| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生
| 目次 |
| 1)胚発生 |
| 2)栄養発生 |
| 3)生殖発生 |
| 4)維管束形成層vascular cambium |
| 5)花成(生殖発生) |
1)胚発生
胚発生embryogenesis(èmbriɑ'dʒəni)
 ほぼすべての陸上植物にとって、必須の条件は、独立栄養の生活様式を備えることである。好ましい環境で生育する植物は、光合成のおかげで、成長し生存を全うするに必要なエネルギーと栄養を容易に得ることができる。移動する必要のない植物は、動物のように移動するために必要な解剖学的に複雑な構造に進化させる必要はなかった。植物は、光合成を行うために、特に太陽光などの条件がよくない場所や、時間とともに条件が変わる場所でも、固着して生活することが可能な融通の利く柔軟な成長パターンを内在させている。
ほぼすべての陸上植物にとって、必須の条件は、独立栄養の生活様式を備えることである。好ましい環境で生育する植物は、光合成のおかげで、成長し生存を全うするに必要なエネルギーと栄養を容易に得ることができる。移動する必要のない植物は、動物のように移動するために必要な解剖学的に複雑な構造に進化させる必要はなかった。植物は、光合成を行うために、特に太陽光などの条件がよくない場所や、時間とともに条件が変わる場所でも、固着して生活することが可能な融通の利く柔軟な成長パターンを内在させている。従属栄養生物である動物は、固着ではなく移動機能を進化させてきた。代わりに植物は、容易に光エネルギーと栄養が得られるように適応した、比較的厳密に定まった構造が形成されてきた。植物細胞は動物細胞とは異なり、比較的硬い木質性の組織の中で隣の細胞と密着している。この堅固な解剖学的構造によって、植物の成長様式は多くの制約を受ける。分裂組織という限られた場所に置かれた組織構造の働きによって、新たな細胞は、植物体に順々に積み重ねられていく。一方、動物の発生では、基本組織層などの形成などの多くの発生過程では、細胞が新たな別の場所に移動する。
植物は独立栄養の生活様式であるため、比較的単純な体制を持つことになった。また移動しない生活様式は、植物に大きな課題を背負わしている。植物は最適な環境領域を選んで移動できないので、自身が置かれた環境を受容しなければならない。こうした適応は生理学的なレベルで起こるが、栄養成長の特徴である柔軟な発生パターンの調節を介して起こることもある。この適応的成長の鍵は、未分化な細胞の供給源を含む分裂組織が存在することである。これらの細胞の増殖や分化を制御することによって、植物は環境に合わせて、様々で複雑な形態を作り出すことができる。
種子植物の胞子体の発生は、3つの主な段階に分けられる。胚発生、栄養発生、生殖発生である。
胚発生とは、単細胞の受精卵から、ある特徴を持ちながらも、基本的体制を維持した一つの多細胞体に変化する過程を表す。ほとんどの種子植物の胚発生は、花の雌ずいの中の、胚珠と呼ばれる特殊な構造の内部で起こる。胚発生の過程は、かなり予測できるが、これは胚が母親の珠皮の中にきちんと包み込まれていることが、種子の形成のために必要なことを反映している。珠皮は、種子植物の胚珠の外側にあって珠心を包む組織で、1層のものと内外2層からなるものとがあり、胚珠が種子になると種皮になる。胚発生には、植物の基本パターン形成過程の中でも、特に基本的体制が明確になる過程が含まれている。
 胚発生時の単細胞の接合子は、シュートと根の頂端分裂組織に含まれる未分化細胞集団を特徴とする、未発達ながらも極性を持った体制を確立していく。栄養発生の間、無限の成長パターンが、可変的なシュート系と根系を構成する。それは固有の内生プログラムと環境要因からの入力を反映する。胚の発生は、胚における位置に応じて、どの細胞が分化するかを決める枠組みを与える。この枠組みの中で、複数の細胞群が表皮や皮層、維管束組織などに、機能的に組織化する。
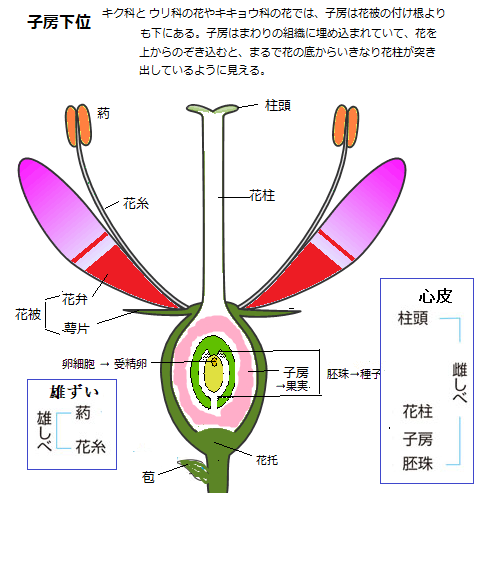
胚発生時の単細胞の接合子は、シュートと根の頂端分裂組織に含まれる未分化細胞集団を特徴とする、未発達ながらも極性を持った体制を確立していく。栄養発生の間、無限の成長パターンが、可変的なシュート系と根系を構成する。それは固有の内生プログラムと環境要因からの入力を反映する。胚の発生は、胚における位置に応じて、どの細胞が分化するかを決める枠組みを与える。この枠組みの中で、複数の細胞群が表皮や皮層、維管束組織などに、機能的に組織化する。頂端分裂組織として知られる細胞群は、シュートと根のそれぞれの成長点に確立し、胚発生以降の栄養成長では、組織や器官を次々と生み出していくことができるようになる。生殖発生の間に、栄養成長のシュート頂分裂組織は、再プログラム化され、心皮carpel(kɑ'ːrpəl)と雄ずいを含む特徴的な一連の花器官を生み出してから、半数体の配偶体世代が始まる。種子植物の花は、一つの枝の各葉が子房・花柱・柱頭と特殊な分化をして、生殖器官となったものとみることができる。このめしべを構成する雌性配偶体、即ち雌ずいを形成する葉を心皮という。シダ植物では大胞子葉がこれに相当する。
雄ずいstamen(stéɪmən)は、種子植物の生殖器官・雄性配偶子で、花粉を含む葯と、それを支える花糸からなる。シダ植物の小胞子葉、裸子植物では花粉嚢のついた鱗片状のものが多く、雄ずいでなく小胞子葉ということが多いが、これが集まったものを雄花とも言う。
胚発生が終わると、多くの生理学的な変化が起こり、胚は長い休眠に入り、過酷な環境条件を耐え凌ぐことができるようになる。
目次へ
発芽する際、胚は休眠状態から目覚め、貯蔵していた物質などを利用して栄養成長を開始する。発芽のタイミングは、植物種にもよるが、季節・湿度・長期の低温や高温(熱)・光といった様々な因子の組み合わせで決まる。マメ科植物の子葉やイネ科植物も胚乳などの貯蔵物質を、はじめに利用することで、芽生えは根端とシュート頂の分裂組織の働きを通して、その基本的形を作り上げる。
エンドウ・ダイズ・ソラマメ・ピーナッツなど豆類の種子は、2つに分類できる。いずれも、栄養分を十分に貯えた子葉であるが、発芽する際、エンドウとソラマメの子葉は地中に残ったまま、地上にでる茎葉部に栄養を供給し続ける。ダイズとインゲンの子葉は、短い茎の上に乗って地上に現れるが、葉のように展開したり緑色を濃くしたりすることはない。子葉の間から出てくる茎葉部などに栄養を供給し続け、栄養分がなくなれば萎れて落ちて役割を終える。いずれも、次世代の植物本体が、独力で光合成を行って自立成長できるまでの、栄養分を補給している。
地上に出て緑色となる大豆の子葉は、かなり厚い。そのため、マメ科の子葉は殆ど光合成に寄与できないようだ。マメ科の子葉は、地上にあっても地下にあっても、基本的に貯蔵器官としてだけ働くと考えられている。それを養分として形態形成されるシュートの芽生えにより、光合成ができるようになり、その結果、栄養成長が進められる。
動物の成長とは異なり、通常、植物の栄養成長はエンドレスである。実は、植物の成長過程は前もって決まっていないため、明確な終着点がない。環境変化に応じる、無限とも言える成長パターンがあり、このバリエーションに、側方器官を繰り返し発生させるプログラムが用意されており、植物は局所的な環境に、最も適した体制の構築を積み重ねて適応しようとする。
 植物の分裂組織の機能は、幹細胞stem cell(分裂して自分と同じ細胞を作るself-renewal能力がある。)の維持と、側方器官のもととなる創始細胞founder cellの特異化の両方を必要とする。根の成長の多くの特徴は、過酷な環境変化に適応する試練を経て形成されてきた。
植物の分裂組織の機能は、幹細胞stem cell(分裂して自分と同じ細胞を作るself-renewal能力がある。)の維持と、側方器官のもととなる創始細胞founder cellの特異化の両方を必要とする。根の成長の多くの特徴は、過酷な環境変化に適応する試練を経て形成されてきた。根は、植物を固着させ、土壌から水とカルシウム・ナトリウム・カリウム・マグネシウム・燐・硫黄・鉄などの無機塩類などの栄養を吸収してきている。しかも、障害となる異質な環境を探索し、それを利用できるように、複雑な成長パターンや屈性パターンを示す。
根端分裂組織で作り出された細胞は、根の頂端から離れていくにつれ分裂し、伸長し分化する。その分化はシュート頂の場合と似ている。しかし、根毛や側根のような根の側方への成長は、細胞伸長が終わった根端から離れた領域で起こる。このように根端とは離れた部位に側方器官を形成することにより、それらが土壌中の剪断力でダメージを受けないようにしているだけでなく、根端が一組の始原細胞群を維持し、それらの分裂活性を制御することに集中できるようになる。
始原細胞initiating cellは、根やシュートの分裂組織で、ゆっくりと分裂する未分化の細胞集団で、それぞれの子孫細胞は、極性パターンを持った細胞分裂により遠方へ押しやられる。
それぞれの分化の運命に従い、根やシュートの放射軸(太さの方向)や長軸の形成や側生器官の発生に寄与する。
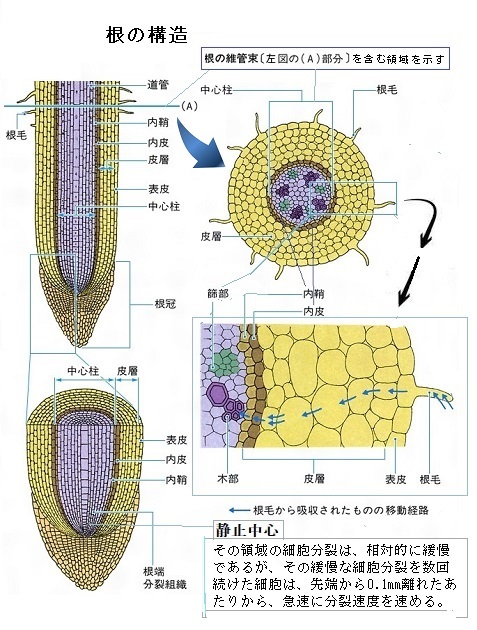
根の放射軸radial axisは、根の外側から中心に向かう軸で、根の横断面を観察してみると、各組織の細胞が表皮・皮層・内皮・内鞘(ないしょう)・維管束などが放射軸に沿って、同心円の層状に存在する。これらは胚の発生時に確定する。
内鞘は、高等植物の茎または根の内皮のすぐ内側にあり、維管束を囲む柔組織の細胞層である。根では、成熟部位の内鞘が分裂して生じるのが側根lateral rootで、皮層や表皮を介して成長する二次分裂組織の形成や、新たな成長軸の形成をする。Lateral(lˈæṭərəl)は、側面の、側生の意味である。
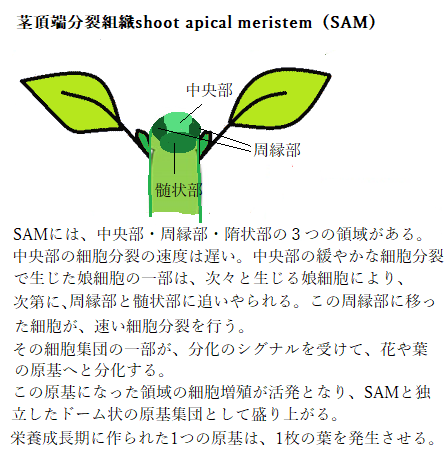
 成長点は分裂組織とも呼ばれる分裂能力の高い細胞の集団で、茎と根の先端にあって茎と葉や根の組織を作る。
成長点は分裂組織とも呼ばれる分裂能力の高い細胞の集団で、茎と根の先端にあって茎と葉や根の組織を作る。植物が成長を続ける間は、茎や根が伸び、次々に葉や花が形成される。これは成長点の細胞が、分裂して茎や葉、根の細胞に変化していくからである(細胞分化)。
ところが、分化した細胞は分裂能力が低下することが知られている。一方、成長点にあって盛んに分裂する細胞は分化していない。そのため、成長点の細胞が分裂して生まれた娘細胞が分化せずに分裂を繰り返すならば、巨大な成長点ができるだけで、新たな組織は形成されない。
植物が成長を続けることができるのは、成長点で生まれる娘細胞には、さらに分化し、他の組織を作る細胞と、分化せず高い分裂能力を保つ細胞の数が、常に一定の比率を保っているためと考えられている。
シロイヌナズナの根の先端にある成長点にある根端分裂組織の中心には、4個の静止中心と名付けられた細胞が集まっており、この細胞群を取り囲んで数十個の始原細胞と呼ばれる細胞がある。静止中心細胞は殆ど分裂しないが、始原細胞は活発に分裂する。各々の始原細胞は一方の側で静止中心細胞と接しているので、分裂によって生まれた2個の娘細胞のうち、一つは静止中心細胞と接し、他方は離れている。静止中心細胞に接している方の娘細胞は分化せず分裂を繰り返し、静止中心細胞と接していない方の娘細胞は分化して根の組織を作ることが知られている。
つまり、分化を抑制し分裂を促進するシグナルが、静止中心細胞から始原細胞に送られ、このシグナルは隣合った細胞には伝わるが、離れた細胞には伝わってはいないようだ。このシグナルを受け取った細胞は分裂するが分化せず、シグナルを受け取らなかった細胞は分化するが分裂しないことになる。このような仕組みで、娘細胞のうち一方は、親細胞と同様な分裂する性質を持ち、始原細胞としての役割を受け継ぐけれども、他方はこの性質を保つことができず分化すると考えられる。ただし、静止中心細胞から送られるシグナルの本体や、始原細胞がシグナルを受け取る仕組みは未だ分かっていない。
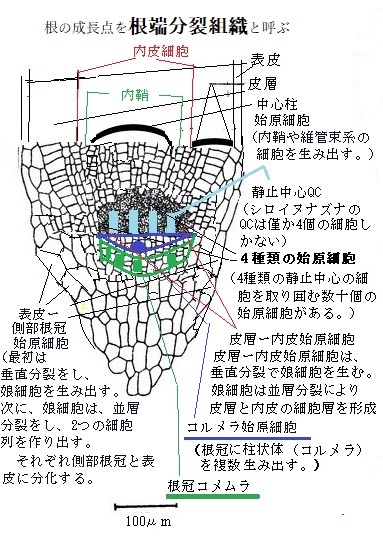 根の異なる組織は、その始まりを特定の始原細胞に遡ることができる。根の組織は、進行的かつ直線的に発生するので、それぞれの始まりが、根の先端部にある特定の始原細胞群にあると予想できる。大部分の植物の根では、正中線を通る縦断面を見ると、縦長の細胞列は、根のやや頂端部分に近づくと、次第に集まって収束する。この収束した部分が、静止中心quiescent(kwɑɪésnt)
center QCで、その名の由来は、根の分裂組織の中心部位でありながら、周りの細胞に比べて細胞分裂の頻度が比較的低いか、あるいは全く分裂しない、ことによる。
根の異なる組織は、その始まりを特定の始原細胞に遡ることができる。根の組織は、進行的かつ直線的に発生するので、それぞれの始まりが、根の先端部にある特定の始原細胞群にあると予想できる。大部分の植物の根では、正中線を通る縦断面を見ると、縦長の細胞列は、根のやや頂端部分に近づくと、次第に集まって収束する。この収束した部分が、静止中心quiescent(kwɑɪésnt)
center QCで、その名の由来は、根の分裂組織の中心部位でありながら、周りの細胞に比べて細胞分裂の頻度が比較的低いか、あるいは全く分裂しない、ことによる。様々な組織を生み出す始原細胞と、その隣のQC細胞とが近接しているのは、両者の細胞間で強い機能的な相互依存関係があることを示唆している。多くの陸上の根では、QC細胞が、時折分裂して隣接する始原細胞と入れ替わることがある。
胚発生の細胞分裂パターンのように、QC細胞と周囲の始原細胞の振る舞いは、植物種の間で異なる。この事から、位置依存的な機構が、これらの細胞タイプを規定するのに重要な役割を果たしていると想定される。
シロイヌナズナの根のすべての組織は、根端分裂組織の少数の始原細胞に由来する。シロイヌナズナのQCは、僅か4個の細胞から成り立っている。これらの細胞は、胚発生以降殆ど分裂しない。その4個のQC細胞を取り囲む、4種類、数十個の始原細胞は、どれもQC細胞に隣接している。
静止中心細胞は、殆ど分裂しないが、始原細胞は活発に分裂する。各々の始原細胞は、一方の側で静止中心細胞と接しているので、分裂によって生まれた2個の娘細胞のうち、一つは静止中心細胞と接しているが、他方は離れることになる。
QC細胞の直下、根冠の頂端側、その中心部の円柱状領域にあるコルメラcolumellaは、その直下の根冠の中心部にコルメラ始原細胞群を生み出す。分化して根冠の中央部を形成する。根の重力屈性では、そのコルメラ細胞が重力を検知している。
QC細胞の横側に位置している表皮―側部根冠始原細胞は、まず垂層分裂をして娘細胞を生み出す。娘細胞は、並層を行い2つの細胞列を作り、それらが側部根冠と表皮に分化する。
表皮―側部根冠始原細胞の内側に接している皮層―内皮始原細胞が、垂層分裂により娘細胞を生み出す。その娘細胞が並層分裂をし、皮層と内皮の細胞層を形成する。
QC細胞の上部に隣接(根の基部側)する中心柱始原細胞は、内鞘を含む維管束系を生み出す。
目次へ
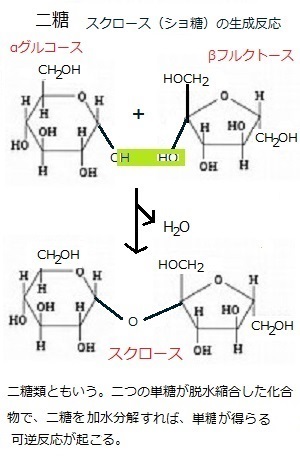
生殖とは、生物が、自己と同じ種類の新しい個体をつくる過程を言う。すべての生物にみられる基本的特徴で、これによって種族の維持がなされる。
一つの相から別の相への移行を、相転換phase transitionという。栄養成長の期間が終わると、植物は植物体内外からの様々な情報、植物体のサイズ・温度・光周期など、様々な組み合わせに応答して、生殖発生へと成長相を移行させる。顕花植物では、この相転換に、花を作るために特化した花芽分裂組織が特定化され、その分化が決定した後、型にはまった器官形成を行う。
春から夏、夏から秋、秋から冬への季節の移ろいとともに、野生植物は、それぞれ定められた時期に花を咲かせる。それは、植物の繁殖適応にとって極めて重要である。他家受粉を行う植物は、一年の内の結実に最も適した時期に、同じ種の受粉相手の開花時期に合わせて、送粉者に同調して花を咲かせなければならない。植物の生活環の中で最も重要な発生上の決断は、いつ花を咲かせるかである。シュート頂分裂組織が、花の形成へと決定づけを遅らせることで、転流translocationに回すことができる炭水化物の蓄えを増やし、成熟過程で種子により多くの貯蔵栄養分を蓄えることができ、より多くの種子を作ることができからである。
植物細胞内部は、わずか脂質2分子分の厚さしかない細胞膜で、細胞壁や周りの環境から隔てられている。この極めて薄い膜の層により、激しく様々に変化する外部環境に対して、比較的安定した内部環境が維持される。細胞膜は、物質の拡散に対して疎水的バリアを形成するだけでなく、細胞が必要とする栄養の取り込み、細胞内物質の排出、あるいは膨圧の調節などのため、特定の物質やイオンの内向き、あるいは外向きの輸送を維持して、常に制御しなければならない。
同じような働きが、細胞各々の中の様々な区画compartmentを仕切る内膜系にも当てはまる。細胞膜は、周りの物理環境、他の細胞からの分子情報、攻撃的な病原体に関する情報を伝える役割を担っている。これらの情報は、しばしば膜を介したイオン輸送の変化として伝えられる。
ある場所から他の場所への分子やイオンの移動は、輸送transportとして理解されている。植物体において光合成産物や栄養塩類などの分子やイオンが、ある器官・組織から他の器官・組織に輸送される。細胞外から細胞内への、あるいは細胞内での居所的な輸送は、多くはタンパク質によって調節されている。近傍の細胞間の物質は、主に原形質連絡を経由した輸送が行われる。
植物器官の間や、植物と環境の間で行われる大量の輸送も、細胞レベルでは膜によって調節されている。例えば、葉から根への篩管を経由するスクロースの輸送、これが転流translocationであるが、葉における篩管細胞内への膜輸送系によって担われ、調節を受けて、篩管から根の貯蔵細胞へと輸送される。生細胞における輸送の分子機構と、植物細胞において特徴的な輸送系を担う様々な膜輸送タンパク質が働いている。
陸上植物が、水が不足がちな陸上環境に適応するために、植物は根と葉を進化させた。根は植物を固定し、水と栄養を吸収する。葉は光エネルギーを得てガス交換する。植物が大型化するにつれ、根と葉の距離は遠ざかる。そのため根で吸収した水やミネラルを葉に送り、葉で合成した光合成産物やエネルギーを根に送る効率的な長距離輸送システムを進化させた。隣接する、もしくは長距離輸送する器官間の物質輸送は、通常は通導組織を経由する。篩部輸送と木部輸送とに大別され、光合成産物の転流はもっぱら前者で行われるが、栄養塩類の転流は両者で行われる。
篩部は複合組織であり、植物全体へ糖や他の有機化合物の輸送を担う細胞全体を篩要素sieve element(sív;ふるい、こしき)と呼ぶ。それは、複数の異なる形態を含む幅広い細胞を含み、典型的な被子植物に見られる高度に分化した篩管要素sieve tube elementや、裸子植物の比較的特化していない篩細胞sieve cellをも含めて篩要素と言う。篩要素に加えて、伴細胞companion cellと柔細胞parenchyma cellがある。
篩要素は、成熟すると核をふくむ原形質のほとんどを失い、互いに連なって篩管sieve tubeを形成し、転流物質の流路として機能する。篩管の両端部分の細胞壁には篩孔sieve pore(pˈɔɚ)と呼ぶ多数の小孔をもった篩板(しばん)sieve plateがあって、孔の開いた仕切りの細胞壁は、篩い(ふるい)状になっており、物質が透過できる様になり、隣接する篩管同士と連絡している。 つまり、細長い篩管細胞が縦方向につながり、細胞の中身がつながった形をしている。
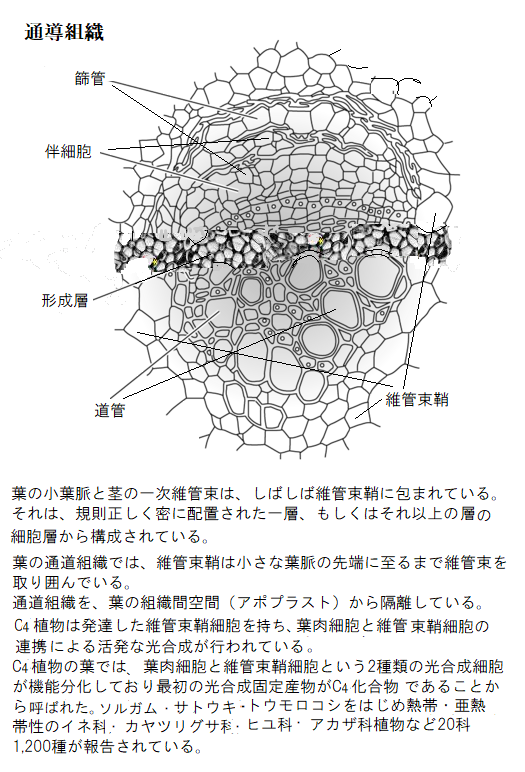 篩管要素には、1つもしくは複数の伴細胞が隣接している。実は、単一の母細胞の細胞分裂により、篩管要素と伴細胞に分化する。裸子植物には、伴細胞がない代わりにタンパク細胞albuminous
cell(ælbjúːmənəsタンパク質を含むの意味)というものがあって伴細胞に似た働きをしていると考えられている。
篩管要素には、1つもしくは複数の伴細胞が隣接している。実は、単一の母細胞の細胞分裂により、篩管要素と伴細胞に分化する。裸子植物には、伴細胞がない代わりにタンパク細胞albuminous
cell(ælbjúːmənəsタンパク質を含むの意味)というものがあって伴細胞に似た働きをしていると考えられている。篩管細胞は、核を含む原形質の殆どを失っている。それでも10年、100年と生きている。細胞が生きて行くのには、核の遺伝子の遺伝情報を用いて作られたタンパク質が不可欠、核のない篩管細胞では、このタンパク質を作れない。必要なタンパク質は、隣接する伴細胞で作られて、篩管細胞に送られているようだ。
篩管細胞と伴細胞には、たくさんの小さな孔が細胞壁を貫いている。多くの場合、伴細胞の方が、複雑に枝分かれしている構造になっている。この原形質連絡の多数の繋がりは、この2つの細胞の機能的関係を物語っている。
伴細胞で作られたタンパク質はこの原形質連絡を通って、タンパク質を作れない篩管細胞に運ばれている。さらに、成熟葉の光合成産物を合成する葉肉細胞(ようにくー)から、その葉の小維管束の篩要素への光合成産物の輸送に、重要な役割を果たしている。葉の葉肉細胞で合成された光合成産物は直接篩管の中には入れず、先ず伴細胞に入り、そこから、これも原形質連絡を通って篩管の中に送り込まれている。
篩管細胞が必要とするタンパク質合成など、様々な生合成能力を失った代謝機能を、伴細胞が、肩代りしていると考えられている。さらに、伴細胞の多くのミトコンドリアが、篩要素へATPとしてのエネルギーを供給しているようだ。
柔細胞は生きている細胞で、原形質を有している。有機養分の貯蔵組織や分泌組織などを主に構成し、養分を合成・分解・貯蔵し放出する。細胞壁は一次細胞壁から成り、薄いのが一般的である。篩部柔細胞は、二次篩部の場合には二次木部と同様、縦に長い細胞からなる紡錘形柔組織と等径的な細胞からなる放射組織である。
輸送細胞transfer cellは、伴細胞に似ているが、指のような突起がある細胞壁を複数持ち、細胞内部に向けて肥厚する点が異なっている。この肥厚は、篩要素と接しない部位の細胞壁で、特に目立つことから、細胞膜の表面積を増大させ、アポプラストを経由して膜を横切る溶質が、細胞膜を通過できる輸送能を高めていると見られる。
輸送細胞と篩要素の間には、原形質連絡があるが、輸送細胞と周辺の篩要素以外の細胞の間には、原形質連絡は稀にしか見られない。篩要素と輸送細胞のシンプラストは、周辺細胞のシンプラストとは完全ではないが、ほぼ隔離状態にある。
輸送細胞は、篩部転流径路上にある節、ソース組織の篩部、篩要素以降の積み下ろし径路でよく見られる。
木部柔細胞xylem(zάɪləm) parenchyma cellは、輸送細胞に変化することがある。アポプラストの一部である木部を移行する溶質を吸収し、その輸送先を変えるためと考えられている。
目次へ
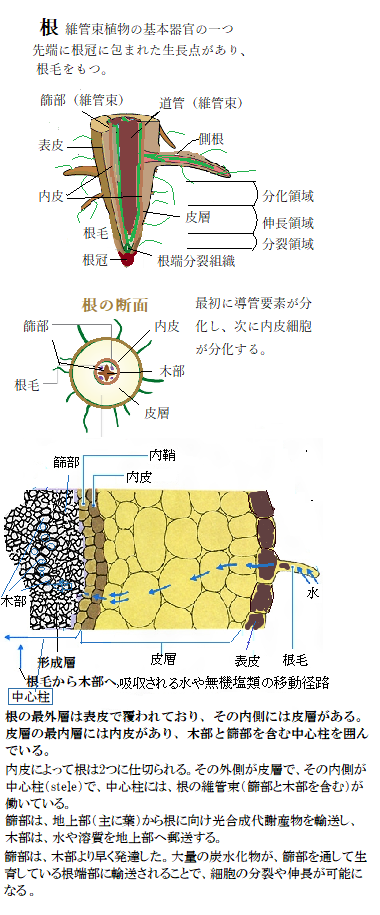 植物の一生は、根・茎・葉・花などの器官を発生させ続けながら成長する。多くの種子植物の体づくりは、器官原基の発生と形成を司る一次成長から、発生した器官の体軸肥厚を司る二次成長secondary
growthへと繋がる。一次成長は頂端分裂組織に由来する細胞が分裂・分化を経て器官形成しながら主に基部から先端軸方向へ伸長する成長過程である。
植物の一生は、根・茎・葉・花などの器官を発生させ続けながら成長する。多くの種子植物の体づくりは、器官原基の発生と形成を司る一次成長から、発生した器官の体軸肥厚を司る二次成長secondary
growthへと繋がる。一次成長は頂端分裂組織に由来する細胞が分裂・分化を経て器官形成しながら主に基部から先端軸方向へ伸長する成長過程である。一方、単子葉を除く種子植物の主に根、茎などの軸性器官で観察される二次成長は、これらの器官の中に存在する前形成層procambiumと内鞘細胞が分裂活性を獲得して、体軸が中央から周辺軸方向へ肥厚する成長過程をたどる。維管束を形成し、やがて木部や篩部が分化するが、根では内皮より内側の基本組織も前形成層から生じる。
植物の放射軸に沿った、内側から外側への方向性を持った成長を二次成長と呼ぶ。形成層cambium(kǽmbiəm)と言う分裂組織が、その役割を担っている。茎や根の太さが増す成長を司る分裂組織を維管束形成層という。単に形成層と言う方が多い。
木質部分を作り出す維管束形成層は、一次成長により作られた植物体の木部と篩部の間にある1層の維管束内形成層intrafascicular cambiumと維管束内形成層同士をつなぐ維管束間形成層interfascicular cambiumからなる。裸子植物や双子葉植物では両者ともが発達して輪状の維管束形成層となるものが多い。裸子植物や双子葉植物では維管束形成層はふつうに見られるが、キンポウゲ属のようにこれを全く欠くものもある。単子葉植物では維管束内形成層はしばしば見られるが、維管束間形成層は発達しない。
維管束形成層は一度分化した組織から生じるとされるため、シュート頂や根端のように最初から分裂能を保持している一次分裂組織に対して二次分裂組織とよばれる。しかし実際には一次分裂組織から派生した前形成層procambiumの一部にあたる、木部と篩部にはさまれた部分が、分裂能を保持したまま維管束間形成層になるため、維管束形成層は本来の意味での二次分裂組織とは言えない。維管束間形成層は、維管束内形成層が形成された後にその間をつなぐように形成され、維管束形成層は全体で環状を呈するようになる。
維管束形成層の細胞は、伸長軸に平行して分裂を繰り返し、茎や根の外側、または内側に向かって細胞を供給する。その一方、放射軸に平行しても分裂し、そのため外側に向かって放射状に、物質を輸送するための放射状柔組織rayができる。
維管束形成層の分裂により生じる細胞のうち、ふつう内側にあるものは、二次木部を、外側に位置するものは、二次篩部を作り出していく。その内側の細胞は、水や養分を土壌から植物体の様々な器官へ輸送するための二次木部secondary xylem(záiləm)へと分化する。温帯域では、秋から冬にかけて形成される目幅が狭く色の濃い部分の晩材と、春から夏にかけて形成される目幅の大きな淡色の部分の早材が、交互に積み重なり年輪が形成される。早材は木細胞が大きく粗いため材質も粗く、晩材は木細胞が小さく密度が高いため材質は硬い。
維管束形成層に由来する細胞のうち、外側へ移動した細胞は、二次篩部secondary phloem(flóuem)に分化する。二次篩部は、一次篩部と同様に、光合成産物を葉から他の器官へ輸送する。二次篩部には、篩部線維phloem fiberが付随し、これが他の繊維と同様に茎に強度を与えている。 維管束形成層は、自身が作り出した二次維管束による二次木部の成長により、次第に外方へ移動することになる。それにともなって維管束形成層は細胞数が増加し、その周長が広がる。
茎の一次成長から二次成長への移行は、その維管束形成層が、最初は一次木部と二次木部からなる維管束の中の成長域である維管束内形成層で分断されているが、成長するに従い維管束間形成層により連結される。それにより維管束形成層が環状につながると、内側方向に分裂した細胞が二次木部を形成して成長し、維管束形成層の環状の外側で分裂した細胞が二次篩部を形成する。篩部繊維やコルク形成層を含む周皮periderm組織、及びコルク皮層は、いずれも皮層から生じる。成長に伴い表皮は破断し、放射状柔組織が内側と外側の維管束を繋ぐ。
コルク形成層は、木本の表皮の内側に作られる頑丈な周皮を形成する形成層で、多くの場合、二次篩部の内部に毎年生じる。一次成長により作られた茎や根の外層の組織は、その内側にできる防水性のコルク層に阻まれて、木部からの水の供給が止まるため、乾燥により壊死する。
根の一次成長から二次成長への移行は、根の維管束とそれによって囲まれた髄を含む中心柱protostele(ちゅうしんちゅう)は、一次篩部と一次木部を含む。中心柱とは、皮層の最内層である内皮よりも内側の部分をいう。内皮は根の中心に環状に存在する細胞群を形成する。内皮によって、根は2つに仕切られる、外側が皮層、内側が根の維管束がある中心柱である。内皮細胞層の壁は肥厚し、壁の細胞に面した側から、外側に向かってスベリンsuberinを分泌し、カスパリー線Casparian stripを形成する。スベリンは、植物のコルク組織の主成分で、その疎水性の重合体は、化学的にも生物学的にも、植物の表皮のクチクラ組織にあるクチンと類似している。根の内皮細胞壁のカスパリー線は、中心柱と皮層の間に疎水的なバリアとして働くため、水や溶質のアポプラスト経由の移動がここで遮られる。
カスパリー線には、細胞壁中の限定した領域で重合したリグニンを含んでいる。 種子植物の茎では、殆ど内皮が認められないが、この場合でも境界をとくに限定しないで中心柱を想定する。維管束のすぐ外側の柔細胞が、特にデンプン粒を多量に貯めているものがあり、このような構造をデンプン鞘starch sheathという。これを、内皮細胞層のデンプン鞘と呼ぶ。 茎の場合と同様、維管束形成層は、成長に伴って連結され、外側へと成長して二次篩部と放射状柔組織を形成する。根が太くなるに従い、内鞘から根の周皮組織が生じ、その一方で、表皮・皮層・内皮が剥離する。篩部繊維・放射状柔組織及び側根は、いずれも内鞘から生じる。維管束形成層は二次篩部と管状の二次木部を形成する。
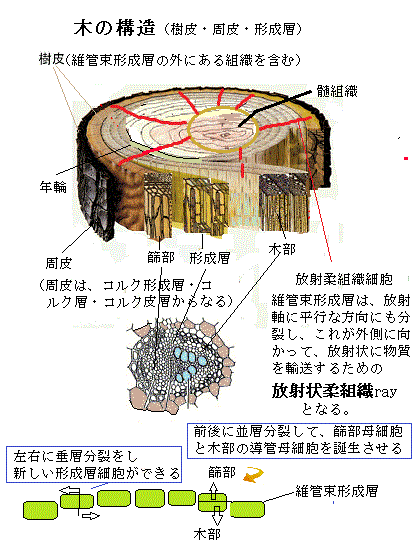 樹皮という語は植物学用語ではないが、しばしば周皮にのみを指すと誤解されている。一般的には、維管束形成層の外側にあるすべての組織を含む。機能を持つ二次篩部・つぶれて機能を失った二次篩部・つぶれた一次篩部とそれから周皮からなる。
樹皮という語は植物学用語ではないが、しばしば周皮にのみを指すと誤解されている。一般的には、維管束形成層の外側にあるすべての組織を含む。機能を持つ二次篩部・つぶれて機能を失った二次篩部・つぶれた一次篩部とそれから周皮からなる。木本で言う樹皮barkは、通導組織として二次篩部を主体とし、二次篩部繊維・皮層(茎のみ)・内鞘(根のみ)及び周皮(コルク形成層・コルク層・コルク皮層)と言った複数の組織を持つ構造である。維管束形成層は、細胞分裂中の組織を含んでいるので、その内外のどちら側の二次組織よりも脆いため、特に樹皮は樹から剥離しやすい。
通導組織として機能を持つ二次篩部を主体とする部分を内樹皮、その外側にありつぶれて機能を失った二次篩部・つぶれた一次篩部、さらにその外側に形成されるコルク組織(周皮)をまとめて外樹皮と呼んでいる。
木本性の茎の一次成長は、春に起こり、引き続き二次成長へ移る。太くなる方向への成長は、二次維管束組織を生み出す二次分裂組織の、維管束形成層とコルク形成層によってなされる。
維管束形成層の領域は、1層の形成層幹細胞からなる。その内外領域に、その形成層幹細胞に直接由来した組織ができる。外側が二次篩部細胞で、内側が二次木部細胞である。
周皮は、コルク形成層とコルク細胞からなる。
陸上植物の細胞壁は一次細胞壁と二次細胞壁に大きく二分される。すべての植物細胞が持つ一次細胞壁は、セルロース・キシログルカンなどのヘミセルロースとペクチンの3 つのグループからなる多糖類と構造タンパク質から構成され、植物細胞の形や生理学的機能を制御している。
これに対して二次細胞壁は、一次細胞壁の内側に形成される肥厚した細胞壁で、セルロースや、主にキシランなどのヘミセルロース、そしてリグニンを主要な構成成分とし、維管束組織や表皮組織などの一部の組織と細胞で特定の発生段階で形成される。リグニン化により、幹の重量を支える構造的な役割を担う。
形成層での細胞分裂面の方向は、直径方向の成長と周長方向への成長のバランスが保たれながらなされる。形成層細胞は、初めは垂層分裂anticlinal division(茎の表面に対して垂直な分裂)によって新しい形成層の始原細胞を生み、形成層の周長を増やす。その同じ始原細胞は並層periclinal division(茎の表面に平行な分裂)をして導管母細胞と篩管母細胞を作り出しつつ、その背後に、常に始原細胞を残してい。
垂層分裂は、形成層に細胞を供給し、茎の周長を伸ばす方向に働くから、それなくして形態学的に形成層の他の細胞と区別がし難いため、形成層始原細胞の位置を示す指標となる。最も活発な垂層分裂は、篩部よりも手前側1番目あるいは2番目の細胞層で専ら観察されるので、これにより維管束形成層のおおよその位置が知られる。
典型的な両面性の形成層では、並層分裂は、木化した茎の外側に篩部を、内側に導管を生み出す。木部の増殖は、導管要素細胞の、数日間という寿命の間、継続されるので、他とは不釣り合いとなるほど活発で、いろいろと複雑に見える。
篩部組織と導管組織に加えて、維管束形成層は放射柔組織細胞という、茎の中で側方への物質輸送や、冬期の休眠時のような不適な環境にある際に、物質を貯蔵する際の通り道になる柔組織を作る。放射柔組織細胞は、単列で、あるいは複数列で、篩部・形成層・木部を貫いて並ぶ放射組織を形成する。
殆どの被子植物と裸子植物の木本性の種類は、放射柔組織始原細胞が放射線状に並んだ細胞系譜を持っていて、これが栄養の輸送や貯蔵の役目を果たす。
殆どの木本性の真正双子葉類(真正双子葉類は、被子植物の単系統群の一つで、従来の双子葉類の中から、単子葉類が分岐する前に分岐した古いグループのモクレン・センリョウ・スイレン・シキミ・アムボレラなどを除いたもの。)と裸子植物は、周皮を作り出すコルク形成層と呼ぶ二次形成層を発達させた。コルク層は、スベリン化した細胞壁を持つ死細胞からなる多層の保護組織である。そのコルク層の内側に生じたコルク皮層は、生きた柔組織である。
通導組織として機能しなくなった篩部、さらにその外側に形成されるコルク組織をまとめて外樹皮と呼んでいる。しかし、樹木が生きている限り内樹皮に、コルク形成層は常に存在している。
樹皮の形成層から新たにできた二次篩部の経時的形態変化は、形成層からは篩細胞が数層形成されると一層の柔細胞が形成されるという過程を繰り返している。この繰り返しが数層になると一番外側の柔細胞が活性化、不規則な形状となり、その一部は厚膜細胞に転化する。形成層で篩細胞や柔細胞が新生されれば、外側にある古い篩部、柔細胞は押し出されて不規則な配列、柔細胞のさらなる分裂、厚膜細胞の形成を経て、最外層の外樹皮へと移っていくことになる。形成層から篩部細胞とともにつくられた柔細胞の活発な分裂活動で複雑な構造の内部樹皮が形成される。ついには新たに形成されたコルク形成層によって外樹皮へと転換していく。
コルク形成層は通常一層の細胞層で、外側にコルク細胞を、内側にはコルク皮層を形成し、コルク形成層は、茎肥大成長にともなって崩壊すると新たに二次コルク形成層が発達し、コルク細胞、コルク皮層の形成を繰り返えす。しかし、最初に形成された一次コルク形成層が、何回ほどで、コルク細胞やコルク皮層を作り続けて崩壊し、二次コルク形成層となるかについては、コルク層の厚さを生み出す活性の程度が、木の種によって大きく異なり、寿命も種によって大きく異なるため明らかになっていない。
樹高 18mにもなる、ブナ科の常緑高木で、スペインを中心とする地中海沿岸地域のコルクガシは、永続的に活動するコルク形成層を持ち、樹齢 20年に達するとこのコルク層をはぎ取っても、樹木の生育は阻害されず、新しいコルク層を再生する。その後9年~12年ごとにほぼ 150年ぐらいにわたって、厚さ4~5cmのコルクを約12回、収穫を行うことができる。世界全体のコルク生産量の約50%はポルトガルが占めている。コルクには細胞膜にスベリンが沈着し、酸やアルカリに強く、しかも適当な弾性があるため水や空気を通さない。瓶の栓として最適である。その厚いコルク層は、暑く乾いた地中海気候の環境下で、樹の主幹を乾燥から護っているようだ。
維管束形成層による肥大成長によって本来の表皮が崩れ落ちると、そのかわりに植物体の表面を保護する周皮がつくられる。周皮は表皮の内側の、細胞が分裂能を回復させたコルク形成層とそこから作られた組織からなる。
すべての裸子植物と、真正双子葉類のうち木本性の灌木や木、及び大型の草本性の殆どは、茎や根が放射成長(太さ方向への成長)する側生(⇔頂生)の分裂組織を発達させた。その側生分裂は、二次成長secondary growthと呼ぶ。二次成長にかかわる分類組織の1つが、維管束形成層であり、もう1つがコルク形成層である。そのコルク形成層が、植物体を保護する外部組織・周皮を作り出す。
維管束形成層による二次成長は、維管束植物の進化の過程で幾たびも登場している。多くの絶滅したグループにも、明らかな二次維管束組織が認められている。一方、単子葉類は、維管束形成層を持っていない。それで、二次成長が明白に確認できない。
裸子植物や真正双子葉類eudicotsによる一次成長から二次成長への切り替えは、シュート軸に沿って明らかに見ることができる。ポプラの一次成長では、先端の8節間のシュート頂分裂組織SAMから15cmほどのところまで見られる。一次成長は、その後の二次成長段階で木質化し、二次篩部と二次木部を作る。一次成長と二次成長との領域は、時間的にも空間的にも切り離されている。容易に区別が可能で、ポプラのような成長の速い種類では、急速に1~2ヵ月の内に発達する。
大木とでしか見えない単子葉植物ヤシ目ヤシ科のヤシ類ですら、維管束形成層を欠くので、その太さの増大は、葉原基leaf primordiumの直下の頂芽に局在する、特殊化した分裂組織の一次肥大分裂組織primary thickening meristemのみによってなされる。シュート頂直下の茎の直径を太くする働きをし、それにより幹は非常に高く伸びることができる。維管束を含む一次組織を付加する種類によっては、一次肥大分裂組織は、活動が永続して、柔組織や維管束をも付け加えていくことがある。
Primordium(praimɔ'ːrdiəm)は、生物学では、原基・始原体の意味、植物の幹細胞stem cell、始原細胞initiating cellなどの未分化細胞を刺激することにより植物生体を活性化し成長促進を図る。根と茎は頂端分裂組織を持つが、葉原基は茎頂端分裂組織shoot apical meristem《⇔根端分裂組織root apical meristem、mérəstèm;分裂組織》によって作られる。葉は、根や茎と同格ではなく、茎の付属物として考える。花原基は茎頂の先端に、葉原基は生長点の頂端から、ややはずれた側方にこぶ状の膨らみを付ける。
目次へ
種子植物の胚発生embryogenesisは、たった一個の細胞の受精卵(接合子)から、かなり複雑な個体を、成熟した種子の中に作り出す。その胚発生には、多くの発生過程をたどる原基が、未分化のまま含まれている。やがて、それぞれの過程をたどり、植物の基本的な形態が構築される。
その過程は、形を作る形態発生morphogenesis(mɔ̀rfədʒɛ́nəsəs)、機能的に組織化された構造を形成する器官発生organogenesis、そして細胞が分化し機能的に明確になる組織発生histogenesis(hìstədˈʒenəsis)などがある。これらの基本構造を発生させる最重要な役割を果たすのが、シュートと根にある頂端分裂組織である。これらの組織が植物の栄養成長を無限成長に近づけるほどに支えていく。
 植物は胚発生の最終段階で、胚を長期間にわたって不活性にする複雑な生理的変化・休眠dormancyと、その後に期待されている環境情報を確かに認識し成熟胚が再び成長する発芽germination(dʒ`ɚːmənéɪʃən)の過程に入る。
植物は胚発生の最終段階で、胚を長期間にわたって不活性にする複雑な生理的変化・休眠dormancyと、その後に期待されている環境情報を確かに認識し成熟胚が再び成長する発芽germination(dʒ`ɚːmənéɪʃən)の過程に入る。上記の栄養成長の期間が終わると、植物は植物体内外からの様々な情報、植物体の成長度・温度・風雪・光周期・春化などの組み合わせに応答して、生殖発生reproductive developmentへと生殖相を移行させる。顕花植物では、これらの相転換に、花を作るために分化した花芽分裂組織が関与してくる。その花成へ移行には、複数の因子と制御径路が関わる。
自律的な制御により花成する植物と、光周期に頼る植物のいずれでも、葉で作られる伝達性のシグナルが、シュート頂における花成決定に必要である。
一度、花成の惹起を遅らせることは、食害や非生物的な環境ストレスの危険を増し、繁殖前に他の植物との競争に負けてしまうリスクが大きくなる。そのため、植物は変転極まりない環境に、多岐にわたる繁殖適応能で凌ぎながら、一年生と多年生の生活環などを進化させてきた。
ノボロギクはヨーロッパ原産の1年草、明治始めに帰化し、畑や畦、道端や側溝など、野に咲く花、花期はほぼ通年である。そのノボロギクは、発芽後数週間で花を咲かせる。樹木では花を咲かせるまで20年、あるいはそれ以上かかるものもある。種によって花を咲かせる齢(plant age)は、実に多様で、個体の齢や大きさなどが、生殖成長への切り換えを調節する内生要因としてあるのだろう。
花成が、いかなる特定の環境条件とも独立し、内的な発生要因だけに応答して起こるような場合、自律的制御autonomous regulationと呼ぶ。一方、花成のためには特定のセットの環境刺激を絶対的に要求する種では、花成は絶対的な応答であるとみなされる。もし花成が、ある環境刺激environmental stimulationによって促進されるものの、そのようなシグナルがない場合でも最終的には起こる花成応答は、例えば、条件的な花成応答を持つシロイヌナズナでは、生殖成長を促進するのに、環境シグナルと自律的シグナルの両方に依っている。
光周性photoperiodismと春化vernalizationは、植物の季節応答の背景にある2つの最も重要な機構である。光周性は、昼と夜の長さに対する応答であり、春化は長期間の低温による花成の促進を指す。光質や外気温、非生物的なストレスといった他のシグナルも、植物の発生を促進する重要な外的信号刺激となる。
内的な自律的調節系と外的な環境感知的調節系の両方が進化したことにより、植物は花成を正確に調節し、繁殖が成功する最適な時期に行われることを可能にした。特定の種の多くの個体群において、花成は同調しており、それは外交配には好都合である。
環境からのシグナルに応答した花成は、殊に水と温度という観点からも、好条件下で種子が産生されている。このことは、地球環境の温暖化のような、花成の時期を支配する制御ネットワークを変えてしまうような急激な気候変動には、植物は極めて脆弱であると言える。
ひとたび花成が惹起されると、花の形成が始まる。配偶体を保護しつつ、送受粉動物を惹き付け、自家受粉ないし他家受粉を促し、多様な果実と種子を生産する。この過程における適応進化により、花の形は多様に千変万化した。その多様性にも関わらず、1つの遺伝子ネットワークが、シロイヌナズナの花ばかりか、大きく異なるキンギョソウの花などの形態形成を調節していることが明らかになった。
生殖分裂組織は、通常、栄養分裂組織よりも大きい。栄養分裂組織では、中央領域にある細胞は、その細胞分裂周期をゆっくり完結させる。栄養成長から生殖成長への相転換は、シュート頂分裂組織の中央領域における細胞分裂の頻度を著しく増加させる。それは、中央領域内の分裂組織が増大する様子が、視認できるほど膨らむ。
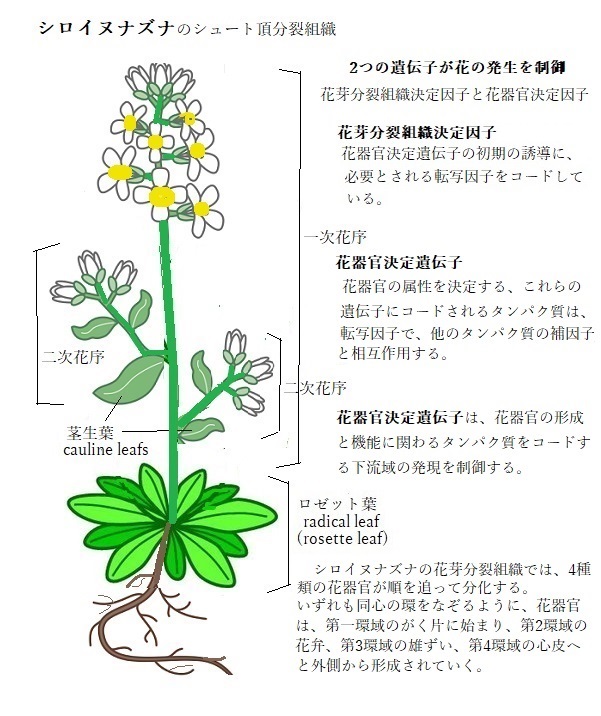
栄養成長の間、シロイヌナズナのシュート頂分裂組織は、個体の成長の各段階で異なる器官を作り出す。成長の初期には、地面に接する茎の下部に、非常に短い節間で複数の葉を伸ばし、その植物体の下部にロゼット葉(根生葉)を形成する。生殖成長の開始とともに、花成への移行が始まると、シュート頂分裂組織は、一次花序分裂組織に転換し、花を付けた長く伸びた茎を立ち上げる。その伸長した花序の軸に、2つのタイプの側生器官を形成する。茎生葉(あるいは花序葉)と花である。花成に先立ち発生した葉原基は、茎生葉に分化する。この一次花序に複数発生した茎生葉の腋には、二次花序分裂組織が形成される。その活動は一次花序分裂組織の発生パターンを繰り返す。
シロイヌナズナの花序分裂組織は、無限に成長する潜在能力がある。それ自身が何らかの器官を形成して成長を止めることがない「無限成長indeterminate growth」を示す。花は最初に、花序分裂組織の側方に形成される花芽分裂組織として誕生する。その花芽分裂組織の成長の方は、有限である。
 花芽分裂組織は、花器官の4つの異なる形成形態、がく片・花弁・雄ずい・心皮と、外側から中心へと、環域と呼ばれる花芽分裂組織をとりまく同心円の環状領域の外側から中心組織へと向かい順々に形成されて行く。
花芽分裂組織は、花器官の4つの異なる形成形態、がく片・花弁・雄ずい・心皮と、外側から中心へと、環域と呼ばれる花芽分裂組織をとりまく同心円の環状領域の外側から中心組織へと向かい順々に形成されて行く。最内部に形成された心皮の発生開始により、花芽分裂組織頂端部のドーム領域の分裂組織細胞が使い尽くされるが、花芽の発生が進むと、各花器官の原基だけが残り、そのため局所的な細胞の分裂が活発になる。
花芽形成と花成を制御する転写因子をコードする遺伝子は、通常、極めて多く、そのため花の発生過程には、直線的でない遺伝子ネットワークが複雑に関わってくる。しかも、これらのネットワークには、個々の遺伝子が複数の役割を果たしている。
例えば、シロイヌナズナでは、進化によって、APETALA1(AP1)という同一の転写因子が、花芽分裂組織の運命を決定し、次いで花器官の第一環域と第二環域を制御している。別の転写因子AP1は、がく片と花弁の形成に関与し、がく片の腋からの二次花成形成の抑制にも関与している。
花芽分裂組織を、その側部に形成するシュート頂分裂組織は、花序分裂組織と呼ばれている。その側部の原基が、花芽分裂組織になるには、花芽分裂組織決定遺伝子が作用しなければならない。例えば、花芽分裂組織決定遺伝子FLORICAULA(FLO)遺伝子に欠損があるキンギョソウの突然変異体の花序では、花は作れない。またflo変異体では、苞葉bractの腋に花芽分裂組織ができず、代わりに新たな花序分裂組織が形成される。
苞葉は、開花前の蕾全体を包み込む葉のようなものを言う。葉に似ていることが多いことから、包葉とも呼ばれる。また個々の苞を苞片と言う。多くの場合、普通の葉より小さく 、一般に緑色で、花序を包む包葉は、複数からなり総包と呼ばれる。ドクダミやヤマボウシでは総包は 、4枚からなり白色花弁状である。ミズバショウなどサトイモ科では、仏炎包と呼ばれる大型の包葉が特徴となっている。
ザゼンソウは、車山高原では、3月頃、暗紅色の仏炎苞に包まれた肉穂花序をつける。開花する際、肉穂花序が発熱し約25℃まで上昇し、周囲の雪を溶かして、いち早く顔を出す。同時に悪臭も放ち、発熱と悪臭の戦略で、未だ数の少ない花粉を媒介するハエ類を独占し、受粉の確率を上げている。
 花器官の属性を決定する遺伝子は、花のホメオティック突然変異体を通して発見された。一群のホメオティック遺伝子homeotic gene(dʒíːn)は、特定の構造のための遺伝的なプログラム全体を活性化させる主要な発生スイッチとして機能する。それにより、ホメオティック遺伝子の発現が、器官に固有の属性を与えることになる。
花器官の属性を決定する遺伝子は、花のホメオティック突然変異体を通して発見された。一群のホメオティック遺伝子homeotic gene(dʒíːn)は、特定の構造のための遺伝的なプログラム全体を活性化させる主要な発生スイッチとして機能する。それにより、ホメオティック遺伝子の発現が、器官に固有の属性を与えることになる。花器官決定遺伝子は、当初、花器官の属性が変化し、花器官のうちのあるものが、違った場所に形成されるホメオティック突然変異体として同定された。
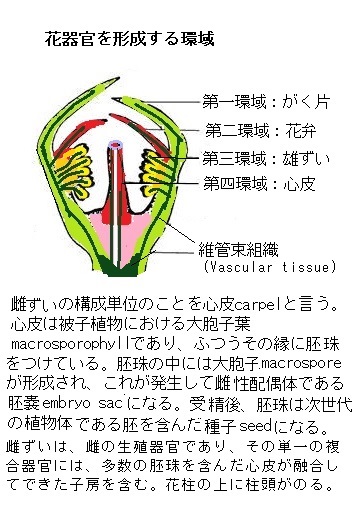
花の4種類の器官、ガク片・花弁・雄ずい・雌ずいに関する変異体のうち、例えば、雄ずいが花弁に転換するようなホメオティック変異体に絞り、それらを交配実験によって分類した。その結果、5種類の主要な花器官決定遺伝子が見つかった。それが、APETALA1 (AP1)、APETALA2 (AP2)、APETALA3 (AP3)、PISTILLATA (PI)、AGAMOUS (AG)である。これらの遺伝子に起きた突然変異は、2つの隣接する環域にある花器官の構造を劇的に変えてしまう。
例えば、ap2変異体では、がく片と花弁を欠く、ap3変異体、あるいはpI変異体では、第2環域に花弁の代わりにがく片が生じ、第3環域では雄ずいの代わりに心皮が形成される。これらの遺伝子の突然変異は、花の発生を妨げないが、花器官の属性を変えるため、ホメオティック遺伝子の定義「生物が正しく発生、分化していくためのカギとなる遺伝子」と一致する。
ap1・ap2・ap3(pI)・agの4つの突然変異を持つ四重変異体の植物では、花芽分裂組織は花器官自体すら形成できず、その代わりに緑色の葉状の器官を形成する。しかも、それらの葉状器官は、正常な花に典型的な輪生状の葉序を形成する。
この実験結果により、シュート頂分裂組織によって形成される花器官の基底状態ground stateは葉であり、この葉状器官が、がく片・花弁・雄ずい・心皮に変形するためには、AP1遺伝子や AP2遺伝子などの遺伝子の活性が不可欠であることが分かった。
ドイツの詩人・自然学者ヨハン・ヴォルフガング・フォン・ゲーテによる、1790年「植物変態論」の著書の中で、「葉という基本的な器官が変形して花となった」と記した観察は正確であった。メンデルの法則が世に知られる100年以上も前に、その洞察力で、「地面の下で湿潤だけを吸収する葉を根と呼ぶ」「すぐに拡張する葉は葉柄ないし茎である」と提唱した。
花器官が劇的に置き換わるパターンによって、3つのグループに大きく分けられ、それぞれA、B、Cに分類された。シロイヌナズナのA遺伝子には、APETALA1 (AP1)とAPETALA2 (AP2)、B遺伝子には、APETALA3 (AP3)とPISTILLATA (PI)、C遺伝子にはAGAMOUS (AG)と、それぞれのクラスは、異なるタイプの活性を表現する。そのため、ABCモデルと呼ばれているが、4種類の花器官と3つの遺伝子グループとでは、数が合わない。一対一対応にはなっていない、という点がABCモデルを考える際のヒントになっている。そして、これらの変異体を組み合わせた多重変異体を作ることで、花器官決定遺伝子の表現型が解明された。
ABCmodelは、2つの遠縁な真正双子葉植物、シロイヌナズナとキンギョソウ双方における観察で、比較的少数の鍵制御因子の組み合わせにより、複雑な結果をもたらす方法が明らかになった。
ABCmodelでは、各環域に形成される器官の属性は、それらの環域における3つの花器官決定遺伝子活性のユニークな組み合わせによって決定されているようだ。
クラスA活性は、単独でがく片という属性を決定する。
クラスA活性とクラスB活性は、花弁の形成に必要である。
クラスB活性とクラスC活性により、雄ずいが形成される。
クラスC活性は、単独で心皮の属性を決定する。
このモデルでは、さらに、クラスA活性とクラスC活性は、花器官の属性決定に際し、それぞれの発現領域では、互いに相手の発現を抑制し、その活性を排除する。
ただ、ABC遺伝子のみによって、葉は花器官に転換しない。シロイヌナズナのクラスE遺伝子は、ABC遺伝子それぞれの活性に不可欠である。ABC遺伝子が同定された後で、花のホメオティック遺伝子がもう一つ、クラスEが発見された。しかも、クラスE遺伝子は、クラスAおよびクラスB遺伝子と組み合わせて発現させると、子葉と普通葉を花弁に変換させることができる。
ABCmodelは、シロイヌナズナとキンギョソウにおける遺伝学な実験で定式化されている。様々な種類の植物の花は、ABCmodelの働きと、制御ネットワークの一部を変更させることで、多様な形に進化させてきた結果のようだ。
目次へ