| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生
花粉の形成と構造 雌ずい群 花粉管の先端成長
| 目次 |
| 1)力学的仮説(花粉管を雌ずい内に導くもの) |
| 2)化学走性仮説 |
| 3)重複受精における精細胞の挙動は3段階に分けられる |
| 4)胚珠原基 |
 受精するまで、花粉管は胚珠の珠孔までの道のりをたどらなければならない。さらに、最初に珠孔に到着するためには、花粉管相互の競争があり、それに勝って初めて卵細胞と受精がかなう。
受精するまで、花粉管は胚珠の珠孔までの道のりをたどらなければならない。さらに、最初に珠孔に到着するためには、花粉管相互の競争があり、それに勝って初めて卵細胞と受精がかなう。花粉管を取り込む雌ずいの組織も、花粉管相互の競争に関わっている。雌ずい組織内の因子が、花粉管の伸長や珠孔への到達を促してもいる。
胚珠への花粉管の伸長を説明するのに、力学的仮説mechanical hypothesisと化学的走性仮説chemotropic hypothesisの2つが提唱されている。
力学的仮説では、雌ずいの構造自体が花粉管の通り道であり、この通路が、胚珠へ向かう狭い伝達組織transmitting tractへ導く。胚珠に至る過程で、花粉管は伝達組織の細胞外基質の成分と直接接触する。伝達組織の細胞外基質は、細胞壁タンパク質群からなる複雑な混合物である。主に細胞壁には、局在しているアラビノガラクタンタンパク質(AGP)や、プロリンに富む糖タンパク質(PRP)、およびヒドロキシプロリン(Hyp)と糖タンパク質(HRGP)が含まれている。
AGPには、一般的に、ヒドロキシプロリンに富むコアタンパク質core proteinに、ガラクトース(Gal)とL-アラビノース(L-Ara)に富むアラビノガラクタン(AG)糖鎖が結合している。コアタンパク質の種類が多いのも、糖鎖の構造が複雑で多様性に富んでいるからである。AGPは植物の分化・成長に関わる多彩な生理機能を持つ情報因子として注目されており、多様な糖鎖構造の解明と生理機能の相関性についての研究も進展しつつある。
これらのタンパク質は、花粉管が定位置を保つため接着分子となり、また花柱内を伸長するための牽引力を与えている。また細胞外基質は、花粉管の代謝活性を支えるため、栄養素も供給していると見られている
「細胞外基質extracellular matrix」は、動物では細胞の外、つまり細胞膜の外にあるタンパク質や糖などの巨大分子からなる構造体である。植物でも、動物と同様、細胞膜の外側という意味合いから、この用語が使われるが、植物の細胞外基質の大部分を占める細胞壁は、細胞の一部として重要な機能を果たしている。その点、用語の使い方に矛盾が生じている。
細胞膜plasma membrane(mémbreɪn;膜)は、細胞質と外界の境界となる流動的な膜である。動物や酵母・細菌などの細胞膜も同様である。
細胞質cytoplasm(sάɪṭəpl`æzm)は、その細胞膜に取り囲まれた可溶性でコロイド状のサイトゾルcytosolと、核以外のすべての細胞小器官および細胞骨格cytoskeltonを含む。また、核膜に包まれた核質は細胞質には含まれない。
動物細胞には細胞壁がないため、動物胚の細胞は、胚の中を移動することができる。そのため動物の発生過程の各組織や器官は、個体中の別の部分に由来する細胞を含むことがある。
真核生物が、各種の構成成分を細胞内に一定の秩序を保って配置したり、多様な形を構成したり、外部環境と機械的な相互作用をしたり、調和のとれた運動を継続できたりするのは、すべて細胞骨格のおかげで、細胞骨格により細胞は形を保ち、中の構成成分を配置したり、線維芽細胞などの細胞がはい回れるようにしたりする。
動物の線維芽細胞は、主に線維状タンパクのコラーゲンが引っ張り強度を担う結合組織connective tissueの中に住み、細胞外マトリクスを作る。動物の組織内で細胞分裂が起こる時に、その細胞の解離と接着のサイクルに応じて、隣接細胞や細胞外マトリックとの繋がり方が再編され、細胞分裂で新しく生じた細胞が組織内で適切な位置を占めるようになるのだろう。
動物の結合組織は、極めて多様で、腱や皮膚の表皮のように強靭でありながら柔軟なものも、骨のように固く緻密なもの、軟骨のように弾力性に富み緩衝能力が高いもの、目のガラス体のように柔らかく透明なものなどがある。骨などの結合組織では細胞外マトリックが容積の大部分を占め、強度を全面的に支えている。
細胞骨格は、中間径フィラメント・微小管・アクチンフィラメントと呼ばれる3種類のタンパク線維で構築されている。その3種類のタンパク線維は、それぞれが別種のタンパク質が構成単位となっているため、機械的な特性が異なる。
いずれも何千もの構成単位が集まってタンパク質の糸を作り、それが細胞全体を横断していることもある。
細胞骨格が細胞質に広がり、そのタンパク線維網目構造が、細胞質空間を支えるからで、細胞骨格成分は細菌にも多少あるが、特に細胞壁をもたない動物細胞には不可欠で、大きな構造と複雑な真核細胞には特に重要である。
細胞骨格は、人体の骨格とは異なり、極めて動的な構造体であり、細胞が環境に応答して形を変えたり、分裂する際には、それに合わせて常時作りに直されたりしている。細胞骨格は、細胞の骨であり、筋肉でもあり、細胞が動き回る時や、筋細胞の収縮、胚発生時に起きる細胞の変形などの大規模な動きにも関わっている。細胞骨格が存在しなければ、傷は治らず、筋肉は収縮できず、精子は卵に泳ぎ着かない。
中間径フィラメントは、ロープ状の線維で直径は10nmで、これを構成するのは、糸状の中間径フィラメントタンパクである。一群の線維状タンパクが、長くねじれたロープをたくさんより合わせたようなロープ構造となって束になり、機械的刺激を受けると変形はするが断裂はしない、非常に柔軟で引っ張り強度に優れている。殆どの動物細胞の細胞質に存在し、通常は核の周辺から細胞の周縁部まで網目状に広がっている。通常、細胞同士を細胞膜で繋ぐデスモソームdesmosomes(細胞間結合装置の1つ、例えば上皮細胞の間に形成される、隣り合った細胞のケラチンフィラメントを繋ぎ、引っ張り強度に貢献する)と呼ばれる細胞間結合のところで細胞膜にくっついている。中間径フィラメントは、真核細胞の核内にもあり、核膜を裏打ちして補強する細かい網目模様が、中間径フィラメントで、核膜内膜の直下にあり、「核ラミナ」と呼ばれている。
核は、核外膜と核内膜と呼ばれる、機能分化した、性質の全く異なる2層の脂質二重膜から形成されている。核外膜は脂質成分が多く、小胞体膜と連続して繋がっている。小胞体は、膜でできた袋や管からなるひとつながりの構造で、細胞全体に広がっていることが多い。細胞内での膜の新たな合成は、主として小胞体で行われる。
一方、核内膜は脂質というよりむしろタンパク質に富む構造で、動物細胞では核ラミナと呼ばれるメッシュ状の構造に裏打ちされて、クロマチンと物理的に繋がっている。2,000年頃から、核内膜因子群の変異が早老症などの多くの遺伝子疾患の原因になることが明らかにされ、転写や複製などの遺伝子機能の制御に核膜が重要な役割を果たすと考えられている。
核膜のもう1つの特徴は、外膜と内膜を貫通する核膜孔と呼ばれる複合体(総重量100MDa)、超分子構造体が存在し、核と細胞質間の連絡は、この構造体を通して行われている。
他にも、細胞質を横断して細胞に構造強度与えているもの、上皮組織で細胞質を貫いて細胞間結合同士を繋ぎ、機械刺激を分配するものもある。中間径フィラメントの非常に柔軟で引っ張り強度に優れた構造と集合により、細胞が強靭になり断裂から守られている。
一方、植物細胞の細胞骨格には、その張力に耐えられる中間径フィラメントがないので、本来引っ張り強度がない。そのため細胞の外壁が不可欠になる。植物細胞の細胞膜の周囲は、セルロース性の細胞壁cell wallに取り囲まれている。その隣り合う細胞壁は、中葉middle lamellaと呼ばれる薄い層を介して接着されている。そのため細胞の移動は起こらない。そのため植物の発生は、細胞分裂と細胞成長のパターンに完全に依存している。
典型的な中葉は、酸性多糖類(ペクチン類)に富み、それらがアミノ酸の一種であるヒドロキシプロリンhydroxyprolineに富む糖タンパク質(HRGP)と複合体を作ることがある。中葉の起源は、細胞質分裂cytokinesis時に形成される隔壁の前駆体である細胞板と見られている。中葉は、細胞が拡大する過程でも、素材が補給され続けられ、その主要な機能は、細胞間における柔軟な接着層の形成である。
植物の細胞壁には、一次細胞壁primary cell wallと二次細胞壁secondary cell wallの2種類がある。一次細胞壁は、若く成長をし続けている細胞に見られる。一般的には1μm未満とごく薄い。
二次細胞壁は、細胞の成長がほぼ終了した後に、一次細胞壁の内側に沈着する。それは一次細胞壁よりも分厚く強固である。その細胞壁の強靭さと耐久性は、かたい膠(にかわ)状の物質であるリグニンligninによる。二次細胞壁がリグニン化する進化により、植物は地面に対して垂直に屹立する強度を獲得し、陸上に進出することが可能になった。同じ陸上植物でありながらコケ類が、数cmしか成長できないのも、リグニン化した細胞壁を持たないからである。
目次へ
近年、とりわけ複雑な花器官の特定の細胞を研究対象として解析する様々な技術が開発され、受精の関わる分子とその立体的構造が急速に明らかになりつつある。解明された物質や細胞の動態から、生殖細胞の形成・花粉管誘導・重複受精といった現象について植物の戦略が解明されてきている。
特に、様々なペプチド(1つのアミノ酸残基と次のアミノ酸残基の間の繋がりは、アミド結合またはペプチド結合と呼ばれる)によるシグナル分子を介した多様な細胞間の相互におけるコミュニケーションの詳細が、徐々に解読されている。
化学走性仮説chemotropic hypothesisの屈化性とは、上下関係の階層性をもつ分子群の刺激に反応して起こる屈性で、植物の根・花粉管が、ある濃度の糖類やタンパク質に反応して、一定方向に曲がる性質を指す。
花粉管の先端を胚珠に向かって成長させ、最終地点の珠孔まで誘導するという、諸々の仮説の積み重ねが、その実相を明らかにしてきている。
花粉から伸長した花粉管は、雌ずいの各々の組織から、多段階のガイダンスを受け取る。受精に失敗した胚珠は、残りの助細胞を使い2本目の花粉管を積極的に誘引し受精が回復する。
例えばシロイ ヌナズナでは、
1)柱頭
2)伝達組織
3)合点chalaza
4)珠柄(胚珠の柄)をよじのぼる段階
5)珠孔(胚嚢へつうじるトンネル)の入り口に入る段階
と言った5段階のガイダンスが観察されている。
しかも、1つの胚珠に複数の花粉管が向かわないようにする反発作用も提唱されている。
化学走性仮説では、シグナル分子が順々に花粉管の先端を刺激することで、花粉管を胚珠へ向かわせ、雌ずいで発現される分子のいくつかが、花粉管を誘引する機能を持つことが発見されている。
 ユリでは、低分子の分泌性タンパク質であるstigma/style cysteine-richadhesin(SCA)が同定されている。SCAは、脂質輸送タンパク質lipid
transfer proteinファミリーに属するタンパク質であり、中空の花柱に並んだ伝達組織(雌ずいの中央を貫いている花粉管の通り道にあたる組織)の表皮から分泌されて、その伝達組織に沿った花粉管pollen tubeの伸長と接着に関わる。
ユリでは、低分子の分泌性タンパク質であるstigma/style cysteine-richadhesin(SCA)が同定されている。SCAは、脂質輸送タンパク質lipid
transfer proteinファミリーに属するタンパク質であり、中空の花柱に並んだ伝達組織(雌ずいの中央を貫いている花粉管の通り道にあたる組織)の表皮から分泌されて、その伝達組織に沿った花粉管pollen tubeの伸長と接着に関わる。別の低分子の分泌性タンパク質であるblue copper protein(ブルー銅タンパク質)のファイトシアニンファミリーに属するケモシアニンchemocyaninも、方向性を制御する合図として働く。
ユリの柱頭stigmaから見い出されたペプチドであるケモシアニンは、花粉管化学屈性を誘導する。花粉管との接着・方向性の制御・伸長の促進、そして、近年見い出された受精能の制御などを担っているようだ。
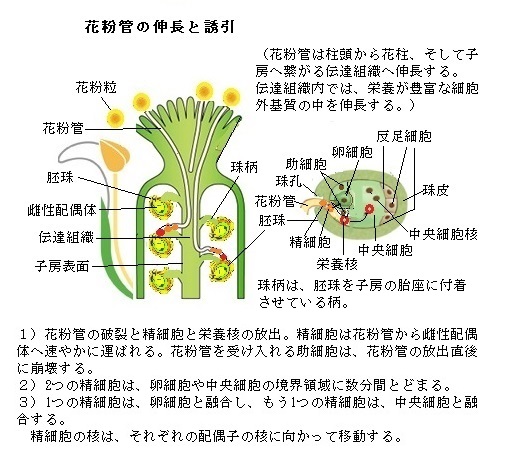
花粉管が、雌ずいの基部の子房に達すると、胚珠組織内の胚嚢によるガイダンスの領域に入る。既に140年ほど前、培地に取り出した胚珠に、花粉管が向かうことが観察されていた。花粉管を胚珠に導く誘引物質が存在するのではないかと考えられ探索されてきた。しかし、その分子は長らく不明のままであった。
シロイヌナズナを用いた分子遺伝学的な研究が進められている。花粉管伸長の初期段階は、伝達組織内の胞子体細胞により制御されるが、雌ずい組織を通り抜けたシロイヌナズナの花粉管を用いた遺伝子的解析によって、雌ずい組織の通過によって、花粉管の遺伝子発現が著しく変わることが示されている。花粉管は柱頭から子房へ向かう際、花柱を通り抜けるが、花柱組織は伝達組織や子房に至る径路を提供するだけではなく、花粉管が胚嚢の助細胞から産出される誘引シグナルを受容できるように花粉管の状態を整えている。
胚嚢の半分ほどが胚珠から突出し、卵細胞がむき出しとなっているアゼトウガラシ科のトレニアという植物を用いた研究により花粉管の誘引物質の研究が進展した。トレニアにおいて卵細胞に隣接する2つの助細胞が拡散性のシグナルにより花粉管を誘引することが観察されている。しかも、その誘引物質に花粉管が応答するためには、花柱の役割が重要であることが明らかになった。
トレニアは、ラベンダーやライラックと同様、シソ目の一種であるが、胚嚢から放出された花粉管誘導物質に花粉管が応答し始めるには、花柱の働きが重要な役割を果たしていた。
一般的に「トレニア」と呼ばれているのは、トレニア・フルニエリTorenia fournieriで、市販の種を春に播けば、暑い夏から次々と多くの花を咲かせ、晩秋まで楽しめる。耐陰性のある育てやすい植物で、しかも、前年に栽培していた場所に、こぼれた種が発芽して増え続ける。
殆どの被子植物では、胚嚢を胚珠の胞子体組織から取り外すことは不可能である。
トレニアなどのいくつかの植物種では、胚嚢は珠孔から飛び出し、珠柄に沿って成長をするから、卵細胞・2つの助細胞、それに中央細胞の約半分が胚珠の外側に出ている。トレニアの胚珠を胎座から切り離すと、剥き出しの胚嚢が直接培地に置かれることになる。単離培養したトレニアの胚珠の領域から飛び出している胚嚢の珠孔端に到着した花粉管の顕微鏡像から、雌雄配偶体の2つの助細胞による化学的シグナルも、花粉管を胚珠へ導くのに重要な役割を担うことが知られた。
この実験により、トレニアの胚珠と、栄養を含む培地上で発芽させた花粉管と、共培養しても、その花粉管は胚珠に向かって伸長しない、ところが、トレニアの花粉粒を生きた柱頭上で発芽させ、花柱の切り口から出てきた花粉管を使うと、花粉管は胚嚢の珠孔端に向かう。花粉管は、雌の胞子体組織と相互作用することで、雌性配偶体からの合図に応答できる状態に整えられ、珠孔へ向かって伸長ができるようになる。
胚嚢の特定の細胞をレーザーによって直接破壊する実験によって、トレニアの花粉管誘引物質の供給源となる細胞が特定された。卵細胞や中央細胞ではなく、助細胞が死滅した時にのみ、花粉管は胚珠に向かって伸長できなくなった。
さらに顕微鏡により取り出した助細胞において発現している遺伝子を解析した結果、助細胞からはシステインcysteine(硫黄を含むアミノ酸の1つ)に富む分泌性と考えられる多くの種類のポリペプチド群が発現していることが明らかになった。そして、助細胞において特異的に強く発現する、トレニアの花粉管に対する化学物質として、LURE(lˈʊɚ)タンパク質群と呼ばれるシステインに富むポリペプチド群が、花粉管の誘引物質として同定された。
LUREタンパク質群は、動物と植物で発見されたている抗菌性タンパク質である「ディフェンシンdefensin」に似たタンパク質群である。しかもトレニアの様々なLUREタンパク質群は、シロイヌナズナでも同定されており、種特異的に作用するようで、シトイヌナズナのLUREを発現させたトレニアの胚珠は、シロイヌナズナの花粉管を選択的に誘引する。
これらは、花粉管を引き寄せる性質からLUREと名づけられた。Lure(lˈʊɚ)とは、「ルアー釣り」の原義に通じ、「誘惑するもの」「おとり」である。その高い発現量は、花粉管の長い誘引距離を、十分にガイダンスできると考えられている。
目次へ
シロイヌナズナの精細胞の挙動は、3段階に分けられる。
最初に花粉管が破裂する。
花粉管を受け入れる助細胞が崩壊するのと同時か、その直前、数秒以内に精細胞が胚嚢内に運ばれる。
次に、放出された2つの精細胞は、卵細胞と中央細胞の境界領域に約7分間とどまる。
最後に、1つの精細胞は卵細胞の核と融合し、もう1つの精細胞は中央細胞の核と融合することで、重複授精が完了する。
ただ、動物とは異なり、植物は系統により、それぞれ異なる生活環を持つ。種子植物の配偶体における精子と卵の形成や、受精の際の動態は、種子植物の種によっても様々である。被子植物が行う重複受精double fertilizationでは、2つの精子が作られ、その内の1つのみが卵と受精する。もう一つの精子は、雌性配偶体の中央細胞の2つの核と融合し、被子植物の種子の貯蔵組織となる核相が3N、即ち染色体セットを3つ持つ胚乳を生じさせる。ただし、被子植物の中には、さらに核相の高い胚乳を生ずるものもある。
裸子植物は、重複受精を行わないため、裸子植物の種子では、1N(単相)の配偶体の組織貯蔵組織として働く。
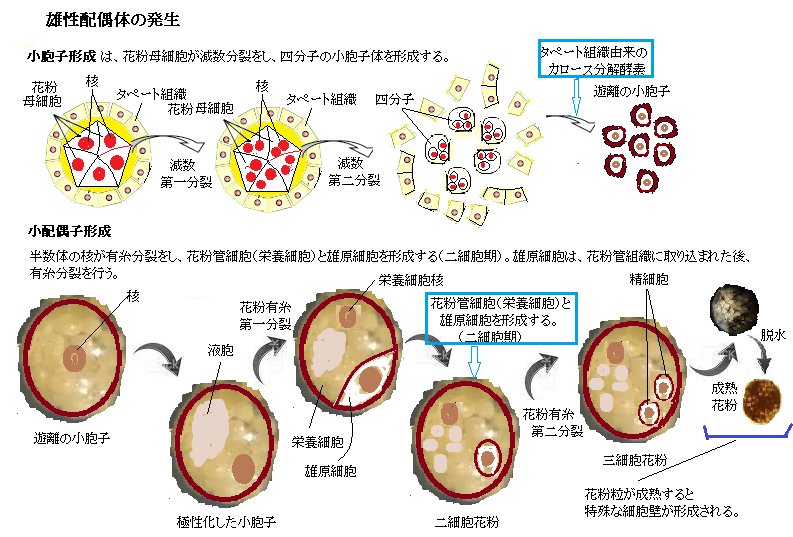 植物は、動物のように減数分裂によって、単細胞の配偶子を直接作るのではなく、減数分裂はするが、それにより単独で栄養成長する胞子を生み出す。その1Nの胞子が、有糸分裂をして多細胞体の雄性配偶体と雌性配偶体を形成する。配偶子となる精子と卵は、それぞれの配偶体の有糸分裂で形成される。精子と卵が受精されてできた2N(複相)の接合子は、多細胞体の種子を形成して、やがて発芽して成熟した胞子体に成長し、再び減数分裂を経て配偶体を形成する。
植物は、動物のように減数分裂によって、単細胞の配偶子を直接作るのではなく、減数分裂はするが、それにより単独で栄養成長する胞子を生み出す。その1Nの胞子が、有糸分裂をして多細胞体の雄性配偶体と雌性配偶体を形成する。配偶子となる精子と卵は、それぞれの配偶体の有糸分裂で形成される。精子と卵が受精されてできた2N(複相)の接合子は、多細胞体の種子を形成して、やがて発芽して成熟した胞子体に成長し、再び減数分裂を経て配偶体を形成する。重複受精には、未だ多くの疑問が残っている。例えば、花粉管の破裂はどのように制御されているのか?
 あるモデルによれば、助細胞の細胞膜上に存在する受容体様キナーゼ(kinaseは、リン酸化を触媒する酵素のことで、タンパク質などの基質にATPに由来するリン酸を付加することにより、その性質を変化させる。)が活性化され、活性酸素種reactive oxygen species(ROS)の生産とカルシウムイオンの取り込みが促進される。外部からヒドロキシルラジカルhydroxyl
radical(活性酸素の中でも最も反応性が高い化合物、過敏反応を通して細胞死の誘導にも寄与する。)を投与すると、カルシウムイオンに依存して花粉管の破裂が誘導されることが知られているので、ヒドロキシルラジカルと高濃度のカルシウムイオンとの組み合わせによって、花粉管の破裂が引き起こされるのかもしれない。
あるモデルによれば、助細胞の細胞膜上に存在する受容体様キナーゼ(kinaseは、リン酸化を触媒する酵素のことで、タンパク質などの基質にATPに由来するリン酸を付加することにより、その性質を変化させる。)が活性化され、活性酸素種reactive oxygen species(ROS)の生産とカルシウムイオンの取り込みが促進される。外部からヒドロキシルラジカルhydroxyl
radical(活性酸素の中でも最も反応性が高い化合物、過敏反応を通して細胞死の誘導にも寄与する。)を投与すると、カルシウムイオンに依存して花粉管の破裂が誘導されることが知られているので、ヒドロキシルラジカルと高濃度のカルシウムイオンとの組み合わせによって、花粉管の破裂が引き起こされるのかもしれない。植物にとっては、ある種の金属は、必要量はごく僅かであっても必須必要な金属であることが少なくない。特に鉄欠乏は、現代の人間にとって、世界中で最も一般的な栄養欠乏であり、カルシウムと並んで不足しがちな栄養素の代表格である。鉄は、その60~70%は血液に含まれるヘモグロビンの成分として、20~30%は肝臓や脾臓、骨髄などに「貯蔵鉄」として蓄えられているほか、筋肉の中や酵素の形でも存在している。
植物体内では、他の物質にキレートchelateされた金属イオンは、道管内に輸送され、蒸散流にのって植物全体に分配され、それぞれの金属イオンは、細胞内の適切な場所に運ばれる。
キレートとは、分子の立体構造によって生じた隙間に金属を挟む姿から「カニのはさみ」を意味する。中心に金属イオンを挟むような形で、キレート分子の2つ以上の配位原子が結合する。配位結合とは、形成する2個の原子の一方のみから、結合電子が供与される結合のことで、その化合物となる配位結合は、多くの錯体にその例がみられる。キレート化により、吸収されにくい養分をアミノ酸や有機物によってカニバサミのようにはさみ込んで、吸収されやすい形に変えたり、反対に有害物質を無害化したりする。
金属を有機物が包み込んだ状態で、吸収されやすい一つの物質のように振る舞う化合物を錯体と言う。血液中のヘモグロビンは鉄を包み込んだ錯体である。
植物では、鉄の大半は葉緑体に見いだされる。鉄は、葉緑体において、電子伝達系や活性酸素種(ROS)除去などに関わるタンパク質の補因子として必要である。ところが、葉緑体自身が主要なROS発生装置である。
鉄・銅などの金属イオンの過剰蓄積は、有害な活性酸素種を産出する。金属イオンをキレートする物質は、この毒性から植物を守り、液胞に金属イオンを輸送する輸送体は、金属濃度を毒性レベル以下に維持するのに重要な働きをする。
その一方では、植物にどのように鉄分が貯蔵されるか分かるにつれ、穀物に含まれる栄養素としての鉄分が、その穀物の評価を高めるようになっている。
 植物の生存と成長は、細胞内・細胞間のエネルギーや物質の流れを制御する同化・異化径路の複雑なネットワークに依存する。乾燥・オゾン・塩・強光・低温・凍結・熱・紫外線や嫌気条件から好気条件への変動など活性酸素種(ROS)が生まれる環境要因により、このネットワークが阻害されると、同化・異化径路の協調性が崩れる。
植物の生存と成長は、細胞内・細胞間のエネルギーや物質の流れを制御する同化・異化径路の複雑なネットワークに依存する。乾燥・オゾン・塩・強光・低温・凍結・熱・紫外線や嫌気条件から好気条件への変動など活性酸素種(ROS)が生まれる環境要因により、このネットワークが阻害されると、同化・異化径路の協調性が崩れる。例えば、代謝酵素の至適温度は、しばしば異なるので、周囲の環境温度が上下すると、同一の、または関連する代謝酵素の径路の中で、その一部の働きが阻害され、その代謝経路に、機能的に連関する部位が破綻し、有毒な副産物が生じかねない中間代謝産物の蓄積が起こることがある。
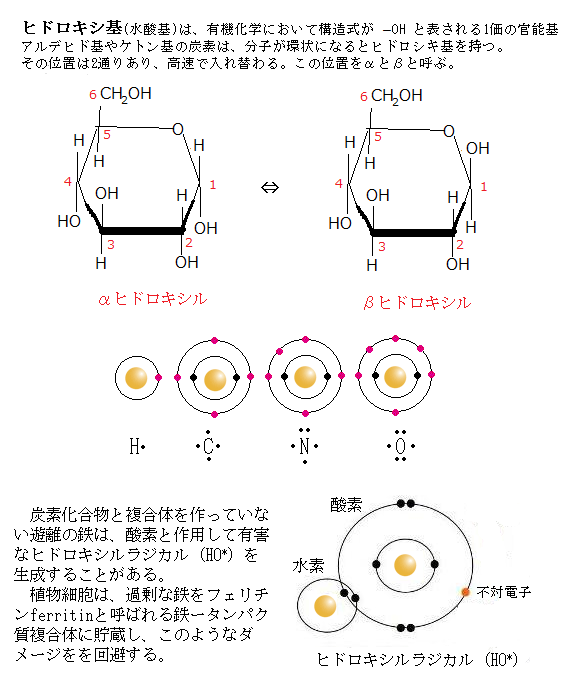
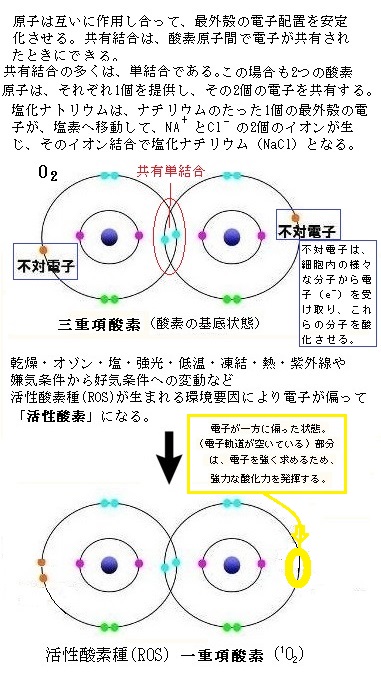
様々なストレスにより共通して生成される、毒性のある中間体として活性酸素種(ROS)がある。ROSでも過酸化水素(H2O2)と一重項酸素(1O2)は、不対電子を持たないが、過酸化水素は不安定で酸素を放出しやすく、生体内では、フェントン反応(下の式)によって、非常に強力な酸化力を持つヒドロキシルラジカル(HO*)を生成しやすい。
Fe2+ + H2O2 ⇒ Fe3+ + OH- + OH*
通常、大気中に存在する酸素は、三重項酸素(O2)と言う基底状態にあり、その原子に光や電子などの粒子を当ててエネルギーを加えると、励起状態に移り一重項酸素になる。
そのエネルギーを放出して、元の安定した三重項酸素になりやすいため、反応性に富む。
通常のROSは、反応性の高い酵素の形態で、電子の最外殻軌道に少なくとも1つの不対電子を持つ。
不対電子unpaired electronとは、原子核のまわりの最外殻軌道に位置する電子が2個の対となって、軌道上に存在する電子の一方が失われて、通常の共有結合を作る共有電子対や非共有電子対に比べ、化学的に非常に不安定であるため、化学反応性に富み、他原子あるいは他分子と容易に反応する。
酸化と還元(reduction)は、共有結合で結ばれた原子間で電子が部分的に偏っている場合にも用いられる。電子の輸送も酸化還元反応(redox reaction)である。電子は、最外殻軌道電子との親和性が相対的に低い分子から、親和性の高い分子へ自発的に受け渡される。それにより、分子及び原子間で、電子を与えたものは酸化され、電子を受け取ったものが還元される。
ROSは、自己触媒的に生体膜の構成成分を酸化し、細胞小器官organelleや細胞膜の分解や、細胞死をもたらす。それぞれのメカニズムは異なるが、殆どの非生物的ストレスは、ROSを産出させる。
各代謝系は、細胞内の特定の部位に区画化されているが、環境ストレスは、この代謝系の区画化も阻害する。酵素活性を阻害する極端な温度、即ち高温化では膜の流動性が増し、低温下では低下する。その膜の流動性の変化により、活性酸素種が生成される。その活性酸素種の酸素に電子が渡され、葉緑体やミトコンドリア内膜のタンパク質複合体間にある分子が壊され、その結果、協調が阻害され、電子伝達系の制御が破綻する。
 典型的な植物細胞は、エネルギー生産を行う2種類の細胞小器官、即ちミトコンドリアと葉緑体を持つ。どちらも外膜と内膜の2重膜でサイトゾルから隔てられ、自前のDNAとリボソームribosomeを持っている。
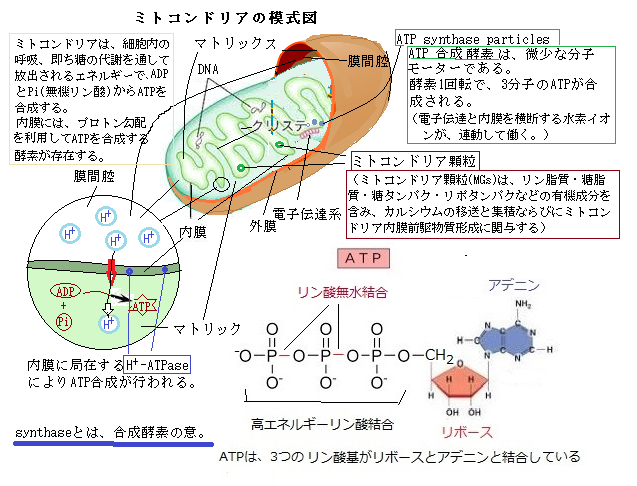
典型的な植物細胞は、エネルギー生産を行う2種類の細胞小器官、即ちミトコンドリアと葉緑体を持つ。どちらも外膜と内膜の2重膜でサイトゾルから隔てられ、自前のDNAとリボソームribosomeを持っている。ミトコンドリアは、細胞内呼吸により糖を代謝して放出されるエネルギーを用いてADP(アデノシン二リン酸)とPi(無機リン酸)からATP(アデノシン三リン酸)を合成する。
ミトコンドリアの内膜には、プロトン(水素イオン)の勾配を利用して細胞に必要なATPを合成するATP合成酵素synthaseが存在する。内膜に埋め込まれた、または内膜表面に存在する一群の電子輸送体、電子伝達系electron transport chainの働きにより、プロトン勾配proton gradientが生成される。膜を横切るプロトンの濃度勾配は、葉緑体やミトコンドリアで利用されATP合成の駆動力になる。
葉緑体のATP合成酵素は、チラコイド膜上にある。光合成が進む間、光を原動力にした電子伝達反応によりチラコイド膜を挟んでプロトンの勾配が生じる、ミトコンドリアの場合と同様、ATP合成酵素を介してプロトンの勾配が散逸する際にATPが合成される。そのATPは、葉緑体では、サイトゾルには運ばれず、空気中の二酸化炭素から炭素を固定する反応を含む、多くのストロマでの反応に利用される。
 酸素分子には、最外殻軌道電子に反応性の高い不対電子を持たないのに、その様々の分子種のROSは、細胞内の様々な分子から酸素電子(e-)を還元され、その相手の分子から酸素電子を酸化する、極めて反応性に富んだ分子である。
酸素分子には、最外殻軌道電子に反応性の高い不対電子を持たないのに、その様々の分子種のROSは、細胞内の様々な分子から酸素電子(e-)を還元され、その相手の分子から酸素電子を酸化する、極めて反応性に富んだ分子である。様々な非生物的ストレスは、細胞内にROSを蓄積させ、植物の発生や成長、及び収量に負の影響を与える。その一方では、ROSの蓄積により、様々な環境及び内生シグナルに反応し、順応機構の誘導に関与するシグナル伝達経路を活性化させことで、細胞に正の影響を与えるシグナル伝達分子であることが明らかになって来た。この反応は、ROSを蓄積が抱える負の影響を打ち消すほどの効果が認められている。
殆どのROSは、ミトコンドリアと色素体、ペルオキシソームperoxisome及び細胞壁で生成される。細胞膜に局在するNADPH酸化酵素が、ROS生成酵素の最もよく理解されたファミリーである。
細胞内膜系に由来するにも関わらず、独立して成長・増殖・融合する細胞小器官がある。オイルボディ・ペルオキシソーム・グリオキシソームが代表的である。
多くの植物は、種子の発達過程で多量の油脂を合成し貯蔵する。これらの油脂はオイルボディと呼ばれる細胞小器官に蓄積される。小胞体からの出芽によって分離したオイルボディは、特殊なタンパク質であるオレオシン・カレオシン・ステロレオシンを含んだ「半単位膜(単層のリン脂質の膜)」の内部に、油脂が合成され蓄えられている細胞小器官である。
種子が発芽するにつれオイルボディは分解するが、その際は脂質酸化酵素を含む細胞小器官であるグリオキシソームglyoxysomeに結合している。グリオキシソームは、種子の油脂に富んだ組織に存在する、1つの膜で囲まれた細胞小器官である。
マイクロボディmicrobodyは、1つの代謝機能のために特殊化した、一重の膜に包まれた球状の一群の細胞小器官である。脂肪酸のβ酸化やグリオキシル酸glyoxylate代謝などの複数種の機能の内の1つを担う。上記の2つの機能を、それぞれが担うマイクロボディが、ペルオキシソームとグリオキシソームである。ペルオキシソームとグリオキシソームは、かつては、小胞体から、それぞれ独立して形成された異なった細胞小器官と考えられていた。しかし、キュウリの芽生えでは、緑化していない子葉は、初めの段階では、グリオキシソームを持つが、緑化した後は、ペルオキシソームのみとなる。
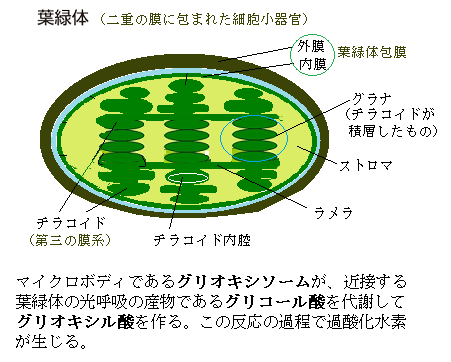
マイクロボディであるグリオキシソームが、近接する葉緑体の光呼吸の産物であるグリコール酸を代謝してグリオキシル酸を作る。この反応の過程で過酸化水素が生じる。グリオキシル酸は、2炭素アルデヒド酸で、グリオキシル酸回路glyoxylate cycleの中間体である。グリオキシル酸回路とは、グリオキシソーム内で、2分子のアセチルーCoAをコハク酸にかえる一連の反応を言う。
脂肪性種子の脂質がスクロースへ変換するのは、発芽がきっかけで起こる。その変換は、オイルボディに蓄積されたトリアシルグリセロールの遊離脂肪酸への加水分解から始まる。その脂肪酸が、グリオキシソームと呼ばれる、一種のペルオキシソームperoxisomeで酸化され、、アセチルーCoAが生産される。
アセチルーCoAは、グリオキシソームとサイトゾルで代謝されコハク酸となり、グリオキシソームからミトコンドリアへ輸送され、そこで最初にフマル酸に変換され、それからリンゴ酸に変換される。
この過程は、サイトゾルで起こる糖新生によるリンゴ酸のグルコースの変換と、グルコースからスクロースの変換で終わる。
殆どの脂肪性種子で、アセチルーCoAの約30%が呼吸によるエネルギー生産に使われ、残りがスクロースに変換される。
糖を分解してビルビン酸にする解糖は、殆どの生物で起こるが、多くの生物では、類似の径路を逆方向に働かせることもできる。この有機酸から糖を合成する過程を、糖新生gluconeogenesisと呼ぶ。コハク酸はミトコンドリアへと移行し糖新生に利用されるなど、炭水化物以外の前駆体からグルコースが生合成される。解糖系の中間体、アミノ酸・脂肪酸・グリセロールなどクエン酸回路中の物質などから糖形成ができる。
種子は、炭素を油脂(トリアシルグリセロール)として蓄積する。その種子が発芽する時、油は糖新生によりスクロースに変換され、発達中の芽生えの細胞に供給される。
もう1つの疑問は、花粉管から放出された精細胞の挙動を決めているのは何かである。
放出された精細胞は、融合の準備をするために雌性配偶子とシグナルを交換している可能性がある。例えば、シロイヌナズナでは、精細胞の到達直後に卵細胞からシステインに富むタンパク質が放出される。精細胞は、細胞表面に特異的な膜タンパク質を分泌して、これに応答している。この精細胞表面のタンパク質は、雄と雌の配偶子の融合を促進しているらしい。この仮説を裏付けるかのように、この表面タンパク質を欠く変異体では、精細胞は卵細胞や中央細胞のいずれにも受精することができない。
目次へ
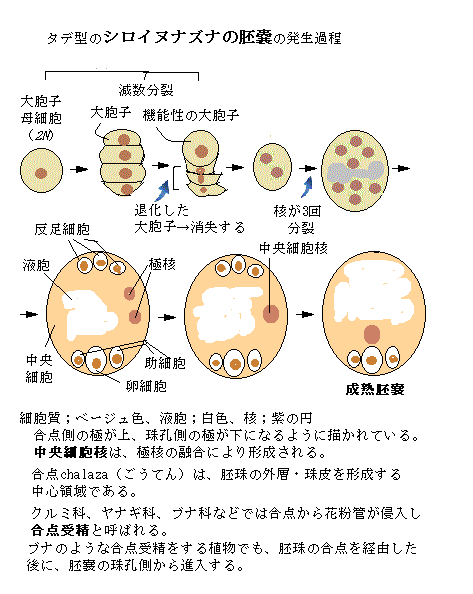
 胚珠原基は、先端が丸みを帯びた円錐状の突起として胎座に沿って生じる。原基の発生初期から3つの領域に分かれている。
胚珠原基は、先端が丸みを帯びた円錐状の突起として胎座に沿って生じる。原基の発生初期から3つの領域に分かれている。茎状の珠柄に分化する基部領域。
減数分裂の場となる珠心nucellusが生じる末端部の珠孔領域。
胚珠の外層である珠皮を形成する合点chalaza(kəléɪzə)と呼ばれる中心領域。
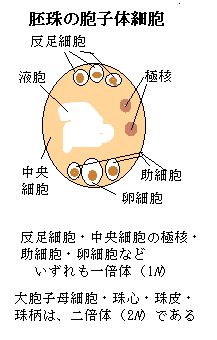
その中心領域は、大胞子母細胞へと発達する胚珠原基の段階では、珠皮に包まれた胚嚢を持つ「基部領域」にあった。
大胞子母細胞megaspore mother cellへと分化する細胞は、細胞自体や核のサイズが大きく、しかも細胞質密度が高い。そのため始原珠心の中に大胞子母細胞として、はっきりと観察される。大胞子母細胞は、機能性の大胞子として発達し、中央細胞の2つの極核は融合して、中央細胞核が形成される。珠孔は、受粉後に花粉管が伸長する伝達組織の出口に位置するようになる。
胚嚢の合点側にある3つの一倍体(1N)細胞は、反足細胞antipodal cellと呼ばれている。antipodalとは、「地球上の正反対側の」・「正反対の」の意味である。それらは、同じ胚珠の中で、珠孔とは逆側に位置しているからである。微細構造解析から、反足細胞の膜は大きく陥入していることが分かっている。栄養交換やホルモン情報伝達を担っている可能性が高い。
しかしながら、スイレン目の植物やアカバナ科のマツヨイグサの仲間は、反足細胞を持たず、これら2つの植物群では、成熟胚嚢は4核である。シロイヌナズナなど多くの種は、反足細胞は受精前に退縮するから、受精には直接関わっていないようだ。
一方、イネ科の植物では、反足細胞が増殖するため、受精に関与している可能性が高い。
反足細胞は、たとえばモデル植物のシロイヌナズナでは、胚嚢の成熟に伴いしだいに退化するのに対し、トウモロコシでは、反足細胞は、さらに分裂を続け、数を増やしたりする。植物種による違いが大きい細胞である。
生殖に対する特定の機能は提唱されていない。ただ、反足細胞や、あるいは受精後の胚乳endospermの細胞で最も反足細胞側にある細胞など、このあたり(胚嚢の合点側)にある細胞が、胚乳への養分の通り道にあたり、その供給にとって重要であろうと考えられている。
タデ型の胚嚢を持つ植物の重複受精では、2つの精細胞の内の1つが、卵細胞と受精して胚を形成し、もう1つの精細胞は中央細胞と受精して三倍体の一次胚乳細胞primary endosperm cellを形成する。一次胚乳細胞は、有糸分裂を繰り返し種子の栄養器官である胚乳へと発達する。その胚嚢型により極核の数が異なるため、胚乳の倍数性レベルは、マツヨイグサ属の2Nから日本にもシマゴショウとサダソウの2種が自生しているペペロミア属の15Nまで多種多様となる。
目次へ