| Top 車山高原 車山高原お知らせ 車山ブログ
|
 酸素呼吸(好気呼吸) 酸素呼吸(好気呼吸)FADH2は、NADHと同様、高エネルギー電子と水素の運搬体である。 NADHやFADH2の転移しやすい高エネルギー電子に蓄えられたエネルギーは、酸化的リン酸化によりATP合成に使われる。 酸化的リン酸化は、ミトコンドリア内膜で起こり、真核細胞における食物の酸化的異化のうちで大気中のO2を直接必要とする唯一の過程である。 クエン酸回路については、反応を進めるのに必要な大気中のO2が、廃棄物として放出される CO2と結合していると誤解されがちだが、実際は、クエン酸回路に入ったアセチル基をCO2にするのに必要な酸素原子は、大気中のO2ではなく地中のH2Oから供給される。 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応
| 目次 |
| 1)植物のストレス |
| 2)植物の水分状態water status |
| 3)馴化acclimationと適応adaptation |
| 4)環境馴化hardening off |
| 5)好気呼吸 |
植物重量・草丈・種子数は、合わせて植物の「全バイオマスtotal biomass(生物体量) 」を構成する。植物の最適な生育条件とは、その植物の植物重量・草丈及び種子数で測定される、成長と生殖の潜在能力が最大限に発揮され時、それらが最大の数値に達した際の諸条件と定義できる。
ストレスstressとは、いかなる環境条件であろうとも、植物が最大限の遺伝的能力を発揮できない条件と定義できる。例えば、光強度が減少すると、植物の光合成活性は低下するので、その植物のエネルギー供給能力が弱まる。このような状況では、植物は生合成速度を遅らせ成長速度を低下させる。あるいは、デンプンのかたちで貯蔵された貯蔵食物をとりくずすことで凌ぐことができる。
水利用の制限も同様に、成長に有害な効果をもたらす。植物が水ポテンシャルの低下を凌ぐ方法の一つは、気孔を閉じて蒸散による水分の損失を防ぐことである。ところが、気孔を閉じれば、二酸化炭素の取り込み量が低下するため、光合成も低下し成長が抑制される。
発生・成長・エネルギー生産・イオンと栄養のバランス及び貯蔵など、植物の各過程が環境の影響を受けながら互いに相互作用する。各過程の相互バランスは、植物ゲノムにコードされている。環境を感知し適合するためのセンサーやシグナル伝達経路の支配を受けている。
環境ストレスシグナルは、様々で、植物は各過程に配分するエネルギーを調節しながら、外部環境の変化に最も適合した、新たな内部環境、つまり生存に適した状態に調節する性質がある。その調節には、自ずと限界がある。イネは穏和な乾燥には耐性があるため成長の阻害とはならないが、厳しい乾燥下では、栄養成長が困難になる。
ストレスのない条件では、成熟した植物は栄養成長の遺伝的な発生プログラムを完了してから、生殖成長へと移行する。ストレス条件では、栄養成長プログラムは未完遂のまま終了し、直ちに生殖成長相が開始されることが多い。この場合、植物は完全に成長しない小型のまま、開花・受粉・結実を行う。準最適条件では、植物の葉の数が減少し光合成産物の供給が低下するので、種子の数やサイズが減少する。日照不足や栄養条件の悪化などによる植物の矮小化が、顕著に表現して見せる。
植物は、非生産的ストレス下においても、潜在的生殖能力を最大限発揮する。独特な発生径路をたどるが、それは生活環に大きく依存する。一年生植物annual plantでは、1シーズンでその生活環を達成する。そのシーズンの環境条件が異常であっても、代謝や発生プログラムを調節して、生殖能力のある種子を最大限に生産しようとする。
多年生植物perennial plantでは、多数のシーズンにまたがる能力を秘める種子を生産できるはずが、環境条件が満たされなければ、翌年まで植物の生存を全うさせるため、種子の生産を犠牲にしてまで、最適な食物貯蔵量を確保する、代謝と発生のプログラムを調整する。
目次へ
 隣り合う細胞間の重力ポテンシャルの差は、浸透ポテンシャルや圧ポテンシャルと違い、無視できるほど小さい。そのため、細胞レベルの水輸送では考慮されないことが多い。
このような場合、水ポテンシャルの式は、
隣り合う細胞間の重力ポテンシャルの差は、浸透ポテンシャルや圧ポテンシャルと違い、無視できるほど小さい。そのため、細胞レベルの水輸送では考慮されないことが多い。
このような場合、水ポテンシャルの式は、Ψ = Ψp + Ψs と簡略化される。
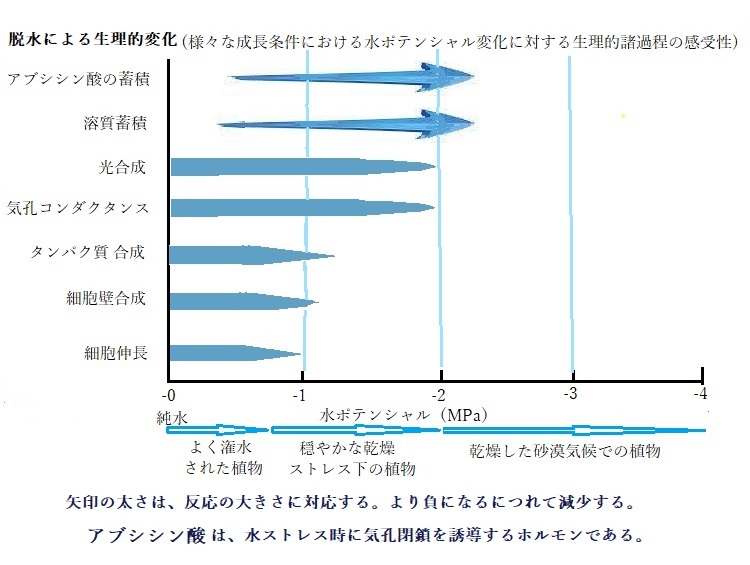
植物の生理的な諸過程は、その水分状態に影響される 水ポテンシャルは、植物の「水分状態water status」の尺度となる。植物は大気への蒸散により水が絶えず失われているため、常に水が不足がちである。
乾燥条件下では、植物の成長や光合成が阻害されなど、生理的変化が起こる。その生理過程がどのようであっても、水不足による感受性の殆どは、置かれた環境下に対応して、水を利用する様々な手段を講じている。
水不足に最も影響されるのが、水ポテンシャルの低下によって減少する細胞伸長である。多くの植物では、水の供給不足によりシュートshoot(1つの茎頂分裂組織に由来する茎と茎の付属物としての葉)の成長と葉の展開が阻害される。その一方、水を求めて根の伸長が促進される。それらは、水ポテンシャルの低下に適応するためで、水が不足すると、植物は、各組織における生理活性を維持するためにコストをかける。しかし、乾燥は植物の生理的諸過程に、絶対的な制約となる。しかも、それらの水ポテンシャルは、植物種により様々に異なる。
光合成を行う植物は、水分消失と脱水の脅威は避けられない。乾燥を避けるため、水は根により吸収され、植物体を通して輸送される。水が不足すると、植物は、溶質を蓄積するためにエネルギーを使い、膨圧を維持し、根などの非光合成器官の成長に投資し、水の獲得能力を高めようとする。また、木部道管を構築し、大きな水の張力を保持する。このような水不足により生じる生理的応答に必要なコストと、様々な環境条件下で、成長などの生理的な諸過程を進行させることとの利益とのバランスが働いている。
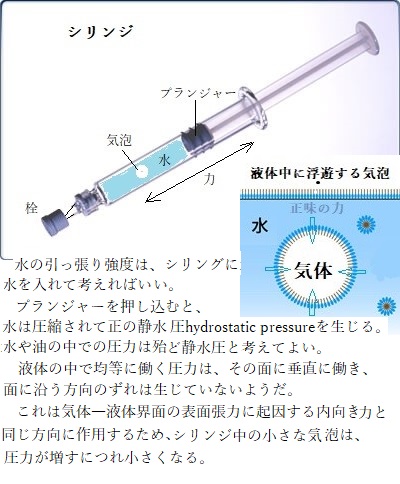 Ψpは、圧ポテンシャルpressure potentialと呼ばれる、水の自由エネルギーに対する静水圧(密閉空間内に水を押し込むときに発生する圧力)の効果を表す。植物の細胞壁は、細胞内に大きな正の静水圧(膨圧)を発生させる。膨圧は植物の多くの働きに関わる。生きた細胞では、Ψp > 0以上のレベルと想定される。
Ψpは、圧ポテンシャルpressure potentialと呼ばれる、水の自由エネルギーに対する静水圧(密閉空間内に水を押し込むときに発生する圧力)の効果を表す。植物の細胞壁は、細胞内に大きな正の静水圧(膨圧)を発生させる。膨圧は植物の多くの働きに関わる。生きた細胞では、Ψp > 0以上のレベルと想定される。負の膨圧は、細胞壁が木質化した生細胞で起こるのだろうか?
正の膨圧Ψp > 0は重要で、植物細胞は、膨圧により細胞壁を伸展させ成長を促進する。水不足による膨圧の喪失は、水ストレスにより細胞伸長が抑制され、水不足による成長抑制が細胞の浸透ポテンシャルに左右されることが分かる。溶質は水を薄めることにより、水の自由エネルギーを減少させる。
正の膨圧が重要である第2の理由は、膨圧が細胞や組織の機械的強度を増加させるからである。
伸展により活性化される情報伝達分子が、細胞膜に存在することから、植物細胞は、膨圧よりも細胞の体積変化を通して、水分状態の変化を感受しているようだ。
植物は、過酷な環境で成長し生殖するが、そこには様々な非生物的・化学的・物理的諸要素が含まれ、それらは時間及び地理的位置によって変化する。
植物の成長に特に影響するのが、光の強度・光質light quality(波長分布特性)・光周期(波長)、土壌からの水の利用度と外気湿度、二酸化炭素と酸素、土壌栄養素の含量と利用効率、日照時間と温度、風の強度、及び植物にとっては毒物とも言える重金属や塩分などがあげられる。これらの非生物的要因が、ひとたび正常値から逸脱すれば、植物は、生化学的にも生理学的にも過重な負担に晒される。植物は移動して、その悪環境から逃れる手立てが無いため、その度重なる非生物的ストレスの多い状況をやり過ごさなければならない。そのため、成長や生殖を維持するに必要な様々な生理と発生のプロセスを進化させてきた。
植物という複雑な生物を組織形成する細胞の核は、細胞の代謝・成長・分布を制御する主要な遺伝情報を含む細胞小器官である。核に存在する遺伝子とその介在配列を合わせて核ゲノムnuclear genomeと呼ぶ。植物の核ゲノムは、モデル植物として有名なアブラナ科のシロイヌナズナが1.2×108(1.2×1億)塩基対であるに対して、ユリ科のバイモ属の植物では、1.0×1011(1.0×1,000億)塩基対と、その大きさに大きな幅がある。更に遺伝情報は、核ゲノム以外に、2つの半受動的な細胞小器官である色素体とミトコンドリアにも含まれている。
植物は、多くの生物と同様、タンパク質・調節分子・シグナル伝達因子、及び生合成化合物などが合わせて、無限とも思える相互リンクした径路やネットワークを形成している。通常の生育環境下では、環境からのインプットを成長や生殖に必要な遺伝的アウトプットと適合させるために、これらの異なる化学や物理的径路とシグナル伝達ネットワークを協調的に働かせている。
ひとたび、好ましくない外部環境に晒されると、植物は、ストレスの負の衝撃を最小限にして代謝の平衡を維持する。植物は、この複雑な相互作用を働せて「恒常性」を維持しようと調整する。この誘導される防御応答だけではない。植物には、即座に稼働できる恒常的防御機構が常に準備されている。
恒常性homeostasis(hòumiəsˈteisis)とは、生物は外部環境の影響により、体内の環境(内的環境)が常に変化するような圧力を受けているが、この内的環境を一定に、あるいは生存に最適の状態に保つ性質のこと言う。植物は、様々な環境の変化に対応して、内部状態を一定に保ちながら、生存を維持する。動物における血液の性状の一定性や体温調節などもその例と言える。
種子といえども、種皮によって外界と隔絶された状態になってはいない。種皮の厚さ・堅さなどの違いは様々であるが、多くは細胞壁成分と同じで、主成分は多糖類で水に高い親和性を持つ物質から出来ており、外部環境に対しては、決して閉鎖的ではない。種子が守るものは、次世代の植物となる胚である。種子形成は、胚発生期(細胞分裂➝細胞分化➝細胞拡大)、成熟期(貯蔵物質の集積)、乾燥期と段階的に進行する。胚発生は通常の水分状態で行われ、一定の発生段階に達すると発生を中止する。
胚発生の中期から植物ホルモンの一つであるアブシシン酸abscisic acid C15H20O4が大量に貯まるようになる。アブシシン酸は、休眠中の種子・樹芽・球根などには多量に含まれている。特に、種子では胚の未熟発芽を防ぎ、完熟種子の休眠状態を維持する。このアブシシン酸がいろいろな作用をする。まずは、胚の乾燥耐性を高める作用である。増加したアブシシン酸は、中期から後期の発生胚の遺伝子発現に影響を与え、胚に耐乾燥性を与える特殊なタンパク質の合成を促進する。同時に一定の発生段階に達した胚を休眠状態に移行させる。次の成熟期に入ると、貯蔵物質の蓄積を促す。
成熟期中期頃から種子の乾燥が始まり、胚も乾燥する。そのため、種子内の塩濃度が上昇することは十分考えられ、また、細胞外の塩濃度が高まれば、細胞内への吸収量も増加することが考えられる。アブシジン酸は塩耐性を強める作用があることも明らかにされている。アブシジン酸の作用で、種子は休眠を保ち、塩や乾燥に耐えるようになっていると考えられている。
目次へ
ストレスstress(物理的・化学的な圧力や障害)とは、刺激により引起される非特異的な生体反応を言う。苦痛・苦悩・難儀・災難などを意味する distress が短くなった単語とされる。生体に加わる力をストレッサー、それによって起る生体の反応をストレスと呼ぶ。
個々の植物が、生存と生殖の能力を高めるため、温度・日照時間・標高・風などの気候や環境の変化に応答して、生理機能や形態を変化させて順応することを馴化という。こうした反応には、新たな遺伝的な変化を必要としない。環境ストレスに繰り返し晒され、植物のストレス応答性が向上する場合の応答を馴化と呼ぶ。馴化によって起こる変化は、環境変化に比較的速やかに応答して起こる、生理機能や形態の非永続的な変化でためか可逆的で、環境ストレス状態が変化・解消すれば元に戻りうる。元の環境に戻せば、植物は元の生理状態に戻る。
ある種の植物は、光・高温・低温・乾燥・塩に関して準限界ストレスを短期間経験することにより馴化し、その後より厳しいストレスにも耐性となる。その後、ストレスが解消すると細胞は元の生理状態に戻る。光・高温・低温・乾燥・塩ストレスなどの物理化学的ストレスに馴化する植物が、数多く知られて、その植物の馴化は、新規の遺伝子発現を伴うものもある。
一方、遺伝暗号まで変化はさせないが、遺伝子発現を変化させるエピジェネティックepigenetic機構が働くと、馴化応答の持続時間を延長し、遺伝させることができる。エピジェネティック機構とは、DNA塩基配列の変化を伴う「突然変異」とは異なり、DNAの配列に変化が生じないまま、環境要因の架け橋となって応答し、遺伝子発現を制御・伝達する機構である。
植物集団全体の遺伝的変化が、環境の選択圧によって多くの世代にわたって固定されれば、その変化は適応と呼ばれる。その結果、構造・習性・特性などを遺伝的に変更・調整し、環境変化に応答して自らの状態を改善する。ストレスの適応は、新規の遺伝子発現による変更・調整であるから、後代にわたる遺伝的改変となる。
極端な非生物的ストレス環境の適応は、例えば、蛇紋岩土壌における植物の生殖に見られる。鮮やかな緑色をした蛇紋岩は、翡翠と見紛うほど美しい。蛇紋岩は、マグネシウムを多量に含む岩石である。その多量のマグネシウムが、植物の水分吸収能力を低下させると言う。また、蛇紋岩が風化して形成される土壌は、土壌粒子間の結合が弱いため、土壌中に水分を含むと液状化や地層の流動を起こしやすい。蛇紋岩地層の特に南向きの切り土法面において、緑化は困難を極め、外来牧草などによる種子吹付は芽生えるものの、成長できずに失敗に終わることも多く、厚層基材吹付工も高い効果を発揮しにくいと言う。
蛇紋岩地域であっても、土壌が厚く堆積している場所であれば、時として植物に障害を与えることになるニッケルなど重金属による母岩の悪影響が弱まり、普通の植生が可能になる。土壌が薄い急傾斜地や、母岩の影響を直接受けることになる露岩地では、殆どの植物にとって、厳しいストレス条件となる。
単純な移植実験により、蛇紋岩土壌では、適応集団のみが生育し増殖することが分かっている。どのような遺伝子的交雑により、どの遺伝子の変化が、安定した適応に寄与しているかも既に明らかになっている。
ある特定の環境変動に応答する植物の適応進化では、一般的に、その環境条件に内在する有害な影響を回避する遺伝的変化が、その過程で生じている。そのため、もっとも蛇紋岩地域らしい植生が発達する。このような場所の特異な環境条件に耐えることのできる植物や、根の浅いツツジ科植物やアカマツなどが、乾燥に耐えながら貧弱な植生集団を作る。その一方では、通常の土壌で生育している植物集団から遠く離れた蛇紋岩土壌で、その遺伝的近縁種が適応して集団をつくることが少なくない。蛇紋岩地帯の植生は、他の種との生存競争が低いことから、遺伝的に特異な植物群からなっていることが多い。
北海道の日高山脈支稜線西南端に位置するアポイ岳の高山植物群落では、「アポイツボスミレ」や「アポイアズマギク」「ヒダカイワザクラ」といった地元の名を冠した固有植物が多い。ミズバショウの花咲く尾瀬ヶ原の彼方にある残雪の至仏山(しぶつさん)も同様に有名である。山体が蛇紋岩でできているため、オゼソウ・ホソバヒナウスユキソウ・タカネバラなど、特殊な蛇紋岩植物と呼ばれる植物群が生育する 低い海抜高度では、貧弱なアカマツ林や、ツツジ類が有占する低木群落になることが多い。
適応は、生物学的には進化の選択圧による種の自然選択の結果であり、生物が自らに積極的に働きかけて遺伝的変化を起こしたわけではない。馴化は、温度・日照時間・標高・風などの気候や環境の変化に対して、植物が生理的に順応する。適応によって起こった変化は、時を要する進化的経過の結果でありため、子孫に受け継がれる形質となるのに対し、馴化によって起こった生理的変化は、環境変動への速やか対応を可能にし、しかも可逆的である。
例えば、生葉の光合成を測定する際に、前もってサンプル葉を暗処理する操作を暗馴化(dark acclimation `ækləméɪʃən)とぶ。暗所で生育させた植物を徐々に光強度を上げてゆき、強光でも枯れなくなるようにする操作を明馴化(light acclimation)と呼ぶ。これに対し、林床に生育する、光合成量・呼吸量とも小さい耐陰性の高い植物(陰性植物)は、その光環境に適応しているという。
目次へ
植物栽培における馴化の一例として、ハードニングオフhardening offが知られている。苗木などを、寒さや乾燥などによる悪条件になりやすい環境に馴化させることを言う。ハードニングhardeningとは、植物本来の脆弱性を減らすことで、カエデ・アヤメ・トマトといった多くの植物は、気温が数日から数週間かけて夜ごとに徐々に低下していくなら、氷点下の気温に耐えることができる。この過程もハードニングオフと呼ばれ、水分量の減少と糖含量の増加によって樹液の融点を低下させるなど、様々な変化が関与している。
園芸愛好家は、野外で果樹を種子から生育させるより、その成長速度を上げるため、最初は、鉢植えにして屋内の最適条件下で苗になるまで生育させる。その後、一日のある時間帯だけ屋外に鉢を移動し、植物が屋外環境に十分に馴化してから屋外に移植する(環境馴化)。
前年の秋に果実が実り、種ができる。秋に発芽しても、そのあとの冬の寒さで枯死してしまうため、通常、発芽せずに休眠する。この時点の休眠を一次休眠と言う。夏に育つ植物は、一般に冬の寒さで一次休眠から覚める。しかし冬の寒い時期や、乾燥した時期には発芽しない。これを環境休眠と言う。
春、暖かくなり、水分やその他の条件が整えば、ようやく休眠から覚めて発芽する。しかし、種子が親植物から離脱・散布されてある期間を経た後も、乾燥・低温・低酸素などによる、発芽を妨げる環境条件が続けば発芽しない。これを二次休眠と言う。二次休眠は、その原因となった環境要因が取り除かれた後も継続する。
その後、秋が過ぎ冬の寒さで、再び、休眠から覚め、春の発芽の環境条件が整うのを待つ。これが、夏野菜の発芽までの道のりである。
逆に秋に発芽する冬野菜は、一次休眠が夏の暑さとなる。白菜の生育適温は15~20度、収穫期は10~1月、その種子の二次休眠は、悪い環境に置かれた時などより誘導される。このように発芽したり、休眠したりするのは、環境と種自体の状態による。環境としては、温度と水分の影響が大きい。どちらも、生き物が活動する上では、極めて重要な要素となる。
また、植物は酸素呼吸して、生育に必要なエネルギーを作り出す。酸素を必要とする好気呼吸は、ほぼすべての真核生物に共通しており、動物とはいくつか点で異なるも、植物の呼吸の概略は、動物や他の好気性真核生物と類似している。
好気呼吸とは、細胞呼吸であり、グルコースなどの呼吸基質が完全に酸化分解されて、二酸化炭素と水とに分解されと、解糖系(嫌気呼吸)に続く好気呼吸の主要な反応経路がある、細胞内のミトコンドリア・マトリックにおけるクエン酸回路でなされる。グルコース分子から解糖系およびクエン酸回路で生じた水素が、最後に酸素と化合して水となり、この呼吸よって自由エネルギーが放出され、そのエネルギーが、一時的にATP(adenosine triphosphate;アデノシン三リン酸)として蓄えられる。そのATPが、成長や生命維持に必要な細胞内の反応に利用される。
ある特定の環境条件に対応する植物の適応機構の進化は、一般的に、その環境条件に内在する有害な影響を回避する過程で起こる。英国イングランド南西地方のヒ素汚染鉱山地帯に適応して生育しているイネ科のシラケガヤ(白毛茅、学名:Holcus lanatus)の集団は、鉱山により汚染された土地のヒ素の毒性を回避できる遺伝子的変化をしている。シラケガヤは、ヨーロッパ原産で、世界中に帰化植物として広がり、日本では北海道から九州まで分布している
 。適応能力が発達した雑草だ。
。適応能力が発達した雑草だ。植物が極限環境に耐えられるか否かは、遺伝的適応と馴化の両方が関与する。ヒ素に耐性のあるシラケガヤの集団では、遺伝的適応により、ヒ素の吸収が低減されるが、完全に停止できたわけではない。その後もヒ素は、ある程度蓄積されている。
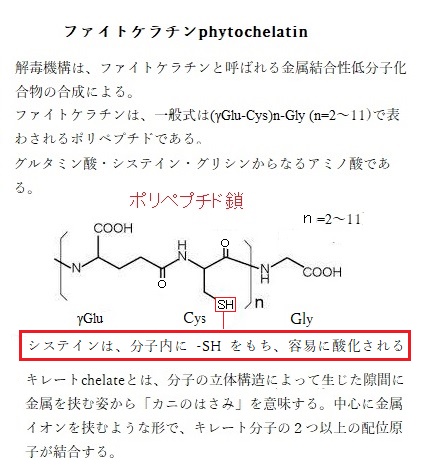
ヒ素に適応したシラケガヤは、非適応植物と同様の生化学的メカニズムを用いて、組織中に蓄積されたヒ酸の毒性を軽減する。この解毒機構は、ファイトケラチンphytochelatinと呼ばれる金属結合性低分子化合物の合成による。
ヒ素汚染鉱山廃棄物の環境で生育するために、シラケガヤが、耐性集団特異的な遺伝的適応と馴化の双方を獲得した典型例と言える。
キレートchelateとは、分子の立体構造によって生じた隙間に金属を挟む姿から「カニのはさみ」を意味する。中心に金属イオンを挟むような形で、キレート分子の2つ以上の配位原子が結合する。配位結合とは、形成する2個の原子の一方のみから、結合電子が供与される結合のことで、その化合物となる配位結合は、多くの錯体にその例がみられる。
錯体とは、金属または金属類似元素の原子・イオンの周囲に、配位子と呼ばれる原子・イオンまたは原子団が立体的に結合し、一つの原子集団を作る立体的分子構造を呼び、この結合の中核をなす配位子をキレート剤と言い、それにより形成される環構造をキレート環と言う。
キレート分子は、硫黄・窒素・酸素など、配位に必要な異なる原子を持っており、これらの原子は、キレートするイオンに対して、それぞれが異なる親和性を持っている。その結果、イオンを自らが包み込むようにして錯体を形成することで、キレート分子はイオンの化学活性の能力を低下させ、それによりイオンの潜在的な毒性を弱める。その後、錯体は、植物体の他の場所に移動するか、あるいは細胞質の液胞に貯蔵される。
ファイトケラチンは、一般式は(γGlu-Cys)n-Gly (n=2~11)で表わされるポリペプチド(アミノ酸の鎖)である。生物に対するカドミウムや水銀などの毒性を防御のために、植物や一部の微生物が合成する低分子のチオール化合物で、グルタミン酸・システイン・グリシンからなるアミノ酸である。重金属とキレート化合物を形成すれば、その重金属を無毒化できる。このチオール化合物は、ファイトケラチン合成酵素PCSによって合成される。植物や一部の真核微生物などに存在する。この酵素は、カドミウムなどの有害金属によって活性化し誘導合成され、細胞内でその濃度を上昇させる。
チオールは R−SH(Rは有機基質)であらわされる構造を持ち、官能基として呼称される場合は、そのままチオール基(-SH)と呼ばれる。カドミウムはSH残基(システイン残基;phytochelatin)と結合する性質があり、ファイトケラチンのSH残基がカドミウムと結合し、カドミウムの毒性を軽減する。残基とは、重合体の主鎖以外の部分で、もとの単量体に相当する部分を呼ぶ。
ファイトケラチンのチオール基は、カドミウムやヒ素などの微量元素のイオン配位子として働く。ファイトケラチン-金属錯体が形成されると、液胞に輸送されて液胞で貯蔵される。ファイトケラチンの合成は、カドミウムやヒ素に対して有効な抵抗を示す。キレートする以外では、液胞内や細胞外へ金属イオンを能動輸送することも、金属に対する内部耐性にも貢献する。錯体が、根からシュート組織へキレートイオンを長距離輸送することは、シュート組織に金属類を過剰蓄積する重要な過程となる。鉄キレート剤であるニコチンアミン、及び遊離アミンのヒスチジンは、この輸送過程で金属キレート剤として働く。
海水の影響のある地域では、藻類である海藻は普通に見られるが、種子植物はごく少ない。種子植物・シダ植物・コケ植物などの陸上植物は、すべて淡水から陸上で進化したものと考えられている。そのため、塩分への耐性が弱く、台風で海水の飛沫を浴びると、広く植物が枯れる現象(塩害)が見られる。塩分・アルカリストレスは、農作物の減産を引き起こす重大な環境要因である。
馴化の例として非塩性植物glycophyteの耐塩性機構としては大きく浸透圧ストレスとイオンストレスに対するメカニズムがあり、関与する候補遺伝子群に関する研究成果が実りつつある。ただ、塩生環境で生育できるような遺伝的適応はしていないようだ。環境の塩濃度の上昇により起きる生理的変化に対処するため、塩濃度の上昇に晒されるとストレス応答機構を活性化させている。
塩濃度の浸透圧ストレスを軽減するメカニズムには、浸透圧調節・水吸収の促進・蒸散の抑制などがある。塩分ストレスは、細胞内イオン、特にK+やNa+のバランスや分布を乱す。特に細胞質におけるNa+濃度が上昇すると、余分なNa+濃度により酵素活性が損なわれ、細胞における正常な生理・生化学的代謝活動が抑制される。イオンストレスに対しては、体内特に地上部へのNa+の移動・蓄積を低減する回避と、Na+を体内組織内に取り込んだ場合の組織耐性の2つのタイプに分けられる。前者には根や葉などからのNa+の体外への排出、地上部から根への還流、組織の含水量の増加による希釈などがある。後者では、液胞での隔離と排出、カウンターイオン溶質の合成、K+によるNa+の置換などがある。
高等植物では、酵素自体の耐塩性に、大きな差は認められていないが、何らかの手段で酵素活性をNa+から保護している。根が高塩環境に晒されると、短時間で地上部が反応することから、根から地上部への何らかのシグナル伝達機構があることが分かる。
環境の塩濃度の上昇に晒されて起こる生理的変化に対処するため、ストレス応答機構を活性化させる。初期の根の反応では細胞質Ca2+の増加がみられる。Ca2+信号に依存する植物における耐塩・耐アルカリ信号の伝達経路であるSOS径路の関与が示唆されている。それは、塩高感受性の突然変異体Salt Overly Sensitiveの解析から解明されたシグナル伝達経路で、細胞からのNa+の排出と塩NaClによる毒性の低減に寄与する。
塩分ストレス下におけるSOS経路の反応過程は、植物が塩分ストレスにさらされると、細胞膜にある何らかの受容体によりストレス信号が識別され、細胞質内のCa2+濃度が上昇する。このCa2+信号によりSOS径路が活性化される。活性化されると、細胞膜や液泡膜に含まれるNa+/H+輸送タンパク質が、Na+を細胞から排出または液泡から分離し、細胞質におけるNa+イオン濃度を引き下げる。
目次へ
光合成は、すべての生物が必要とする基本的な有機成分を供給する。呼吸は、関連した炭素代謝とともに、炭素化合物に蓄えられた化学エネルギーを、厳密に制御された方法で、細胞内の必要性に応答して遊離し、それに伴い生合成に不可欠な多くの炭化化合物の前駆体も生産する。
動物細胞も植物細胞も、有機分子の化学結合に蓄えられた化学エネルギーに頼る。有機分子とは、植物が光合成の第2段階で自家消費用として作った糖や、動物が食餌により獲得する大小の分子の化合物である。生物は、このエネルギーを活用するため、緩やかな酸化、則ち制御された燃焼を行う。
酵素を要求する好気呼吸は、ほぼすべての真核生物に共通するが、植物の呼吸の概略も、動物や他の好気性の真核生物の細胞呼吸と類似し、解糖系・クエン酸回路・電子伝達系の3段階で進行する。しかし、植物の呼吸は、いくつかの点で動物のものと異なっている。
それを生み出した植物の功績により、地球の大気には、酸素が21%含まれている。その酸素は、エネルギー的に最も安定した形が、炭素と結合するCO2であり、水素と結合するH2Oである。細胞は、糖などの有機分子の炭素原子と水素原子を酸素と結合させている。これが酸化である。それぞれCO2 とH2Oに変えてエネルギーを獲得する。この過程が呼吸respirationである。
光合成と細胞の呼吸は、相補的な過程である。これは植物と動物の関係が一方的でないことを意味する。植物・動物・微生物は、地球上で長く共存してきたため、それぞれが互いの環境の一部として不可欠な存在になっている。光合成によって放出される酸素は、殆どの生物が行う有機分子の酸化分解の際に消費される。草木の葉が、光合成によって有機分子として取り込むCO2分子の一部は、動物や菌類、あるいは植物自体の呼吸によって、さらには化石燃料の燃焼によって、大気中に放出されたものである。このような酸素の利用は、生物間の境界を越えて、生物圏全体を巻き込む巨大な循環を形成している。
光合成と細胞呼吸は、生物界において相補的な関係にある。日光エネルギーを利用して大気中のCO2の炭素原子から、糖とそれから生成される巨大な脂肪やタンパク質などの有機分子が作られる。これらは他の生物の食物となる。光合成微生物を含む殆どの生物が行う細胞呼吸では、O2を用いて食物分子を酸化し、その炭素原子をCO2として大気中に放出する。この過程で生物は、生存に必要な使いやすい化学総合エネルギーを獲得する。
好気呼吸aerobic(eróʊbɪk) respirationは、還元された有機化合物が、厳密に制御された方法で酸化される生物学的プロセスである。呼吸によって自由エネルギーが放出され、そのエネルギーは、一時的にATP adenosine triphosphate(アデノシン三リン酸)として蓄えられ、成長や生命維持に関わる細胞の反応の動力に活用される。
 一般的には、グルコースなどが呼吸の基質として、細胞呼吸である好気呼吸に使われている。一方、植物細胞で使われる還元炭素の殆どは、二単糖のスクロースやその他の糖、有機酸、光合成から得られる3つの炭素からなるトリオースリン酸triose
phosphate;fάsfeɪt、脂質やタンパク質の分解によって得られる代謝産物などの化合物から得られている。
一般的には、グルコースなどが呼吸の基質として、細胞呼吸である好気呼吸に使われている。一方、植物細胞で使われる還元炭素の殆どは、二単糖のスクロースやその他の糖、有機酸、光合成から得られる3つの炭素からなるトリオースリン酸triose
phosphate;fάsfeɪt、脂質やタンパク質の分解によって得られる代謝産物などの化合物から得られている。呼吸の基質は、細胞で起こる他の過程で合成される。呼吸の径路に入るとサイトゾルcytosol(細胞質ゾル;細胞質のコロイド状の溶質で、細胞質からリボソームなどの細胞内小器官や、細胞内外の運動に必要な物理的力を発生させる繊維状構造の細胞骨格を除いた部分)やプラスチドplastid(未分化の色素体で、葉緑体など、どの色素体にも発達できる、植物の半自律的な細胞小器官である。糖や脂肪などの貯蔵、様々な種類の化合物の合成などを担う。核ゲノムとは別に独自のゲノムDNAを持つ。)にある解糖系glycolysis(glaikɑ'ləsis)と酸化的ペントースリン酸径路oxidative pentose phosphateは、糖をヘキソースリン酸やトリオースリン酸を経て、ビルビン酸のような有機酸に変換して、NADHまたはNADPHとATPを生成する。
解糖系は、サイトゾルとプラスチドの両方に局在する酵素によって触媒される一連の反応からなる。解糖はすべての生物、原核生物や真核生物で起こり、植物での解糖の主要な反応は、動物細胞のものと殆ど変わりない。解糖は、いくつかのソースから供給される炭水化物を代謝する。例えば、スクロースは六炭糖のヘキソースリン酸や三炭糖のトリオースリン酸を経て部分的に酸化され、主にピルビン酸などの有機酸になる。このプロセスで少量のエネルギーではあるがATPが得られ、還元力も還元型のNADH(ニコチンアミド ヌクレオチドnicotinamide nucleotide)として得られる。
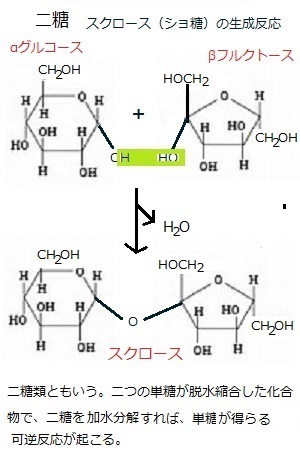 酸化的ペントースリン酸径路では、六炭糖のグルコース6-リン酸が、五炭糖のリブロース5-リン酸に酸化される。炭素がCO2として失われ、還元力は別の還元型のNADPH(ニコチンアミド ヌクレオチド)として保存される。その後、酸化的ペントースリン酸径路で引き続く平衡反応に近い反応によって、五炭糖のリブロース5-リン酸は、3個から7個の炭素原子を含む糖リン酸へと変換される。これらの代謝中間体は、ヌクレオチドなどの生合成径路で利用されるか、解糖系へ戻っていく。
酸化的ペントースリン酸径路では、六炭糖のグルコース6-リン酸が、五炭糖のリブロース5-リン酸に酸化される。炭素がCO2として失われ、還元力は別の還元型のNADPH(ニコチンアミド ヌクレオチド)として保存される。その後、酸化的ペントースリン酸径路で引き続く平衡反応に近い反応によって、五炭糖のリブロース5-リン酸は、3個から7個の炭素原子を含む糖リン酸へと変換される。これらの代謝中間体は、ヌクレオチドなどの生合成径路で利用されるか、解糖系へ戻っていく。ミトコンドリアの内膜で区切られたマトリックで、ビルビン酸が段階的な酸化を経てCO2にまで完全に酸化される。この過程のスクロース分解で得られた大部分の還元力は、スクロース1分子あたり16分子のNADHと4分子のFADH2と少量のエネルギーATPを生み出す。有機酸は、クエン酸回路citric acid cycle(citric;sɪ́trɪk;クエン酸の)で酸化され、そのとき生産されるNADHとFADH2は、ミトコンドリア内膜にある一連のタンパク質複合体である電子伝達鎖electron transport chainで、流動的な電子伝達成分(ユビキノンとシトクロムc)と繋がり、NADHからO2へ電子伝達を触媒する。この過程で多量の自由エネルギーが放出され、酸化的リン酸化を行なうATP合成酵素にATP合成に必要なエネルギーを供給する。一部は電気化学ポテンシャルであるプロトン(水素イオンH+)勾配proton gradientとして保存される。それは膜を横切るプロトンの濃度勾配で、葉緑体やミトコンドリアで利用され、ATP合成の駆動力になる。
酸化的リン酸化oxidative phosphorylationでは、電子はミトコンドリアの内膜に埋め込まれた一群のタンパク質合体からなる電子伝達鎖に沿って伝達される。このシステムにより、電子が、解糖系・酸化的ペントースリン酸径路・クエン酸回路で生じたNADHや関連した分子から酵素へ伝達される。この電子伝達系は、大量の自由エネルギーを放出し、そのエネルギーの大部分は、ATP合成酵素によって触媒されるADPと無機リン酸からATP合成を通して保存される。電子伝達鎖の酸化還元とATP合成は、総括して酸化リン酸化と呼ばれる。

アミノ酸・グリセロールなどの糖でない物質から糖を作る糖新生gluconeogenesisは、解糖とは逆反応による糖の合成である。脂肪酸の酸化形式の1つのβ酸化(β-oxidation)と炭素数2つのグリオキシル酸glyoxylate(化学式:CHOCOOH)は、その代謝に特化したマイクロボディmicrobodyであるグリオキシソームで分解される。糖新生は、単純に解糖系の逆反応で起こるわけではない。その経路は異なっており、異なる反応と異なる酵素によるものである。
マイクロボディmicrobodyは、1つの代謝機能に特化した一種の膜に包まれた球状の細胞小器官である。グリオキシソームは、ミトコンドリアとオイルボディに密着して、中間代謝物を交換する。脂肪を貯蔵物質とする種子の発芽過程で出現する細胞内の小顆粒(顕微鏡により細胞内に見られる粒状の構造)で、酸化酵素を含み、脂肪を糖に転換する。
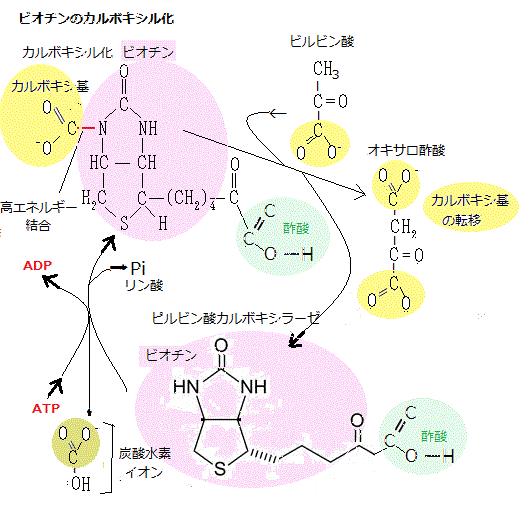
グリオキシル酸回路を巡ると、2分子のアセチルCoAから水素2原子がとれて、1分子のコハク酸が合成される。植物ではこの回路の酵素をグリオキシソームと呼ぶ。植物がある条件下に置かれると、この回路を利用して、脂肪の糖への転換が活発に行われる。
 グリオキシル酸回路で代謝された1分子のコハク酸は、グリオキシソームからミトコンドリア・マトリックに運ばれ、そのクエン酸回路でコハク酸➝フマル酸➝リンゴ酸となる2つの反応でリンゴ酸に変換される。リンゴ酸は、ミトコンドリア内膜にあるジカルボン酸トランスポーターにより、コハク酸と交換にサイトゾルへ輸送される。リンゴ酸は、そこでリンゴ酸脱水素酵素によって酸化されオキサロ酢酸になる。オキサロ酢酸は解糖系の逆反応(糖新生)で、PEPカルボキシキナーゼによって、ATPを使いPEP(ホスホエノールピルビン酸phosphoenolpyruvate)とCO2に変換される。PEPから糖新生径路を経てグルコースが合成される。スクロースは、この過程の最終産物で、子葉から成長している芽生えの各組織に転流され還元炭素の主要な形態である。
グリオキシル酸回路で代謝された1分子のコハク酸は、グリオキシソームからミトコンドリア・マトリックに運ばれ、そのクエン酸回路でコハク酸➝フマル酸➝リンゴ酸となる2つの反応でリンゴ酸に変換される。リンゴ酸は、ミトコンドリア内膜にあるジカルボン酸トランスポーターにより、コハク酸と交換にサイトゾルへ輸送される。リンゴ酸は、そこでリンゴ酸脱水素酵素によって酸化されオキサロ酢酸になる。オキサロ酢酸は解糖系の逆反応(糖新生)で、PEPカルボキシキナーゼによって、ATPを使いPEP(ホスホエノールピルビン酸phosphoenolpyruvate)とCO2に変換される。PEPから糖新生径路を経てグルコースが合成される。スクロースは、この過程の最終産物で、子葉から成長している芽生えの各組織に転流され還元炭素の主要な形態である。グリオキシソームやオイルボディは、細胞内膜系に由来するにも関わらず、独立して成長・増殖・融合する細胞小器官である。オイルボディは、脂質を貯蔵する細胞小器官で、チューブ状の小胞体から出芽して分離する。オレオシンと言うオイルボディ特異的なタンパク質を含むリン脂質半単位膜、つまり単層のリン脂質が、油脂を貯蔵している。多くの植物は、種子の発達過程で多量の油脂を合成して貯蔵している。これらの油脂が細胞小器官のオイルボディに蓄積され、種子が発達するにつれオイルボディは分解するが、その際には脂質酸化酵素を含む細胞小器官であるグリオキシソームに結合している。
 植物が獲得する化学総合エネルギー、例えばATPは、細胞内のミトコンドリア・マトリックスにあるクエン酸回路で、全体として8個の反応を通過する回路が1周するたびごとに、末端のリン酸基をADPに渡しATPを作る。
植物が獲得する化学総合エネルギー、例えばATPは、細胞内のミトコンドリア・マトリックスにあるクエン酸回路で、全体として8個の反応を通過する回路が1周するたびごとに、末端のリン酸基をADPに渡しATPを作る。FADH2はNADHと同様、高エネルギー電子と水素の運搬体である。 NADHやFADH2の転移しやすい高エネルギー電子に蓄えられたエネルギーは、酸化的リン酸化によりATP合成に使われる。
酸化的リン酸化はミトコンドリア内膜で起こり、真核細胞における食物の酸化的異化のうちで大気中のO2を直接必要とする唯一の過程である。
生物体がエネルギーおよび物質を外部から取り込み(同化)、体内で化学的に変化させる。不用なものは外部に放出する。
食物の主要成分であるタンパク質・脂肪・多糖を、真核細胞がエネルギー源や他の有機分子の単位体として利用するためには、これらを小分子に分解しなければならない。酵素が複雑な有機分子を、簡単な有機分子に分解する過程を異化(catabolism)と言う。
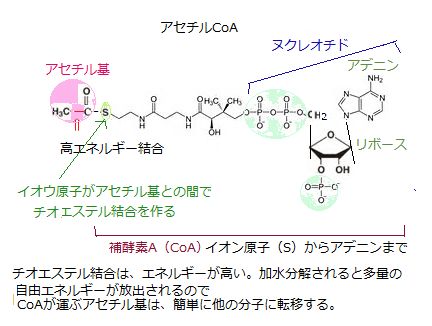
異化の最終段階は、ミトコンドリア・マトリックで起こる。アセチルCoAのアセチル基がオキサロ酢酸分子に転移してクエン酸ができる。ここからクエン酸回路という一連の反応に入る。
転移したアセチル基は、この反応でCO2に酸化され、その際に大量のNADHが生じる。最後に、NADHの高エネルギー電子が、ミトコンドリア内膜にある酵素群である電子伝達系をたどる。
 ここで電子の移動に伴って放出されるエネルギーが、酸化的リン酸化、即ちATPの生成とO2ガスの消費を行う。
ここで電子の移動に伴って放出されるエネルギーが、酸化的リン酸化、即ちATPの生成とO2ガスの消費を行う。この酸化によって放出されたエネルギーで、細胞の殆どのATPが作られる。この最終段階で、糖や脂肪の分解で取り出されたエネルギーが、細胞内で使われやすいATPというエネルギーの塊に変換される。
全体としてグルコースや脂肪酸のH2OとCO2への分解で、理論的に得られるエネルギーの約半分が捕捉され、 エネルギー的に起こりにくい ADP + P1 ➡ ATP の反応を進めるのに使われる。
クエン酸回路については、反応を進めるのに必要な大気中のO2が、廃棄物として放出されるCO2と結合していると誤解されがちだが、実際は、クエン酸回路に入ったアセチル基をCO2にするのに必要な酸素原子は、大気中のO2ではなく地中のH2Oから供給される。
地球上に最初に誕生した細胞は、光合成や細胞呼吸が不可能な環境に置かれた。呼吸ができるくらいに十分なO2が大気を満たすまでは、植物による何十億年にわたる光合成が前提になければならなかった。
光合成で放出されたO2を活用して、初めて、植物のミトコンドリア・マトリックス内のクエン酸回路で8個の反応を行うことができる。
動物・植物・藻類・細菌が腐植・分解した堆積物が、化石燃料として人により燃やされCO2の形で大気中に戻されていく。
目次へ