生物間相互作用biological coaction
地球上には数多くの物質があり、その種類をすべて数えあげるのは不可能と言える。 一方で、物質の種類は無数でも、これら物質は「限られた少数の成分の組み合わせで成り立っているのではないか?」といった考えが紀元前からあった。このような、すべての物質の根源となる成分を元素という。
19世紀にドルトンが、物質の根源となる基本粒子が存在するという仮説を発表した。これをドルトンの原子説という。今日では、無数の実験検証により、物質の根源となる成分は原子であることが証明されている。
元素の正体は原子である。物質の根源を示す概念を元素といい、その元素の実体が原子である。
原子の性質は、その原子が含む陽子の数によって決定される。水素原子・炭素原子・窒素原子・酸素原子それぞれの性質は、その原子が含む陽子の数によって決定される。
例えば、水素原子は1個の・炭素原子は6個の・窒素原子は7個の・酸素原子は8個の陽子があり、それぞれまるで違う性質を持つ。各原子のそれぞれ固有の性質は、各々の陽子の数によって決まる。それぞれに絶対的な相違を生む。
中性子の数が違っても、陽子の数が同一であれば、原子番号は同一であり、同じ元素である。しかし、陽子の数が同じでも、中性子の数が異なれば、違う原子とみなされる。
同じ元素でありながら質量数が異なる原子を同位体という。同一の元素に属する原子は、すべて同一の数の陽子を持つ。原子番号が同一でありながら、中性子数の異なる原子を同位体という。例えば、リチウム6とリチウム7は同位体で、同じ元素である。

| 目次 |
| 1)植物の防御機構 |
| 2)植物-微生物間の有益な相互作用 |
| 3)生物的窒素固定 |
人間も含めて動物たちは、無機物をエネルギー源にはできない。他の生物が作り出すアミノ酸や脂質といった有機物を、直接または間接的に食べて生きている。砂糖はサトウキビやテンサイ(砂糖大根)から得られるエネルギー源であるが、塩は鉱物でエネルギー源に成りえない。このように他の生物に依存する生物を従属栄養生物と呼ぶ。
一方、他の生物が生成する有機物に頼らず生存を全うする独立栄養生物もいる。
陸上植物および緑藻などの緑色植物は、光合成によって太陽エネルギーを利用して自分を支えるエネルギーや有機物を生産している。
化学合成細菌も独立栄養生物であるが、光エネルギーを使わず無機物の酸化還元反応から生存に必要なエネルギーを得ている。化学合成細菌に依存する生物と共に小さな生態系を作っている。
土の中の小さな生き物や微生物は、生物の排泄物や死体などの有機物を分解して無機物に変える働きをしている。自然界の分解者と呼ばれている。それを植物が栄養素として吸い上げるというのが循環の第一歩である。
目次
植物は、自然界において複雑で多様な環境で生息しており、そこには様々な生物間の相互作用がある。
その相互作用が、両者にとって必ずしも必要不可欠とまでは言えないが、明らかに有益に働くこともある。このような双方に利益をもたらす相互作用を相利共生mutualismと呼ぶ。
相利共生の例として、植物-送粉者(花粉媒介者)間の相互作用、窒素固定細菌(根粒菌)とマメ科植物の共生関係、根と菌類の間で起こる菌根形成、葉に内生する菌類であるエンドファイトendophyte(植物体内で共生的に生活している菌や細菌)などがあげられる。
他の生物間相互作用の例には、動物や昆虫などによる食害herbivoryや、微生物病原体microbial pathogenや寄生生物parasiteによる感染、他感作用allelopathy(植物間の化学物質を介した成長阻害)がある。これらはすべての植物にとって有害である。
植物は、これらの有害な生物から自らを守るために精密な防御機構を進化させてきた。一方、有害な生物側も、植物の防御機構を破るため進化してきた。この報復の繰り返す進化過程を共進化coevolutionと呼ぶ。これもまた複雑な相互作用が働いている。
植物と他の生物との相互作用を、単純に有益か有害かのいずれかにあてはめるには無理がある。通常、菌根菌(菌根とは土壌中の糸状菌filamentous fungiと共生した状態の植物の根のことであり、菌根に形成される菌を菌根菌 と言う)は、植物の適応性を促進させる相利共生生物と見られているが、実際には寄生と相利共生のどちらにも当てはめる。同様に、ある植物にとって動物の捕食は有害だが、他の植物にとって花を付ける茎の数を増やす切っ掛けになるため有益に働く。
更に、植物と相互作用をして、有害な作用を全く起こさず利益だけを受容できる生物も存在する。このような植物側だけが利益を享受する相互作用を、片利共生commensalismと呼ぶ。片利共生生物は、他の有害な生物から植物を守るために、植物にとって利益に働く。例えば、非病原体である土壌中の根圏細菌(根の周りに生息する細菌)や菌類は、植物に対して害を与えないが、植物の免疫システムを刺激して、植物を病原性微生物から身を守ることになる可能性が生じる。
植物は、栄養素として鉄(Fe)・マグネシウム(Mg)・リン(P)・カルシウム(Ca)
などいくつかの金属イオンを必要としている。そのいっぽう、土壌中にあるアルミニウム(Al)などは毒性があり成長を妨げる。これらの金属イオンは土壌水中に遊離しているものばかりではない。土壌鉱物や腐植酸などに特殊な形で結合して、植物に吸収されにくい状態にあるものもある。その一方では、植物は根からいろいろな物質を分泌する。その中には、根酸と呼ばれるアミノ酸や有機酸などが含まれている。
根圏では、根から分泌された有機酸organic acid(酸性を示す有機化合物全体の総称)やアミノ酸が、根の周辺の土壌中の微生物菌相に大きな影響を与えている。根圏の微生物は、土壌中の有機物を分解して不溶化されている栄養素イオンを遊離化したり、微生物自体の代謝産物が、栄養素となったりして植物に有利な環境を提供する。そのため、分泌された有機酸やアミノ酸は、根圏細菌の菌相の成立を促進する。人の腸内細菌群が人の栄養、健康にとって重要な働きをしていることと共通する。
植物は、根から分泌した有機酸によって、有害な金属イオンを溶脱することで、植物の生育に有利な環境を作る一方、必要とする元素を、栄養素として利用しやすくする。それにより発根が促進される。
リン酸イオンや鉄イオンの多くは、土壌鉱物や土壌中の腐植酸などにキレート結合(錯体)したり、不溶性になったりして、多くは吸収されにくいため、リン酸欠乏や鉄欠乏になりやすい。根から分泌されたクエン酸やリンゴ酸などは、これらのリン酸イオンや鉄イオンと錯体を形成し、遊離可溶化して吸収されやすくする。また、イネやムギなどのイネ科の植物は鉄欠乏におちいると、大量のムギネ酸という有機酸を分泌し、鉄を可溶化し、キレート化によって体内に吸収して鉄欠乏を回避する。
Alは地 を構成する主要な元素の一つである。pHが中性 に近い土壌では主に珪酸アルミニウムとして存在しているが、酸性土壌で、Alイオンとして溶解し、 植物に吸収され、根に強い障害をもたらす。
一般的に養分に乏しい酸性土壌における、ムギ類の根の伸長阻害など、酸性土壌は、世界の農耕地 の30~40%を占めるが、その作物の生産性は低い。土壌中 の元素の7%を占めるアルミニ ウム鉱物が、pHの低下でアルミニウムイオンとして土壌水中に溶解して、植物に強い毒性を示す。
Alは、特にリン酸との反応性が高く、不溶性のリン酸アル ミニ ウムを形成して、リン酸欠乏をもたらす。特に、低濃度のAlが、極めて短時間で、根の伸長阻害を引き起こす。コムギの根の伸長が強く抑制され、伸長域の細胞死も見られる。トウモロ コシでは、最も強Al毒性が現れるのが、根の分裂域と伸長域の中間域の細胞で、根の伸長の抑制に伴い、根が太くない固くなる。それにより、伸長域の根の表面に亀裂が生じる。分裂中期のタマネギの根端では、7~8時間後には、細胞分裂が停止する。
Alの毒性に耐性を示すエンドウやコムギ系統の耐性植物は、Alイオンを感知すると、リンゴ酸を分泌する。リンゴ酸は、Alイオンとキレート結合物を生成してAlを土壌から溶脱しやすくする。これが、Alイオンの解毒である。
Alイオンは、植物遺伝子を発現させ、それにより、リンゴ酸の輸送体合成が増加促進され、リンゴ酸の分泌が増加するという。同じようにオオムギ、トウモロコシ、インゲンなどの耐性系統は、クエン酸を分泌する。クエン酸もリンゴ酸と同じようにAlと錯体を形成する。蕎麦は有機酸の一種、シュウ酸を分泌してアルミニウムによる酸性害を軽減している。茶も、根が分泌する有機酸によって有害なアルミニウムの動きを封じ込めている。
アルカリ性の土壌に育つヒヨコマメは、根からクエン酸を分泌して根の周りを酸性化し、アルカリ性では溶けにくい鉄を吸収しやすくしている。
植物に害を及ぼす可能性のある生物に対する第一の防御線は植物の表面にある。クチクラ(植物では葉の表皮の表面にあるワックス層)や表皮、その他の物理的障壁が、細菌や菌類(カビや茸、酵母で、細胞壁をもち光合成をしない)、昆虫などの侵入を阻む。
第二の防御線は、殆どの場合、生化学的な防御機構であり、恒常的防御機構constitutive defenseと誘導的防御機構inducible defenseがある。
恒常的防御機構は常にあるが、その中でも物理的障壁と有毒な二次代謝産物は、昆虫や動物の食害に対する植物の恒常的防御機構の二大戦略となっている。
誘導的防御機構は、動物や昆虫及び病原体の攻撃に応答して、遺伝子発現や代謝を変化させることができる、特異的な認知機構とシグナル伝達経路を備える。
目次
藻類と菌類の共生関係は、最初に陸上植物が出現した約4億5千~5億年前頃からあったようだ。菌類と藻類、またはシアノバクテリアとの絶対共生関係と言える最初の地衣類が、約4億年前の化石に現れるが、これは陸上植物と共生する最初の菌類が出現した頃と重なる。緑色植物の陸上への進出が、菌類との共生関係によって可能になったことが推測される。
 |
| 奥蓼科逆川橋辺りのサルオガセ |
地衣類は、一般に、コケ植物の蘚苔類(センタイルイ)などとともに「こけ」と認識されている。実は、「こけ」は「むし」などと同じく雑多な小さな生物群の総称であり専門用語ではない。それで、地衣類を「こけ」と呼んでも誤りではないが、コケ植物、あるいはコケ類と同定すると、形態的にも生態的にも違うことになる。
地衣類を構成している菌と藻による、地衣化の本質は、菌類が藻類に安定した住み家と生活に必要な水分を与えるかわりに、 藻類が光合成で作った有機化合物を栄養として生存を全うしている。その共生関係は、極めて密接で、地衣体の形態・生理機能、繁殖方法などは、単独の生物のように遺伝する。まるで独立した生物のように見える一体化である。
地衣類は世界中に広く分布し、1万4千種とも2万種とも言われ、このうち1,602種が日本から記録されている(2,004年)。
他の植物が生育できないような厳しい環境に進出できるのが、地衣類の特徴で、都会でもコンクリートの表面に懸垂する光景が見られる。奥蓼科逆川橋辺りのサルオガセ(猿尾枷)が有名ですが、蓼科御泉水自然園や御射鹿池付近でも、普通に、見られる。「サルオガセ」は、長く木の枝から垂れ下がり、空気中の水蒸気を吸って自立して育つ。霧藻(きりも)とも呼ばれ、水分と光合成だけで成長し、栄養を他から奪い取る寄生植物ではない。
また、砂漠のような厳しい環境でも地衣類はわずかな水分を利用して繁茂している。
自然界では、多種多様な有用微生物が、陸上植物に定着している。例えば、外生菌根菌(マツ科・ブナ科・カバノキ科・ヤナギ科などの樹木の根に菌類が侵入し、外生菌根という特殊な形態の菌根を形成、マツタケやベニタケ科・イグチ科など、菌糸が、植物の細胞内に侵入することはない)及び内生菌根菌(宿主植物の細胞の内部まで侵入するタイプのカビ総称、一括りにはできないほど多様である)、根や葉の表面にバイオフイルムを形成する細菌、植物内生細菌、そして根粒または茎粒に含まれる窒素固定細菌などがこれにあたる。
ラン科植物では種子発芽後の幼植物体seedling plantにおいて、内生菌根菌から植物に炭素化合物が供給されるため、ラン科植物の中には葉緑素を失ったものもある(無葉緑ラン)。
バイオフィルムbiofilm(菌膜)は、基質に付着した細菌や微細な藻類と、それら自身が産生する粘液とともに作る膜状の集合体で、多糖類で構成された薄い膜を指す。バイオフィルム内の細菌は、細胞外多糖extracellular polysaccharide (EPS)を分泌するため、EPSは、バリアーや運搬経路の役割を果たし、環境変化や化学物質から内部の細菌を守る。その結果、抗生物質や免疫に対する抵抗性が高くなり、恒常性が維持されて、細菌などの生息密度の高い閉鎖的なコロニーが形成される。
目次
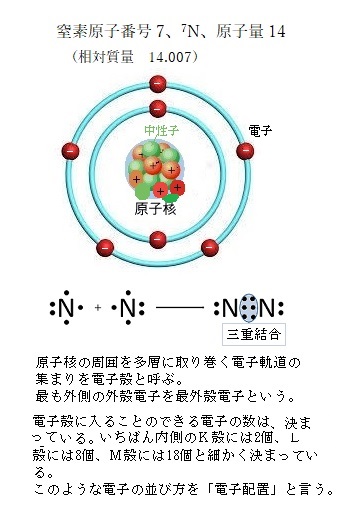 窒素原子番号7、原子量 14(相対質量 14.007)である。
窒素原子番号7、原子量 14(相対質量 14.007)である。窒素化合物(硝酸態窒素を除く)は、土壌中には1%にも満たない程度しか存在しない。また、土壌の肥沃度とはあまり関連が無い。
生物的窒素固定による、大気中のN2をアンモニウムイオンへ変換するシステムが、分子状窒素を生物の体内で活用するためには、不可欠な鍵となるばかりか、その大部分を担っている。
植物細胞内の多くの有機化合物は、窒素を含む。窒素は、核酸やタンパク質の構成成分であるヌクレオチドやアミノ酸に含まれる重要原子である。植物中で原子番号7の窒素より多く含まれる元素は、原子番号1の水素元素・原子番号6の炭素元素・原子番号8の酸素元素だけである。
それなのに、農業分野では、無機態窒素の施肥により、劇的な生産増加に繋がった。土壌中の窒素形態は、有機態窒素と無機態窒素の2つに大別される。無機態窒素は、アンモニア態窒素(NH4-N)と硝酸態窒素(NO3-N)から生じる。大半の植物は、土壌中の硝酸態窒素を吸収する。
土壌中のアンモニア態窒素は、植物が吸収可能な硝酸態窒素の一つ前の形で、やがて微生物等により硝化(酸化)され、硝酸態窒素に変換され、植物に利用される。その硝化行程は、通常、アルカリで+イオンであるアンモニアが、亜硝酸によって、-イオンである硝酸硝酸に酸化される形態変化である。吸収された硝酸態窒素は、植物生体内で、アンモニア態窒素に変換され、その後アミノ酸やタンパク質に合成される。
 硝化にはアンモニア酸化細菌と亜硝酸酸化細菌の2種類の独立栄養硝化細菌(→化学無機栄養生物)、更に従属栄養硝化細菌が関与する。独立栄養硝化細菌の場合、上記2種類の細菌によりアンモニアが硝酸に酸化されるが、従属栄養硝化細菌の場合は、1種類の細菌がアンモニアを硝酸に酸化する。従属栄養硝化細菌1個の硝化力は、独立栄養硝化細菌の硝化力の1,000分の1以下であるが、生息数を考慮すると、グローバルでは両者の硝化力は同程度だという。
硝化にはアンモニア酸化細菌と亜硝酸酸化細菌の2種類の独立栄養硝化細菌(→化学無機栄養生物)、更に従属栄養硝化細菌が関与する。独立栄養硝化細菌の場合、上記2種類の細菌によりアンモニアが硝酸に酸化されるが、従属栄養硝化細菌の場合は、1種類の細菌がアンモニアを硝酸に酸化する。従属栄養硝化細菌1個の硝化力は、独立栄養硝化細菌の硝化力の1,000分の1以下であるが、生息数を考慮すると、グローバルでは両者の硝化力は同程度だという。植物にとって窒素は重要でありながら、自然界からは十分に供給されていないため、施肥が必要になる。
窒素は、大気中に容量比で78%の多量の分子状窒素N2が含まれている。その大部分は、生物にとって直接利用できない窒素である。大気中の窒素を利用するには、2つの窒素原子間の極めて安定した三重結合triple bond(N 三 N)を切断して、アンモニアイオンや硝酸イオンを生成する反応が必要となる。
原子は、最外殻に不足する電子を補い、それに必要な数を共有結合で他の原子から得ようとする。窒素の最外殻電子はL殻であるため、安定するの電子数は8個である。窒素原子番号は、7であるため、最外殻電子は5個となる。最大3つの電子を補うため他の原子と3つ共有結合を作り出せば安定する。共有結合では、原子同士が電子を共有して、それぞれの最外殻を満たすが、その電子は電子間を移動することはない。各共有結合の角度と長さによって決まる正確な三次元構造をなしている。
窒素固定nitrogen fixationと言われる反応には、工業的過程によるものと、自然界で起こるものとがある。
分子状窒素と水素を結合させるアンモニアガスを合成するには、200℃の高温と約200気圧の高圧条件を整え、鉄が殆どであるが、金属を触媒して反応させる。この極端な反応条件は、反応の高活性化エネルギーを作り出すためである。
ハーバー・ボッシュ法Haber-Bosch processと呼ばれる、この窒素反応は、窒素肥料を含む多くの農業用の製品を工業的に製造する出発点となった。世界中で工業的に生産される窒素肥料は、年間1.1億トン以上となる。だが、自然界では、年間約1.9億トンの窒素を固定している。
稲妻は、自然界における窒素固定の全体の8%を担っている。稲妻は、水蒸気と酸素を、非常に反応性に富むヒドロキシルラジカルhydroxyl radical(・OH と表される。活性酸素の中でも、最も反応性が高く、最も酸化力が強い)・遊離水素原子・遊離酸素原子に変換し、これらが分子窒素を攻撃して亜硝酸(HNO3)を生む。この亜硝酸が、雨とともに地表に落下する。
窒素固定全体の2%が、ガス状の一酸化窒素(NO)とオゾン(O3)の間の光化学反応により亜硝酸を生じさせる。
残りの90%が、生物的窒素固定に由来し、細菌やシアノバクテリア(藍藻)が分子窒素を固定しアンモニアを生む。このアンモニアが、水に溶けてアンモニウムイオンを生じさせる。
NH3 + H2O ➡ NH4+ + OH-
農業的には、工業生産された窒素肥料は、経済的にも環境的にもコストが掛かり過ぎる。その観点からも生物的窒素固定は、重要な意味合いがある。
生物による窒素固定は、大気N2を、アンモニウムイオンへ変換する過程であり、そのシステムの大部分を占めている。その窒素固定が、分子状窒素を、生物体内における窒素循環系に入れるための絶対的な条件になっている。
自由生活型free-living単性バクテリアと共生symbioticバクテリアは、窒素固定をする。ある種のバクテリアは、大気中の窒素をアンモニアへ変換することができる。これらの窒素を固定する原核生物prokaryote(明確な核膜を示す細胞核を持たない細胞からなる生物、納豆菌・ビフィズス菌・黄色ブドウ球菌などの細菌類や藍藻類のシアノバクテリアなどが分類される)の殆どは、土壌中の他の生物と共生せずに単独で生活する。
植物と共生関係をもつ原核生物は、宿主植物に固定した窒素を直接提供し、代わりに他の栄養素や炭水化物を受け取る。このような共生は、根粒で起こり、根粒は植物の根に形成された窒素固定細菌を含んでいる。根粒は、マメ科植物の根についてこぶを作らせ、その中に生息しながら共生の関係を結ぶ細菌の総称である。そのこぶを根粒という。最も一般的な共生は、マメ科植物と土壌細菌のリゾビウム属Rhizobiumとの間で成立する。遊離状態では運動性の杆菌(かんきん;形態が棒状または長楕円状で長径と短径がある細菌。結核菌・大腸菌・腸チフス菌など多くの細菌が含まれる。枯草菌・炭疽菌のように胞子を作るものもある)であるが、適当な植物があると根毛内に侵入し、皮部を刺激して根粒を形成し、宿主から無窒素化合物(脂質と炭水化物と粗繊維とに分けられる)・塩類・水を受け取り、一方、空気中の窒素を固定して宿主に提供する。マメ科ゲンゲ属のレンゲソウなど、マメ科植物の数種は、緑肥と呼ばれ窒素肥料として活用される。
 北米原産のマメ科の落葉高木のニセアカシアは、日本には明治の初めに渡来した。林業分野では、ニセアカシア・アカシア類・ハギ・イタチハギ・ネムノキ・エニシダなどのマメ科樹木や、ハンノキ・ヤシャブシ・ヤマモモ・グミ・モクマオウなどの非マメ科根粒植物は、土壌改良に貢献する肥料木と呼ばれ、荒蕪地や砂防地の造林木、あるいは混植樹として利用されいる。
北米原産のマメ科の落葉高木のニセアカシアは、日本には明治の初めに渡来した。林業分野では、ニセアカシア・アカシア類・ハギ・イタチハギ・ネムノキ・エニシダなどのマメ科樹木や、ハンノキ・ヤシャブシ・ヤマモモ・グミ・モクマオウなどの非マメ科根粒植物は、土壌改良に貢献する肥料木と呼ばれ、荒蕪地や砂防地の造林木、あるいは混植樹として利用されいる。ニセアカシアは、窒素固定をする根粒菌と根で共生しているため、治山緑化の 現場で積極的に導入されてきた。冷温帯に分布する落葉樹の殆どは、秋に紅葉(黄葉)する。これは土壌中に窒素含有量が少ないため、葉を落とす前に、窒素を多く含む葉緑素を分解して、樹体内に再吸収するためと見られている。
根粒菌と共生している植物の葉の多くは、緑のまま萎れてしまう。窒素が常に供給されるので、落葉する前に、葉緑素を分解して再吸収する必要がないためとみられる。そのため、これら樹種の窒素分に富んだ葉が、大地に大量に落ちると、土壌は富栄養化する。自然植生におけるマメ科樹木の年間窒素固定量の調査結果の数例を見ると、平均して50~150 kg/haの間と言う。 もう1つ一般的な共生は、植物に内生する放線菌で、カバノキ科のハンノキのような放線菌根生植物actinorhizal plantと呼ばれる数種の樹木と、土壌細菌のフランキアFrankia属との間で起こる。ハンノキからは、良質の炭ができるので、炭を生産していた時代には盛んに伐採されたと言う。ハンノキの根には、放線菌が共生し、根粒を形成する。その放線菌は、空中窒素を固定し、湿原のような貧栄養環境であっても、ハンノキを高木として生長させる。
ハンノキ属についての調査は多い。アラスカの石礫の不毛地で、ハンノキとチョウノスケソウを主体とした林では、毎年62kg/haの窒素の蓄積が生じ、土地が肥沃化する。すると、針葉樹が優生樹種となる。そのような現象が欧州のアルプスの氷河跡地においても起きていると報告されている。アメリカのカリフォルニア湖では、湖畔に面してハンノキ林が密生し、湖畔周囲の土壌及び湖水の水が富栄養化し、プランクトンが異常発生している例がある。
作物の栽培においてもっとも多用される肥料は窒素肥料である。戦後の「緑の革命」において、多量の窒素肥料を投入することにより作物の生産量は飛躍的に上昇し、世界人口の急激的な増加に対応することができた。ただし、民族的・宗教的・政治的独善が関与した場合は、この限りではない。
窒素固定細菌の一種は、C4光合成を行う、サトウキビやススキのようなイネ科とも共生している。草本植物は、光合成様式の違いにより基本的には、C3植物とC4植物に大別される。C4植物の窒素利用効率は、C3植 物の約2倍も高い。これは、細胞が窒素の栄養状態を検知して、二酸化炭素の濃縮に携わる炭酸同化系酵素の遺伝子を選択的に制御する仕組みが、C4植物には備わっているためである。
なお炭酸同化とは、エネルギーを付加し、二酸化炭素から有機物を作ることを言う。光エネルギーを使う炭酸同化を、「光合成」と呼ぶ。炭酸同化に利用されるエネルギーは、光エネルギーだけない、もう一つある。光のエネルギーの代わりに、無機化合物の酸化で生じる「化学エネルギー」で、CO2から炭水化物を合成する炭酸同化の働きをする生物がいる。これを「化学合成」と言う。このクロロフィルをもたずに、化学合成を行う特殊な独立栄養細菌が化学合成細菌と総称されている。
アンモニウムイオンを酸化して亜硝酸イオンにする亜硝酸菌(あしょうさんきん)や、亜硝酸イオンを酸化して硝酸イオンにする硝酸菌などがある。これらの生物は、地球全体の炭酸同化の割合としては少ないが、窒素循環などに、極めて大きな役割を果たしている。
 いくつかの光合成細菌は別として、生物圏の光合成独立栄養生物は、大気CO2同化のためカルビン-ベンソン回路(葉緑体のストロマに存在する反応径路で、ATPとNADPHを用いて、二酸化炭素を固定・還元してグルコースとする一連の代謝反応)を使っている。約3.5億年前から顕著となったCO2の減少とO2濃度の上昇は、光合成生物に光呼吸photorespirationに対応する一連の適応機構を獲得させた。
いくつかの光合成細菌は別として、生物圏の光合成独立栄養生物は、大気CO2同化のためカルビン-ベンソン回路(葉緑体のストロマに存在する反応径路で、ATPとNADPHを用いて、二酸化炭素を固定・還元してグルコースとする一連の代謝反応)を使っている。約3.5億年前から顕著となったCO2の減少とO2濃度の上昇は、光合成生物に光呼吸photorespirationに対応する一連の適応機構を獲得させた。植物は、二酸化炭素が足りないような条件では、ルビスコという酵素が炭素5個からなる物質に酸素をくっつける反応を行ない、その結果出来た物質が還元力とエネルギーを使いながら、様々な反応を経て二酸化炭素と炭素3個からなる物質を作り出す。
ルビスコは、炭素固定において中心的な役割を果たしているにも関わらず、著しく非効率である。通常の酵素は、毎秒1,000個の分子を処理できるが、ルビスコは毎秒たった約3分子の二酸化炭素しか固定できていない。植物細胞は、この処理速度の遅さを、大量の酵素を作ることによって補っている。そのため、葉緑体は、ルビスコで満たされ、その量は葉緑体に含まれるタンパク質の半分を占めている。これにより、ルビスコは、単一の酵素としては地球上で最も豊富に存在する酵素となっている。
このような適応機構には、大気からCO2と炭酸水素イオンの能動吸収やルビコン近傍でCO2濃縮を行う機構が働く。
C4光合成は、陸上植物が低CO2環境のため、光合成の制限要因を補償するためのCO2の濃縮機構の1つとして進化してきた。地球上で、生産性が最も高い陸上植物のトウモロコシ・サトウキビ・ソルガム(タカキビやコウリャンなどで知られるソルガムは、食用としての種子利用では、コムギ、イネ、トウモロコシ、オオムギに次ぐ生産高世界第5位の穀物)などは、ルビスコrubiscoの活性を上げるための機構を持っている。
葉緑体chloroplastのchloroは、接頭辞「緑の」、plastは連結形「(生物用語では)形成されたもの」と言う意で、二重膜に包まれた細胞小器官のもう1つのグループである色素体の一種である。光合成の場である。葉緑体には、内膜と外膜に加えてチラコイドthylakoid(ギリシャ語;袋状の意)と呼ばれる、扁平な袋状の小胞がある。そのクロロフィルを含む扁平な袋が、積層してグラナgranaと呼ばれる構造を形成する。1,883年、Meyerは、光学顕微鏡による観察で、葉緑体には無色の礎質があり、その中の葉緑体が貯留する粒状体を見出しグラナと名付けた。「グラナ」とは「粒」や「粒子」と言う意味がある。
光合成の光化学反応で働くタンパク質と、クロロフィルやカロテノイドなどの色素は、チラコイド膜に埋め込まれている。そのチラコイド膜が積層するグラナは、ストロマラメラstroma lamellaと呼ばれる膜系で繋がっている。stromaは「基質」、lamellaは「膜」、葉緑体の構成する、それぞれに、葉緑体研究の発展過程が序次に物語られている。
チラコイドを囲む流動性の物質は、ストロマstromaと呼ばれ、ミトコンドリアのマトリックに類似している。ストロマは、地球上で最も多量に存在するタンパク質で、光合成で二酸化炭素を有機酸に固定する反応を触媒するルビスコrubiscoは、このストロマに存在している。
ルビスコの小サブユニットは、核ゲノムにコードされているが、ルビスコの大サブユニットは、葉緑体の核ゲノムにコードされている。両ゲノムによる各サブユニット及びその他のタンパク質が協調した発現が、葉緑体の発達や分裂に必要である。
C4植物の光合成での窒素利用効率は、C3植物のそれよりも2倍ほど高い。両種の植物のC/N比 には、概ね2倍の差がある。窒素栄養は、光合成を最も大きく規定する化学環境要因であれば、光合成生産性向上への基本的な戦略は、植物が、最小の窒素投資で最大量の炭素を固定できるかにある。
コケ植物やシダ植物を含む陸上植物の7割以上の種は、グロムス門に属するアーバスキュラー菌根菌と共生する。この糸状菌の一群は、窒素固定細菌との共生とは対照的に、土壌中に普遍的に存在し、植物からエネルギー源となる主にブドウ糖の供給を受ける代わりに、土壌中の希薄なリン酸を集め、宿主植物に供給する。
宿主の系統関係から植物の陸上化にともない共生が確立されたと考えられている。すでに4億年以上まえの化石において、アーバスキュラー菌根菌が感染したと考えられる構造が認められている。このアーバスキュラー菌根菌との共生の起源が古いこと、及び共生の宿主特異性が低いことから、宿主のもつ共生遺伝子は、陸上植物に広く保存されていると考えられている。マメ科のモデル植物の研究において明らかにされたアーバスキュラー菌根菌との共生に必要な主要な遺伝子は、イネとアーバスキュラー菌根菌との共生にも必要であることが立証されている
目次