されるので、娘細胞の染色体のセットの数やDNAの塩基配列は変わらないはずだ。しかし、通常の環境下でも、まれに発生の初期に倍加した染色体セットが2等分されないで4セットの染色体をもった細胞ができ、そのまま植物体になることがある。いわゆる4倍体(4N)であるが、植物種としては同一である。
4倍体植物同士も正常な減数分裂をし、通常に、受精して種子形成する。その次世代も4倍体を保ったままである。2倍体植物と4倍体植物とが交配すると、2倍体(2N)植物の精子・卵子(1N)と4倍体(4N)植物の精子・卵子(2N)とが融合するため、3倍体(3N)の有精卵となる。
3倍体(3N)の有精卵でも種子は形成され、普通の娘細胞への細胞分裂にも支障は生ぜず、3倍体の植物体が正常に作られ花も咲く。外見上、顕著な違いがないため、4Nや3Nは、植物種として独立した分類群とは認めがたい。
ところが3倍体では、減数分裂の際、卵子・精子形成が正常に行われないため、正常な配偶子が形成されない。そのため受精と胚発生が不能となり、種子が形成されない。3倍体の繁殖グループが存在しないのは、正常な減数分裂ができないためである。
目次へ
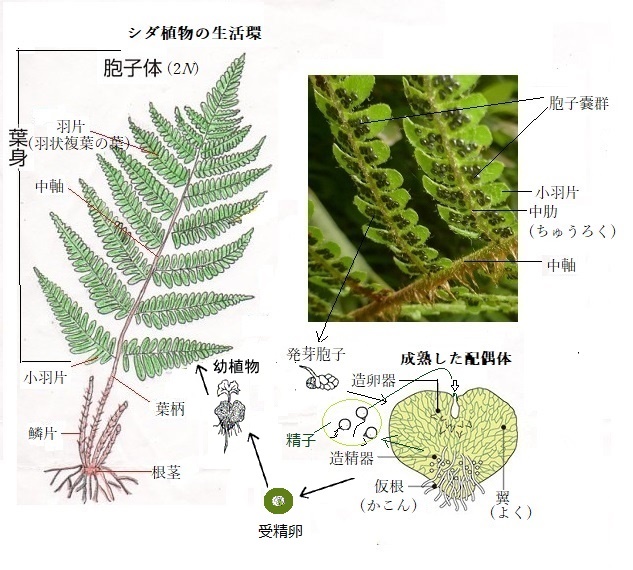 シダ植物やコケ植物は花が咲かない、つまり種子を作らない。シダ植物は、胞子で増える。葉の裏に、丸い小さな粒が列状に並ぶ、これは胞子が詰まった胞子嚢の集まりである。
シダ植物やコケ植物は花が咲かない、つまり種子を作らない。シダ植物は、胞子で増える。葉の裏に、丸い小さな粒が列状に並ぶ、これは胞子が詰まった胞子嚢の集まりである。胞子嚢がはじけると、中の胞子が湿潤な崖・岩・地面へ飛ばされる。
岩などに着生した胞子が発芽し、新しいシダへと成長していく。
シダ植物は、侵入・定着の条件として、湿潤な環境を好むが、生育する環境は、乾燥した場所でもよく適応する。
シダの葉を裏返してみると、緑色・黒色・茶色など粒状ものが並んでいる。ピンセットでめくると、小さな粒が飛び出してくる。胞子が入った胞子嚢が、バネのようにはじけて、胞子を飛ばす。 胞子は湿潤な岩などに着生すると発芽する。直径1cmほどの大きさになる。これが前葉体である。裏側の中央にある突起が造卵器で、その中に卵細胞ができる。その左下には、丸い造精器がいくつかある。ここで精子が作られる。
前葉体が水に浸ると、精子が放出される。しばらくする、精子には鞭毛があるため、活発に泳ぎ回る。 造卵器の入り口に集まってきた精子は、造卵器の管を上り、その付け根にある卵細胞にたどり着き受精する。受精した卵細胞は、細胞分裂を繰り返し、大きく伸びて成長する。胞子体の誕生である。胞子体が大きく成長したものが、通常、観察されるシダである。前葉体で芽生えた胞子体は、成長して胞子を作るようになる。
目次へ