| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成
花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1)
遺伝子の発現(2) 遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝
減数分裂と受精 対立遺伝子と点変異 疾患とSNP 癌変異の集積 癌細胞の転移 大腸癌 細胞の生命化学
イオン結合 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 シグナル伝達 キク科植物
陸上植物の誕生 植物の進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応
根による水吸収 稲・生命体 胞子体の発生 花粉の形成 雌ずい群 花粉管の先端成長 自殖と他殖
フキノトウ
| 目次 |
| 1)減数分裂第1分裂と減数分裂第2分裂 |
| 2)真核生物の体細胞周期 |
| 3)減数分裂には倍加した相同染色体の対合が必要 |
| 4)二価染色体を作る倍加した母方と父方の染色体の間で交差が起こる |
| 5)相同染色体は対合と交差を経て適正に分離する |
1)減数分裂第1分裂と減数分裂第2分裂
有性生殖に関わる基本的な周期の解明は、1888年の報告されたテオドール・ボヴェリ(Theodor Boveri;ドイツの生物学者)の発見から始まった。回虫の受精卵には、4本の染色体があるのに、配偶子の精子と卵は、2本1組しかない一倍体であることが明らかになった。他の細胞は、体細胞はもとより、いずれは配偶子となる生殖系列細胞も含めて二倍体である。しかも、母親由来の染色体と父親由来の染色体が1セットずつ、計2セットを持っている。
 二倍体生物の有性生殖に直接かかわる生殖細胞(germ cell)、即ち配偶子(gamete;gǽmi t)は、殆どの細胞と異なり一倍体であるため、各配偶子には23本の染色体が1組あるだけだ。しかも、殆どの生物は、雄と雌で異なる新たな遺伝子を含む配偶子を作り上げている。動物の配偶子は、大型で運動性のない卵と、小型であるが活発に運動する精子がある。その違いは、卵が大量の細胞質を抱えているが、精子にはそれが殆どないことによる。この特異な2種類の一倍体である配偶子が、融合して二倍体の細胞の受精卵、即ち接合子(zygote;zái
out)となり、やがて新たな遺伝子を受け継ぐ個体を誕生させる。そのため2種類の配偶子は、染色体を正確に半分に分ける特殊な細胞分裂をして作られる。このような細胞分裂を減数分裂と言う。
二倍体生物の有性生殖に直接かかわる生殖細胞(germ cell)、即ち配偶子(gamete;gǽmi t)は、殆どの細胞と異なり一倍体であるため、各配偶子には23本の染色体が1組あるだけだ。しかも、殆どの生物は、雄と雌で異なる新たな遺伝子を含む配偶子を作り上げている。動物の配偶子は、大型で運動性のない卵と、小型であるが活発に運動する精子がある。その違いは、卵が大量の細胞質を抱えているが、精子にはそれが殆どないことによる。この特異な2種類の一倍体である配偶子が、融合して二倍体の細胞の受精卵、即ち接合子(zygote;zái
out)となり、やがて新たな遺伝子を受け継ぐ個体を誕生させる。そのため2種類の配偶子は、染色体を正確に半分に分ける特殊な細胞分裂をして作られる。このような細胞分裂を減数分裂と言う。減数分裂では、1回のDNA複製に続いて2回の細胞分裂が起こる。二倍体細胞が通常の細胞分裂で分裂する場合は、持っている2セットの染色体を前もって2倍にする。これにより、その細胞分裂で新たに誕生する2つの娘細胞それぞれが、完全な母方染色体1セットと父方染色体1セットからなる、ヒトの染色体セットを受け継ぐことになる。
減数分裂では、通常2セット分ある全染色体が、最終的に1セットになり、それが精子と卵からなる配偶子を作る。その前に起こる染色体倍化の現象は、通常の細胞と同様で1回である。生殖系列細胞は、その後独自の過程を経る。DNAの複製を行なうことなく、連続して2回の細胞分裂が起き、むしろ最終的には、細胞内の染色体数を半減させて4個の一倍体細胞である配偶子を作る。生物40億年の進化の歴史は、苛烈な自然選択に、無駄なく合理的応えられる生命体の存続しか認めなかったことを物語る。それ以後の種の存続のために不可欠な突然変異が必ず生じる仕組みを、進化として積み重ねる原動力とした。
生殖系列細胞の減数分裂は、通常の細胞分裂を少し変えていく。DNA複製を完全に省略すれば、二倍体細胞は細胞分裂を1回することで、2個の一倍体を簡単に作れるはずだ。しかし生殖系列細胞は、その後の減数分裂に特異な手順を踏ませた。それを理解するためには、ここでも生物40億年の進化の歴史の重みをわきまえる必要がある。
ヒトの減数分裂は、卵巣や精巣にある特定の二倍体生殖系列細胞で起こる。これらの生殖系列細胞は、体細胞と同じく二倍体で、それはそれぞれの染色体が父方と母方に由来のため、2対存在するようになる。生物学では、父方相同染色体、母方相同染色体と呼ぶ。減数分裂の最初の段階で、これらの染色体は、すべて倍加し2倍になった染色体コピー同士は、通常の細胞分裂の場合と同様に、互いに接着している。ところが、次の段階は、減数分裂に特有で、倍加したそれぞれの父方染色体がまず定位置に並び、その後、それに対応する母方の倍加した相同染色体が接着して対を作る。この対合(pairing)という過程のおかげで、相同染色体は、その後の2回の細胞分裂に際しても正確に分離し、最終的にできる配偶子それぞれに完全な一倍体の染色体セットが入ることができる。
減数分裂で2回連続して起こる細胞分裂、減数分裂第1分裂と減数分裂第2分裂で生じる4個の一倍体細胞それぞれには、完全な染色体セットが分配される。一倍体の娘細胞に父方と母方どちらの相同染色体が振り分けられるかはランダムなので、ここで生じる一倍体細胞からできる配偶子は、それぞれ父母の由来が異なる組み合わせの染色体を持つ。
つまり、減数分裂では、二倍体細胞1個から遺伝的に異なる4個の一倍体細胞ができ、それぞれの染色体は親生殖系列細胞の染色体数の半分になる。これが、通常の細胞分裂では、二倍体細胞1個から遺伝的に同一の二倍体の娘細胞が2個できるのとは異なっている。通常の細胞分裂と減数分裂は、数時間以内に完了するのが普通だが、生殖系列細胞の減数分裂第1分裂は、「倍加した相同染色体の対合」⇒「倍加した相同染色体の対が中期紡錘体に並ぶ」⇒「第1分裂後期に相同染色体が分離」といった前期Ⅰの段階が長いため、数日あるいは数か月、場合によれば数年かかることもある。
目次へ
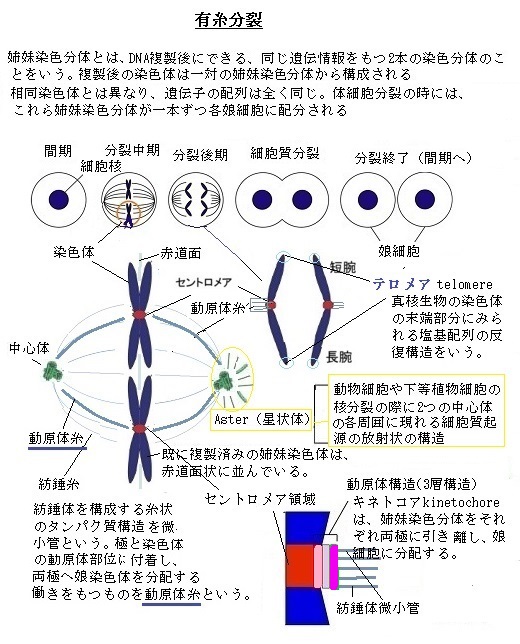
真核細胞の細胞周期は、通常4つの期間で構成される。それを、顕微鏡で観察して最も印象的なのが、有糸分裂と呼ぶ核分裂と、その後に生じる細胞質分裂と呼ぶ2つの過程である。有糸分裂した姉妹染色分体は、中期紡錘体が伸ばす微小管によって引き離されて細胞の両極に分かれ、引き続く細胞質分裂では細胞質が割れて2つの細胞が生み出される。この2つを合わせて細胞周期のM期(Mitotic;分裂 phase)という。典型的な哺乳類細胞では、M期全体で約1時間かかる。これは細胞周期全体から見れば、ごく僅かな時間である。
M期から次のM期までの間は、間期(interphase)と呼び、間期はさらにG1期、S期、G2期に分けられる。この3つの段階が細胞周期(cell cycle)の残りの段階である。 間期の細胞は多数の遺伝子を転写しているが、染色体として機能するには、DNA分子に遺伝子を担う以外の、例えば遺伝子を発現するなど機能も要る。DNAは細胞分裂の度に複製され、そのコピーは2個の娘細胞に確実に分配されなければならない。
間期には、染色体は核内で長く伸びて絡まった細長い糸状のDNAとして存在している。間期の染色体は、細く長く、しかも染色体同士が絡み合うようになっている。それでも核内にきちんと配置さえている。その染色体が、間期の一時期、細胞分裂のため2倍となる。2倍になると、細胞は有糸分裂、つまり核が分裂するM期に進行できるようになる。この有糸分裂では、複製で倍加した染色体が凝縮し、遺伝子の発現が殆ど止まり、核膜が壊れて微小管などのタンパク質からなる紡錘体が形成される。
あらゆる真核細胞では、間期や有糸分裂期を問わず、染色体にDNAがぎっしりと詰まっている。ヒトの第22染色体は、約4,800万塩基対からなり、DNAを伸ばせば1.5cm位になる。それが有糸分裂期には僅か2µm(0.002 ミリメートル)で、DNAは1万倍近く凝縮されている。この圧縮もタンパク質によるもので、DNAをらせん状に巻き、折り畳んで高次構造を作り、さらに何段にも重ねている。ちなみに間期の染色体は、その有糸分裂期の20分の1程度だが、それでも小さく圧縮されている。
間期を顕微鏡で観察すると、単に細胞が大きくなるための休憩時間のように見えるが、実は増殖するため忙しく働いている。そのS期(synthesis; sínθsis合成 phase)に、細胞はDNAを複製する。S期の前後は、「ギャップ期(gap phase)」という細胞が大きくなる時期である 。 G1期とG2期の2期がある。これらのギャップ期に、増殖のための条件が適切であるか、G1期に続くS期における有糸分裂への準備が完了しているかなど、細胞は細胞内外の環境を監視している。G1期とG2期は、そのまま次の段階に進むか、それとも休止して次への準備をもう少し続けるかを、細胞が決定する特定の時点である。
細胞周期制御系は、細胞周期の主要な過程が、正しい順序で行なわれているかを3つの主要な移行点により調節している。S期では、DNA複製や倍加した相同染色体の分離など重要な過程が行なわれているが、制御系では、G1期、G2期、M期の特定の移行点で、細胞外あるいは細胞内の条件が好ましくない場合、周期を一時的に止めることができる。M期では、すべての染色体が、正しく有糸分裂紡錘体に接着したかを確認してから、倍加した染色体を有糸分裂する。G1期からS期への移行時の制御系は、DNAの複製開始前に、細胞増殖に適した環境が、その細胞の内外で整っているかどうかを確認する。動物では、細胞の外部環境に、増殖に十分な栄養と特定のシグナル分子がなければならない。これらの外部環境が好ましくない場合、細胞はG1期の進行を遅らせる。時には、特殊な休止状態であるG0期(Cゼロ期)に入ることもある。
動物では、G1期からS期への移行期が、細胞周期制御系が細胞周期を制御するポイントとして特に重要である。他の細胞からのシグナル、より多くの細胞が必要と時は、細胞増進を促進し、必要でない時は阻止する。そのため細胞周期制御系は、体内の組織における細胞数の調節に中心的な役割を果たしている。制御系が機能せず細胞分裂が過剰になれば癌を発症する。
G2期からM期への移行期では、制御系は有糸分裂に入る前に、DNAの損傷は全て修復されたか、複製が完了したかを確認し、DNAが不完全なままでは、細胞が有糸分裂に進まないようにする。
ヒトの大部分のセントロメア(centromere)は、染色体の中央付近にあり、染色体の結合の中心になっているヌクレオソーム(染色体の基本構成単位)で、ヒストン(ヒストンはDNAに結合するタンパク質の大部分を占め、ヒストンとDNAの重量比はほぼ1:1)からなる芯にDNAが2回巻き付いた部位にある。ヒトのセントロメアは、染色体の長腕と短腕が交差する部位、染色体のほぼ中央に位置することからこの名が付けられた。
典型的な分裂期染色体は、高度に凝縮している。間期にDNAが2倍になるので、分裂期染色体には、2本の全く同じ娘DNA分子が含まれる。染色体が2倍になると、細胞は有糸分裂、つまり核が分裂するM期へ進行する。M期の有糸分裂では、複製で倍加した染色体が凝縮し、遺伝子の発現が殆ど止まり、核膜が壊れて微小管などの細胞骨格(タンパク線維系)からなる有糸分裂紡錘体が形成される。この有糸分裂時には、細胞周期調節制御系は、染色体が2個の娘細胞に分離する前に、有糸分裂紡錘体からなる装置に正しく接着しているかどうかを確認する。
通常の細胞分裂では、凝縮した個々の倍加した母方染色体と父方染色体が、赤道面に2本ずつ並び、その中央部のセントロメアに紡錘体から伸びた微小管が結合して、その一対ごとの姉妹染色体は、1セットずつ細胞の両端へ引かれ分配される。細胞が分裂する直前に分離した染色体セットの周りに核膜が形成される。M期の最終段階で、細胞が分裂し2個の娘細胞ができる。
間期の間、細胞は遺伝子の転写とタンパク合成を続けるため大きさが増大する。またS期と共にG1期とG2期は、細胞の成長と細胞小器官の複製のために必要な時間である。もしこの間期にDNA複製をする時間しかなかったとすれば、細胞は分裂前に2倍に成長できないので、細胞分裂の度に小さくなってしまう。
確実にすべてのDNAと細胞小器官を複製し、正しく分裂が行なわれるためには、細胞は、細胞周期制御系という複雑な調節タンパクのネットワークを備えている。このネットワークが、DNA複製や有糸分裂などの細胞周期の事象を決められた順番で行うため、各々の行程の完了を待って、次の過程を開始していく。このため制御系自体も、進行中の過程からフィードバック調節を受けている。
核が分裂する前に、核DNAの複製が完了するように、またDNA合成が遅れれば、有糸分裂も細胞分裂も遅れるため、このような細胞周期の過程に、中断・遅延などの、異常が生じないように調節されている。つまり、M期の前にS期が完了していなければならない。DNA合成が遅れたり、止まったりすれば、有糸分裂と細胞分裂を遅らせる必要がある。DNAが損傷したら、修復のためのDNA複製の開始と、それが完了するまで、細胞がM期に入る前に、G1期かS期かG2期で止まる必要がある。細胞周期制御系は、ある移行点で周期を止めるため、チェックポイントとも呼ばれる分子のブレーキを利用して、これらすべてを成し遂げている。こうした制御系は、細胞の準備が順序よく整わないと次の過程に進まないようにする。
目次へ
減数分裂であれ、通常の細胞分裂であれ、真核細胞が分裂する前には、まず、すべての染色体が倍加する。倍加した染色体の2つのコピーは、最初、全長にわたって密着したままなので、各コピーを姉妹染色分体と呼ぶ。しかし、この倍加した染色体が、その後にたどる過程は、減数分列と通常の細胞分裂とで異なる。通常の細胞分裂では、倍加した染色体は、中期赤道面に一列に並び、分列が進むと、姉妹染色分体は離れて2個の娘細胞のどちらかに渡される。
複製のための染色体を構成するには、ある程度の準備が必要である。真核細胞では,間期の初期、G1期に複製を開始する核染色体の部位に、タンパク質を集めDNA複製へ向かう。この複製拠点と呼ばれる核酸配列が、DNA合成を制御し実行する、タンパク質とタンパク複合体の着地点として働く。
二倍体細胞の減数分裂では、2セット分の全染色体が最終的には半分になり、染色体が1セットあるだけの配偶子が作られる。それは、最初に起こる染色体倍加の回数は、通常の細胞分裂と同じく1回であるが、その後DNA複製を行なうことなく連続して2回の細胞分裂が起こり、最終的に細胞内の染色体は半減する。第1分裂では、倍加した父方相同性染色体と母方相同性染色体の対合を分離し、第2分裂では、倍加した父方相同性染色体と母方相同性染色体同士で姉妹染色体が分離し、2回目で、ようやく遺伝子的に同一でない一倍体細胞の配偶子が作られる。
一見すると、減数分裂は通常の細胞分裂を少し変えただけのように見える。しかもDNA複製(S期;DNA複製や倍加した相同染色体の分離など重要な過程)を省略すれば、二倍体細胞は細胞分裂を1回することで2個の一倍体細胞を作れるはずだ。しかし、生物40億年の進化の歴史は、当然、無駄のない合理的な仕組みを作り上げていた。
減数分裂は、卵巣や精巣にある特定の二倍体生殖系列細胞で起こる。これらの生殖系列細胞も体細胞と同様で二倍体で、染色体が2コピーずつあり、一方が父方相同性染色体で、もう一方は母方相同性染色体である。減数分裂の最初の段階で、これらの染色体は倍加し、2倍となった染色体同士は、通常の体細胞の分裂同様に、互いに接着し対合(pairing)する。それ以後の段階が、減数分裂独自であり、しかもそれが、複雑に進化した真核生物すべてが実現した到達点であった。
この対合という過程のおかげで、相同染色体は、その後の2回の細胞分裂に際しても正確に分離し、最終的には配偶子それぞれに完全な一倍体の染色体セットが出来上がる。
減数分裂では、2つの姉妹染色分体を分離する必要があるため、分裂機構に特別な仕組みを備えた。減数分裂によってできる4つの一倍体細胞それぞれが確実に、姉妹染色分体から1本ずつ受け取るには、生殖系列細胞が父方と母方の相同染色体(homologous chromosome;króuməsòum)と対応できなければならない。倍加した相同染色体が、父方と母方同士で対合してから中期赤道面に並ぶことで、この対応が可能になった。
この倍加した相同染色体同士が対合してから、4本の姉妹染色体が赤道面に並ぶ構造を二価染色体(bivalent;baivéil nt;chromosome)という。この構造では、4本の姉妹染色体すべてが密着し、細胞分裂の準備が整うまで、この状態が保たれる。そして、減数第一分裂で父方相同染色体と母方相同染色体が、双方にある減数分裂紡錘体に接着し分離し、第2分裂では、姉妹染色分体が、同様に減数分裂紡錘体に接着し分離する。
対合に際して、相同染色体同士と2種類の姉妹染色体同士が、互いを識別する仕組みは完全には分かっていない。多くに生物では、最初の結合現象で、相同染色体の各所にある相補的な母方と父方のDNA塩基配列同士が相互作用をする。また4本の染色分体からなる二価染色体は、一度対合すると非常に安定する。二価染色体は、減数第一分裂の長い前期に作られて、そのまま維持される。一部の生物では、この段階が何年も続くことがある。
目次へ
有性生殖する生物では、母方染色体と父方染色体が対を作る際に、「相同組換え(homologous recombination)」という過程が起こる。この過程では、同一もしくは非常に似た塩基配列の間で遺伝情報の交換が起こる。
損傷して遺伝情報が失われてしまった染色体を修復するのに、相同組換えの仕組みが使われる。この種の修復では、無傷のDNA二重らせんの遺伝情報を使って、損傷した、新たに倍加した相同染色体に正しい塩基配列を復元させる。この修復機構は、どのDNA二重らせんにも遺伝情報が二重に存在することを利用している。たとえ一方の鎖が壊れても、相補鎖に残っている情報を基に修復する。
しかし、二重らせんの両方の鎖が同時に壊れたらどうなるだろうか。細胞の複製装置は、精度が高く校正機能もあり、コピーの誤りを防止する機能もある。それでも稀に誤りが起こる。複製装置がコピーを誤ると、間違って対合したヌクレオチド(誤対合)が、必ず生じ、これが修復されないと、次のDNA複製の際に、2個のDNA分子のうちの1個に永続的な変異として受け継がれる。誤対合修復タンパク複合体は、DNAの誤対合を識別し、DNA鎖の誤りの部分を切り取って除き、その部分を再合成して埋める。この修復機構で、元通りの正しい配列となる。
細胞には、このような複製の誤りを修正するDNA誤対合修復(DNA mismatch repair)という予備の機構が備わっている。複製装置が誤るのは、107塩基に1個の割合であるが、DNA誤対合修復がその都度働き99%修正するので、全体としては109塩基に1個の誤りまでとなり、その精度は極めて高い。
放射線照射・複製中の事故など、様々な化学反応によってDNAの主鎖が切れて、二本鎖切断が生じる。これらの切断が特に危険なのは、染色体が直ぐに小さな断片となり分解してしまうことである。当然、遺伝子は失われる。この種の損傷の修復が特に難しい。どの染色体も、それぞれ特有の情報を含むが、二本鎖切断が生じると切れた部分が離れてしまい、その失われた情報を復元しようとしても、細胞には予備のコピーがないことになる。
重大な結果を招きかねないこの二本鎖切断を修復するために、2つの基本戦略を進化させてきた。1つはDNA切断が離れて遠くに行かない前に、切れた末端を急いで繋ぐ方法である。この修復機構は非相同末端連結(non-homologous end joining)という。多くの型の細胞で働くが、切断された末端をヌクレアーゼなどの特殊な酵素が、損傷したヌクレオチドをDNA鎖から切り除き、修復DNAポリメラーゼがギャップを埋めた後をきれいに整え、その損傷鎖に残る糖-リン酸主鎖の切れ目(ニック)をDNA修復酵素群の一つDNAリガーゼがこれを再び繋ぐ。このやっつけ仕事では、切断は素速く修復されるが代償も払う。末端を繋ぐ時、連結部位のヌクレオチドのいくつかが失われることが多いからだ。
この緊急の機構で修復しても、多くの場合、それ以上の問題は生じないが、「相同組換え」のように元通りに戻らないため、遺伝子が働かなくなるなど重大な影響が出てしまう。そのため細胞は、これに代わる誤りの少ない二本鎖切断修復方法も備えている。この機構を「相同組換え」と呼ぶ。
それと同様な過程が、減数分裂第1分裂の長い前期に相同染色体の対合にも起こる。しかし減数分裂では、相同組換えが、二価染色体を作る倍加した相同染色体の姉妹染色分体同士の間ではなく、姉妹関係にない染色分体の間で起こる。そのため母方と父方の相同染色体の相同部分同士が、物理的に交換される。この複雑で多段階の過程を交差(crossing-over)という。
減数分裂第1分裂の前期が終わるまで、この相同組換えを担うタンパク複合体が、まず2本の染色体の母方か父方の一方のDNA二本鎖に、ヌクレアーゼが5‘末端を分解し切れ目を入れた後、更なるDNA合成と、その後のDNA連結で、もう一方の染色体との鎖の交換を誘導する。この二価染色体における非姉妹染色分体の間で、DNA領域の交換が行なわれるため、各染色分体には、相手の非姉妹染色分体に由来するDNA領域が含まれることになる。減数分裂で染色体の交差ができる過程の多くの段階は、体細胞で起こるDNA二本鎖切断の修復過程と似ている。
交差はシナプトネマ構造の形成により誘導される。この精巧なタンパク複合体は、倍加した相同染色体が対を作ると、できた二価染色体を安定させ、姉妹関係にない染色体同士の間で、DNA鎖の交換が起こりやすいように相同染色体を並列させる。倍加した相同染色体を作る染色分体は、それぞれ非常に長いDNA二重らせんができているが、同じ二価染色体を作る別の相同染色体の染色分体のどちらか、あるいは両方と交差を形成する。シナプトネマ構造は、一本の染色体上で起こる交差と交差の間隔を、一定以上あける役目もしている。
シナプトネマ構造は、第1分裂前期が終わるまでに解体するため、相同染色体はほぼ全長にわたって互いに離れていく。しかしその後も、少なくとも1個のキアズマによって二価染色体は、まだ1つに保たれている。光学顕微鏡で観察すると、バッタのキアズマ結合は3本あった。非姉妹染色体同士が交差する3個の部位である。二価染色体の殆どの場合、2個以上のキアズマができ、相同染色体間では複数の交差が起きている。減数分裂により卵細胞になるヒトの卵母細胞(らんぼさいぼう)では、二価染色体1つにつき平均して2~3か所の交差ができている。
減数分裂で起こる交差は、有性生殖をする種で生じる遺伝的変動の主な発生源である。交差は、配偶子に入る各染色体の遺伝的構成を切り混ぜるので、それ以前にはない対立遺伝子の組み合わせを持った個体を作り出す。
交差は減数分裂にとってもう1つの重要な役割を果たしている。交差でできるキアズマ(chiasma;交叉を意味するギリシア語)により、二価染色体が維持され、相同染色体は、第1分裂前期の結合したままの状態に保たれ、第1分裂時に母方と父方の相同染色体が互いに正確に分離することになる。
目次へ
大部分の生物では、2本に倍加した相同染色体が減数分裂で正しく分離して、別々の娘細胞の核に入る際に、交差が必要である。交差によってできるキアズマは、第1分裂後期に紡錘体によって引き離されるまで、母方と父方の相同染色体を固く結合させる。第1分裂後期の前から紡錘体は、倍加した相同染色体を両極に引き寄せるが、キアズマはそれに耐えて、二価染色体を中期赤道にとどめて安定させる。
キアズマは、母方と父方の相同染色体を固く結合させるが、減数分裂第1分裂では、コヒ-シンというタンパク質によって姉妹染色体の腕が全長にわたって接着されている。それが後期になると、コヒ-シンは急速に分解し、腕が離れて組換えを終えた相同染色体が引き離されるようになる。もし相同染色体の腕が離れないと、倍加した母方と父方の相同染色体は、交換した相同なDNA領域によって互いにつながったままになる。
そして相同染色体が分離した後も、セントロメアだけにとどまったコヒーシンは、第2分裂後期まで姉妹染色体の接着を保つ。キアズマが分解され、それに合わせて相同染色体が分離されて、後期の1回目の細胞分裂が完了すると、2個の二倍体細胞それぞれに姉妹染色体が残る。
減数分裂の第1分裂を終えると、細胞は直ぐに第2分裂に入るが、ここでも細胞内の両端に紡錘体が形成され、それぞれの姉妹染色分体対にある動原体が、通常の細胞分裂と同様に、互いに反対の極に向う動原体微小管に接着する。第2分裂の後期になると、セントロメアで姉妹染色分体を結合させていた減数分裂特有のコヒーシンが分解され、姉妹染色分体は別々の極に引き離され別々の娘細胞となる。
ヒトでも同じ両親から生まれた兄弟姉妹は、一卵性双生児でない限り、前述するように遺伝的同一ではない。この遺伝的差異は、精子と卵が出合うより前の減数分裂第1分裂の段階で、遺伝子情報のランダムな再編成が2回、しかも違う仕組みで起こっているからだ。
最初が第1分裂の最中に、母方と父方の相同染色体が混ぜ合わせられランダムに分配される(相同組換え)。それぞれの娘細胞が相同染色体を1コピーだけ受け取るように巧みに配置されるが、母方と父方の相同染色体のどちらになるかは偶然である。そのため、各配偶子に含まれる1セットのうち何本かは、母方由来でその他が父方由来となる。このランダムに混在するようになるのも、それぞれの二価染色体が、第1分裂中期に赤道面に並び、姉妹染色分体の動原体微小管は、揃って同じ方向に両極にある紡錘体に引き寄せられ、倍加した相同染色体同士が分離される。
その時、それ以前から赤道面に2組の姉妹染色体がならび、交差するキアズマの働きで、倍加した母方と父方の相同染色体と固く結合しているため、赤道面を越えてランダムに母方染色体と父方染色体が再分配される。
こうして母方と父方の相同染色体のランダムな再配分だけでも、1個体が作りだせる配偶子の遺伝的な組み合わせは、原理的には2n(nは一倍体の染色数)通りになる。ヒトの場合、減数分裂で起こる母方と父方の相同染色体のランダムは再配分だけで223通りできる。しかし各個人が生みだせる配偶子の種類は、実際にはそれを遥かに超える。それは減数分裂時の交差が、第2のランダムな遺伝的再配分が起こるからだ。
ヒトの相同性染色体1対につき平均して2~3個の交差が起こり、母方と父方の対立遺伝子を新たに組み合わせて新しい染色体を作る。1本の染色体全体のどこで交差が起こるかはランダムなので、1回の減数分裂で新しい4つの染色体セットが生まれることになる。
母方と父方の染色体のランダムな再配分に加え、減数分裂第1分裂前期に起こるキアズマ交差によって遺伝情報の混和が生じ、1つの個体が作り出す配偶子の遺伝的多様性は無限となり、遺伝的に全く異なる2人から作られる配偶子が融合して子ができるとなると、ヒトの多様性は驚くほどとなり、親子・兄弟でも個人差が生じるのは当然である。
目次へ