| Top 車山高原 車山高原お知らせ 車山ブログ
|
 シダ植物に見る 胞子体と配偶子 シダ植物は、花が咲かないので種子はできない。胞子で増える。 種子が発芽して育つ植物と異なり、胞子が発芽して、まず前葉体ができ、 前葉体で受精してシダ植物になる。 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成
花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1)
遺伝子の発現(2) 遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝
減数分裂と受精 対立遺伝子と点変異 疾患とSNP 癌変異の集積 癌細胞の転移 大腸癌 細胞の生命化学
イオン結合 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 シグナル伝達 キク科植物
陸上植物の誕生 植物の進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応
根による水吸収 稲・生命体 胞子体の発生 花粉の形成 雌ずい群 花粉管の先端成長 自殖と他殖 フキノトウ
アポミクシス
| 目次 |
| 1)アポミクシスapomixisとは、「無性生殖」による繁殖体の形成である |
| 2)アポミクシスapomixisは、穀物の品種改良を促進する契機になる |
農作物の育種は、優れた形質を持つ品種間の何世代にも渡る交雑によって成し遂げられる。その優れた複数の形質が、組み合わされて優良品種が出来たとしても、引き続く有性生殖の過程で、その優れた形質が分離して、他の様々な形質が混入してくる。
その優良品種の栽培のためには、一代限りの雑種(ハイブリッド;haibrid)種子を、種苗会社などに頼り、毎年購入するようになる。
アポミクシスapomixisとは、「無性生殖」を意味するギリシ ャ語で、花粉が卵に受精しなくても種子を形成する「無性生殖」を言う。精子を必要としないため、卵細胞から母親と全く同じ遺伝子を持った、クローン(種子)が作られる。この特性により、簡単にクローン作物を作ることができるため、農業生産では非常に有用な研究となる。しかし、農業における主要な作物やその近縁植物には、アポミクシスの素質がなく、今まで、農業的に利用された例は殆どない。
近年、アポミクシスに関わる遺伝子を見つけようという取り組みが、世界的に活発になっている。 植物の生殖様式として「有性生殖」がよく知られており、 花粉が卵に受精することによって種子が作られる。父親(花粉)と母親(卵)によって子供(種子)が作られる。一方、アポミクシスでは、父親は必要なく、母親の卵から母親と全く同じ遺伝子セッ トを持つクローンの子供(種子)が作られる。この生化学反応は、実に、興味深いだけではなく、簡単にクローン作物を作ることが可能となり、農業的にも極めて有用である。
ところが、農業的に主要な作物やその近縁植物はアポミクシスの素質がなく、これまでに農業的に利用された例は殆どなかった。そのため、アポミクシスに関わる遺伝子の発見研究が、世界的に活発になってきた。
主に植物において、通常、有性生殖によって生じる繁殖体が、受精を伴わない無性生殖によって生じることがある。本来、花を作る位置に球根や零余子(むかご)、あるいは芽が形成されることが、典型的なアポミクシスの例である。一方、、古代から行なわれてきた挿し木や切れ藻からの無性的な生長は、アポミクシスを研究する手法とは、別次元の領域である。
受精を伴わないため、アポミクシスによって生じた繁殖体は、親植物と遺伝的に同じクローンとなる。 植物の中には、減数分裂と受精によって繁殖体が作られるのではなく、胚珠の組織の1つが受精卵へと分化したものから作られるものもある。この場合、胚の遺伝子型(植物の形質を決める遺伝子の総体、あるいは対立遺伝子の総体)は、母親と同一になる。この遺伝子型が同一な子孫を、無性的に生み出す方法をアポミクシスapomixisと呼び、この方法で生み出された植物をアポミクトapomictと言う。
 アポミクシスは、種子などの繁殖体を、受精を伴わずに生み出す生殖過程を総称する用語である。
アポミクシスは、種子などの繁殖体を、受精を伴わずに生み出す生殖過程を総称する用語である。減数分裂によって生じた卵細胞などの生殖細胞から分化し、そのまま繁殖体を生じる単為生殖(雌が単独で新個体を生じさせる生殖法)や、助細胞や反足細胞から繁殖体を生じる単為生殖などがある。
例えば、セイヨウタンポポの無配生殖は、アポマイオシスapomeiosisと呼ばれている。相同染色体の遺伝的組換えなどの交雑を伴わない大胞子母細胞(2N)の減数分裂の結果、半数体の染色体(1N)を持った大胞子が形成される。その結果、単為発生した1Nの卵を有する雌性配偶体(胚嚢)が生じ、そのまま受精を経ずに、種子が形成される過程である。これにより、親と遺伝的に同一のクローン植物が、無限に生じることになる。
この受精を伴わないアポマイオシスによる繁殖様式としている顕花植物は、40 属 400 種以上がそれを行なうが、主要な農作物では殆ど存在しないとされている。
分子細胞学的手法を用いて、遺伝子単離や機能解析が進められているが、未だ未解明のままである。
通常の胚は、配偶子(精細胞と卵細胞)が受精してできた接合子(受精卵)が分裂を繰り返して形成される。それが、種子の中に納まる。アポミクシスによる無融合種子形成には、様々なメカニズムによって生じており、その多様なメカニズムによる種子生産形式を単純に分類することは難しい。
例えば、葉・茎・根・花などの細胞の一部を取り出し、ある条件下で無菌的に培養すると、それらの組織を構成している細胞が分裂をはじめ、細胞が増殖する。これらの細胞は、もとの分化した細胞ではなく、未分化の状態のもので、ただ細胞の塊のようなもので、これをカルスと呼んでいる。
不定胚の方は、植物体の雌性配偶体が接合子と同じような過程をへて直接に形成される胚と同等のものとなる。しがって、不定胚は、分化したものであるから、培養によって完全な個体にまで育てることができる。
また、胚嚢の母細胞からではなく、珠心の体細胞から分化し、単為生殖的に胚嚢が形成され、胚に成長することもある。
アポミクシスには、個体内にアポミクシスと有性生殖が混在する条件的アポミクシスと、アポミクシスしか存在しない絶対的アポミクシスがある。いずれも、同種内に有性生殖個体が存在すれば、優良な一代雑種(F1)の親品種の遺伝子型をアポミクシスで維持し、特性の均一な品種を大量に生産する技術などに応用できる。アポミクシスでは、母株と全く同じ遺伝子型をもつ種子が得られるので、ヘテロシスheterosis(雑種強勢)の固定など、画期的な育種技術の開発に繋がる可能性が高い。
アポミクシスによって生じる植物は、元の植物と遺伝子的に同一なクローンとなるため、農作物で利用される一代雑種の親品種を固定する時などに利用される。
これまでも、近縁アポミクシス種との交配育種により、アポミクシス形質を主要穀類へ導入しようと、様々な実験が行なわれてきたが、成功例は未だない。しかしながら、遺伝子解析から、いくつかの植物種では、アポミクシスが優性遺伝子により支配されていると観察され、DNAマーカーを用いた連鎖解析によるアポミクシス遺伝子の単離が、キビ属(日本では五穀の一つ)などで進められている。
ギニアグラスの学名はパニカム・マキシマムで、イネ科のキビ属、和名はギニアキビと言う。暖地型イネ科植物で、飼料用牧草や緑肥兼用種である。そのギニアグラスでは、有性生殖系統およびアポミクシス性系統のそれぞれに、その系統を掛け合わせて目的の特性を持った品種を作出し、それ以降はアポミクシスによってその特徴を維持することで品種を固定している。
アポミクシス性ギニアグラスに関しては、珠心細胞から胚発生し、繁殖体を生じる無配生殖が、柑橘類やマンゴーでも観察されている。
もともとアポミクシスを行わない植物に、アポミクシス性を導入することによって、一代雑種の生産を容易にする試みがなされている。コムギなどでは、アポミクシスによって繁殖する系統が見出され、育種に利用されている。イネ科のキビ属も、アポミクシス性ギニアグラスが見られる植物である。
アポミクシスは、単子葉植物と真正双子葉植物を合わせて、被子植物の約0.1%にあたる40以上の科でみられる。例えば、ナナカマド・キイチゴ・ブラックベリー・セイヨウヤブイチゴ・柑橘類・マンゴー・セイヨウタンポポ・リンゴやキビ類の牧草などである。
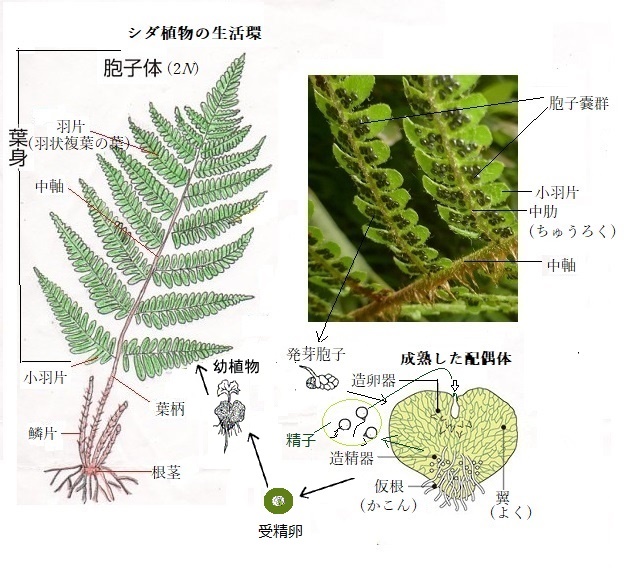
シダ植物の配偶体は、本来雌雄の配偶子が融合して胞子体を形成するが、配偶子が単為生殖によって胞子体を形成する無配生殖を行うことがあり、これもアポミクシスに含まれる。
アポミクシス植物が持つ共通の染色体領域が、イネ科のチカラシバで解明されている。チカラシバ近縁植物種では、生殖様式がアポミクシスである植物種には、相似性の高いアポミクシス特異的染色体領域(ASGR)が存在し、有性生殖の植物種にはASGR が存在していない。しかしながら、アポミクシス植物種において、ASGR が座乗する染色体の形態は、染色体長・腕比・凝縮パターンなどで様々に異なるのみではなく、染色体識別マーカーとして用いられる45SrDNAの有無の違いなどが見られるなど多様である。アポミクシス、はヘテロシス固定・育種年限の短縮などの利用が期待されているが、そのメカニズムの多くは、解明されいないし、ヘテロシス固定のためには欠かせないハプロタイプ(各遺伝子座位にある対立遺伝子のいずれか一方のDNA配列)が座上する遺伝子座も特定されていない。
オスとメスが分かれて いる動植物には、性を決定する重要な性 染色体があり、例えば、ヒトは23番目の染 色体がXXなら女性に、 XYなら男性になる。アポミクシス植物も花粉も作るので、他家受精させることができる。同一植物種間で交雑が起こったときに、アポミクシス特異的染色体領域(ASGR)が有性植物種に伝搬することによって、有性生殖から無性生殖へ転換するという仮説が提唱される。
イネ科のチカラシバ近縁植物種で、生殖様式が無性生殖(アポミクシス)である植物種間であれば、相似性の高いアポミクシス特異的染色体領域(ASGR)があり、その有性生殖の植物種には、ASGR が存在しない。これらの植物種の遺伝的距離は近く、生殖様式の関連性も高いので、ASGRを詳細に解析すれば、アポミクシス をコントロールする遺伝子を発見できるのではないか、と期待されていた。
だが、有性生殖とアポミクシスの生殖様式を有する、同じイネ科の植物種であるチカラシバ属 とキビ属植物のASGRについて比較解析により、当初予想されていたよりも、アポミクシスを有するチカラシバ属では、約50Mbp(百万塩基対単位)の巨大なアポミクシス特異的染色体領域が観察されており、この領域がアポミクシスを制御すると考えられている。モデル植物のシロイヌナズナのゲノムでも、小さな真正双子葉とはいえ、約157Mbpある。ASGRは非常に巨大で、しかも、先述したようにアポミクシス植物種間において、ASGR が座乗する染色体の形態は、染色体長・腕比・凝縮パターンが異なるのみではなく、染色体識別マーカーとして用いられる45SrDNA の有無にまで、差が見られるなど極めて多様である。解明までの道のりは遠く険しい。
目次へ
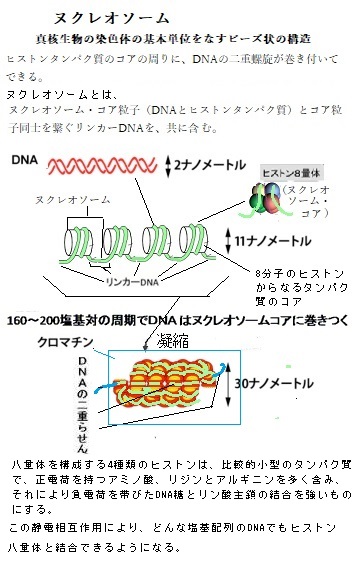 核は、DNAと付随タンパク質からなる染色体の複製と貯蔵を行う細胞小器官である。遺伝子とは、本来、DNAの塩基配列によってコード化された、細胞の代謝・成長・分化を制御する主要な遺伝情報の最小単位のことで、物質ではなく概念である。このDNAとタンパク質複合体が、生物の形質に変化を引き起こしている。
核は、DNAと付随タンパク質からなる染色体の複製と貯蔵を行う細胞小器官である。遺伝子とは、本来、DNAの塩基配列によってコード化された、細胞の代謝・成長・分化を制御する主要な遺伝情報の最小単位のことで、物質ではなく概念である。このDNAとタンパク質複合体が、生物の形質に変化を引き起こしている。DNAとタンパク質複合体(染色体)が、クロマチンchromatinと呼ばれる小さく折り畳まれた構造になっている。
植物のゲノムの全DNAを直鎖状に引き伸ばせば、その長さは染色体が存在する核の直径の数百万倍にも及ぶ。このDNA染色体を核内に収納するために、DNAの二重らせんの糸は、8つのヒストンhistoneタンパク質分子からなる筒状構造に二重に巻き付き、ヌクレオソームnucleosomeを形成する。ヌクレオソームは、染色体DNAの全長にわたり糸を巻いたビーズ状の構造をなす。
細胞周期の間期における、核にある染色体の配置は、ランダムではないし、絡まってもいない。それぞれの染色体は、核内の特定の場所にある。これを染色体テリトリーchromosome territoryと呼ぶ。この間期おける染色体テリトリーが、遺伝子発現に影響するか、ましてどのように影響するかは未解明のままである。
生物の形質変化を引き起こしているのがDNAであることに変わりはない。DNAは小さく折り畳まれて染色体になる。従ってDNAは、どの染色体にも存在している。そのDNAの遺伝的な変化が、その生物の形質に変化を引き起こしている。
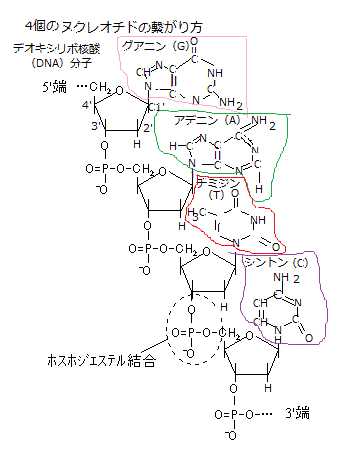
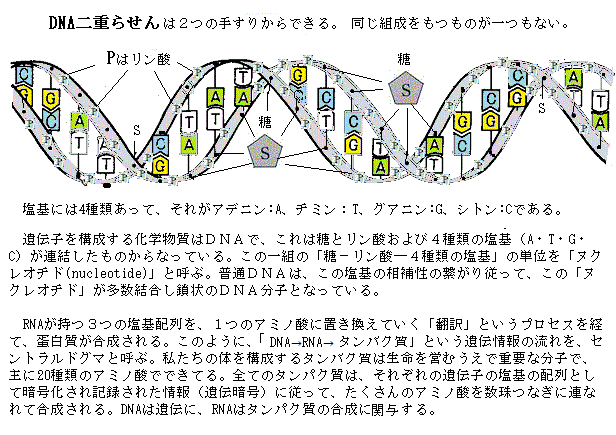
すべての生物は、この4種類の塩基のみを共通に使い、「塩基配列」の順序で遺伝情報を作り出す。実は、この塩基配列が「遺伝子」つまり「生物の設計図」そのものである。親から子へ受け継がれる遺伝子の本体はDNAで、DNA分子中の塩基配列の順序が遺伝情報として意味をなす。
 「遺伝情報の流れは常に DNAから RNAへ、RNAからタンパク質へと流れ、逆流することはない」というセントラル・ドグマ(central dogma)を、 1958年に F.H.C.クリックが提唱した。その後、一部修正を受けたが、基本的にはタンパク質から核酸がつくられることはなく、核酸からタンパク質への情報の流れは常に一方向というのが現在の定説となっている。
「遺伝情報の流れは常に DNAから RNAへ、RNAからタンパク質へと流れ、逆流することはない」というセントラル・ドグマ(central dogma)を、 1958年に F.H.C.クリックが提唱した。その後、一部修正を受けたが、基本的にはタンパク質から核酸がつくられることはなく、核酸からタンパク質への情報の流れは常に一方向というのが現在の定説となっている。核には、核小体nucleolusと呼ばれる粒子状のものが密に詰まった部位がある。ここでリボソームribosomeが合成される。通常、細胞の真核には1つの核小体が存在するが、複数の核小体を持つ核もある。その核小体には、1本から数本の染色体中のリボソームRAN(rRAN)遺伝子が密集している核小体形成領域nucleolar organizer region(NOR)がある。実は、各染色体は核内では、バラバラと見えるほど離れて存在している。その何本かの染色体の中ほどの領域が集合して核小体を形成しているようだ。
核小体で、リボソームタンパク質とrRANが会合して大小のサブユニットが形成され、それぞれのサブユニットは、核膜孔を通って細胞質に運ばれる。その複数のサブユニットが、細胞質で会合して、翻訳装置となる。その実態は、タンパク質合成装置であるリボソームである。その殆どがタンパク質とrRANでできている真核のリボソームと同様の装置が、細胞質に存在する小器官、ミトコンドリアや色素体の原核でも形成される。しかし、真核によるリボソームの方が、より大きい。生物の進化の過程が、ここでも読み取れる。
真核や原核は、細胞のDNA情報を読み出す、転写transcriptionする場である。
リボソームを構成するrRANは、核小体形成領域(NOR)から転写される。多くのリボソームは、タンパク質合成には欠かせないため、NORには、rRAN遺伝子が数百コーピー存在する。植物種によって異なるが、ゲノム中には、1つかそれ以上の個所にNORがある。トウモロコシは、6番目の常染色体に1ヵ所ある。

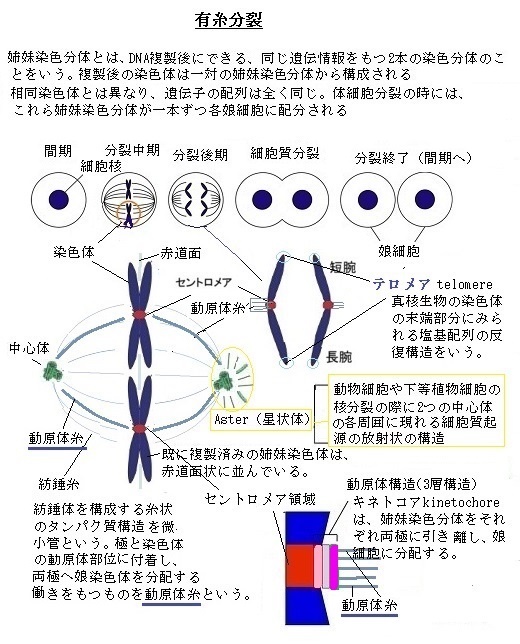
配偶子形成の際の細胞分裂は、体細胞分裂と当然似てはいるが、いくつかの重要な点で異なっている。
その初期の減数分裂の第一分裂では、それぞれの染色体が娘細胞に分かれる前に、相同染色体間で、DNAの遺伝子領域で交換が行われ、遺伝物質の組み換えが起こる(減数分裂組み換え)。
減数分裂の第二分裂では、姉妹染色分体が分離し、もとの細胞1つから4つの娘細胞が生じる。減数分裂は、この2回の分裂からなるが、DNA複製は一度しか起こらない。それぞれの娘細胞は、もともと細胞から半分の遺伝物質しかもたないようになる。
この二倍体の植物体が、半数体の配偶子を作ることを減数分裂還元と言う。
ヒトは、染色体を2組23対もつ(何対もつかは、生物の種類により異なる)。23対のうち22対は常染色体で、1対は性染色体(せいせんしょくたい)といって、性の決定に関わる。この1対がXYなら男、XXなら女。生物種によって、染色体の数やら、性決定の組み合わせは異なるから、注意が必要。
雄はX及びY性染色体を有するのに対して、雌はX性染色体を2本有している。配偶子は一倍体であるから、雄の配偶子(精子)はX、またはY性染色体を運ぶ。女性の配偶子(卵子)はX性染色体のみを運ぶ。配偶子がX染色体が運ばれる数とY染色体が運ばれる数は同数である。卵子がX性染色体を運ぶ精子で受精すると、性染色体はXXとなり、雌になる。卵子がY染色体を運ぶ精子と受精すると、性染色体はXYとなり、雄になる。
それぞれの染色体上に存在する遺伝子は、メンデルの法則にしたがって遺伝する。一般に体細胞の染色体数は2Nで、生殖細胞のそれは1Nで表される。
「近代遺伝学の父」メンデル(G.J.Mendel)の論文は2つある。エンドウマメを材料にした1,866年の第一論文には、「メンデルの法則」がすべて網羅されている。
その後、メンデルは、オシロイバナ・トウモロコシなど様々な植物を使って、法則を検証した。ただヤナギタンポポ属の多くの種は、雄しべを除いて他家受粉をしても、子はすべて純粋な母親の形質を示した。雑種が全くできず、法則に合わなかった。
メンデルが、1,870年に発表した第2論文で、「悪い母」の代表としたのが、ヤナギタンポポ属の多年草コウリンタンポポであった。セイヨウタンポポは、アポミクシスにより種子を作るため、雑種ができにくいのである。
コウリンタンポポは、ヨーロッパの高山が原産だが、北海道・東北やアラスカなどに帰化している。ロゼットから30cmほどの花茎を伸ばし、夏に朱色の美しい頭花を付ける。
ヤナギタンポポ属は、ヨーロッパを中心に世界の温帯に1,000種ある、属名はギリシャ語のhierax(タカ)に由来する。タカが視力を強めるため食べるという伝承からである。
ヨーロパを代表するセイヨウタンポポは、現在は雑草として世界中に拡散している。もともと畑地の雑草であったため、夏に休眠せず、刈られると直ぐに新しい芽をだして光合成を行う。小さな個体であっても、発芽して半年で、花芽を付ける。種子の軽さは、日本の在来種の半分ほどで、タンポポ属の中でも最も遠くへ飛ぶ。また幅広い温帯域で、いつでも活着できる。
タンポポと言う名は、歴とした日本語である。古来、日本に生育していた在来種と、近世に海外から持ち込まれ帰化種がある。在来種は外来種に比べ、開花時期が春の短い期間に限られ、種の数も少ない。また、在来種は概ね茎の高さが外来種に比べ低いため、生育場所がより限定される。夏場でも見られるタンポポは、殆ど外来種のセイヨウタンポポとなっている。
在来種の各種とセイヨウタンポポは基本的に別種ではあるが、細胞中の特定の酵素・アイソザイムIsozymeをターゲットにした解析では、交雑が起こっていることが報告されている。
生物の多くは異なる個体同士で、子孫を残す有性生殖を行ない、遺伝子を交換する。一方で、単独で自分と同じ遺伝子型を持った子孫を残す無性生殖を行なう生物もいる。
有性生殖には、様々な環境ストレスを克服して交配相手を探す難点がある一方で、遺伝的な多様性を獲得することにより、環境変化に強い耐性を持つ確立が高まる。無性生殖では単独での繁殖が可能なので、高い繁殖力と確実性が増すが、遺伝的多様性を保つことが期待できないため、種全体として環境変化や新たな外部ストレスに適応リスクが高くなる。
有性生殖と無性生殖双方の自然環境に対する適応能力を実証するためには、近縁種または 同種内で有性生殖個体と無性生殖個体が、同時に存在していることが望ましい。
陸上植物ではアポミクシスと呼ばれる無性生殖の様式があり、通常、陸上植物の雌性配偶体である胚嚢における有性生殖過程では、珠心細胞から分化して、核分裂を経て卵細胞・中心細胞・助細胞などを含む胚嚢が形成される。その後、卵細胞と中心細胞へ、雄性配偶体の精細胞が重複受精に成立すれば胚発生が起こる。
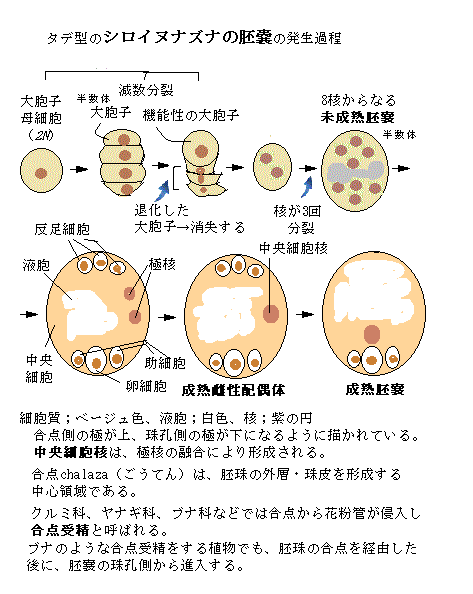
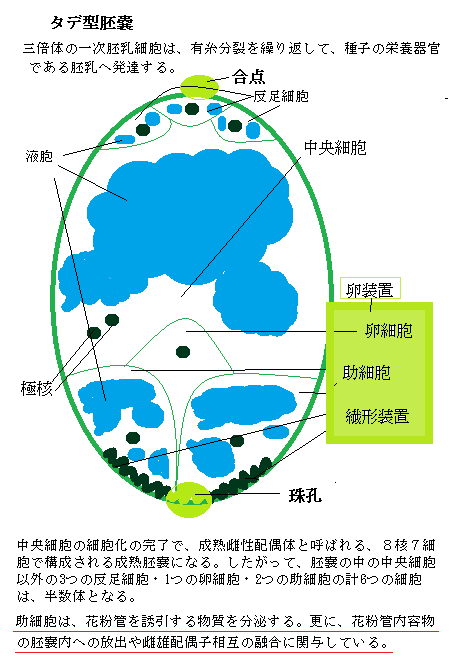 雌性配偶体である胚嚢embryo sacの発生は、雄性配偶体に比べて複雑で多種多様である。被子植物の胚嚢発生パターンは、15種類以上はあると言われ、そのうち最も一般的なパターンは、どこにでも生える典型的な雑草であるタデ属で、タデ型と呼ばれている。
雌性配偶体である胚嚢embryo sacの発生は、雄性配偶体に比べて複雑で多種多様である。被子植物の胚嚢発生パターンは、15種類以上はあると言われ、そのうち最も一般的なパターンは、どこにでも生える典型的な雑草であるタデ属で、タデ型と呼ばれている。そのタデ型の珠心内の胞原細胞は、減数分裂を行う大胞子母細胞へと分化する。胞原細胞とは、雌性の大胞子母細胞megaspore mother cellに分化する細胞群、または雄性の花粉母細胞(小胞子母細胞microspore mother cell)を言う。
上記の図のように、タデ型の胚嚢は、最初に減数分裂により、二倍体(2N)の大胞子母細胞から半数体(1N)の大胞子が4つ形成される。通常、その後、珠孔側の3つの大胞子がプログラム細胞死をし、機能を持った大胞子(1N)が1つだけ残る。
この残された大胞子が、細胞質分裂を伴わない3回の核分裂(有糸分裂)をして多核体coenocyte(ˈsiːnəoseit;大型の細胞に多数の核が含まれた状態)になる。これが、半数性の8核からなる未成熟胚嚢である。胚嚢内には大きな液胞が育ちつつある。8核の内4つは合点側に、残りの4つが珠孔側の核として移動する。各極の3つの核が、それぞれ側で細胞化する。残った2つの核が合体して二倍性diploidyとなり、極核polar nucleiと呼ばれ、胚嚢の中央領域へ移動する。中央領域の細胞質と2つの極核が、細胞膜と細胞壁を形成すると、2核を持つ大型の中央細胞central cellになる。
中央細胞の発生は、卵細胞と異なるが、重複受精時に、2つの精細胞の内の1つと融合するので、中央細胞も配偶子とされている。この中央細胞の細胞化の完了で、成熟雌性配偶体と呼ばれる、8核7細胞で構成される成熟胚嚢になる。したがって、胚嚢の中の中央細胞以外の3つの反足細胞・1つの卵細胞・2つの助細胞の計6つの細胞は、半数体となる。
タデ型の胚嚢を持つ重複受精では、1つの精細胞が卵細胞と受精して胚を形成し、もう1つの精細胞が中央細胞と受精して三倍体の一次胚乳細胞primary endosperm cellを形成する。この一次胚乳細胞は、有糸分裂を幾度か重ねて、種子の栄養器官である胚乳へと成長する。
タデ型の胚嚢の合点側の3つの細胞が、半数性の反足細胞antipodal cell(antipodal;対蹠的な,意)で、栄養交換やホルモン情報の伝達に関与しているようだ。イネ科植物の反足細胞は、増殖するため、受精に関与している可能性が高いと見られている。
胚嚢の珠孔側にあるのが卵細胞egg cellと助細胞synergid cellで、合わせて卵装置egg apparatusと呼ばれる。卵細胞こそが、花粉管の精細胞と融合して胚を形成する雌性配偶子である。
助細胞は、花粉管を誘引する物質を分泌する。「LURE(ルアー;誘引する意)」と名付けられた、植物の種類ごとに異なる低分子のアミノ酸・ペプチドで構成されている。これにより花粉が異なる植物種の雌しべに受粉しても「受精」しない理由の一つとして知られた。
2つの助細胞の最も珠孔端に近い側に葉、繊形装置filiform apparatusがある。filiformとは、「糸状の」の意味で、繊形装置は、複雑に入り組んだ厚い細胞壁が細胞内に突出して、細胞膜の表面積を大きくしている。繊形装置の細胞膜に局在するFER-RLKタンパク質が、雄性配偶体である花粉管からの分泌される特異的リガンドligand(タンパク質が結合する物質は、イオン・有機小分子・巨大分子を問わず、そのタンパク質のリガンドと呼ぶ)を受容し、同種であると認識した場合に、繊形装置の細胞内情報伝達機構が働き、花粉管が胚嚢内への進入を許す生理的変化を示すと見られている。
 アポミクシスでは、胚嚢を生み出す母体由来組織の珠心細胞から、減数分裂を介さずに大胞子母細胞(2n)から配偶体が分化するアポスポリーaposporyと、セイヨウタンポポのように、胚嚢細胞でアポマイオシスapomeiosisが起こり、相同染色体の分離や遺伝的組換えを伴わない、大胞子母細胞の減数分裂
を経て、親と同じ半数体の染色体数を持った大胞子が形成される。
アポミクシスでは、胚嚢を生み出す母体由来組織の珠心細胞から、減数分裂を介さずに大胞子母細胞(2n)から配偶体が分化するアポスポリーaposporyと、セイヨウタンポポのように、胚嚢細胞でアポマイオシスapomeiosisが起こり、相同染色体の分離や遺伝的組換えを伴わない、大胞子母細胞の減数分裂
を経て、親と同じ半数体の染色体数を持った大胞子が形成される。子房内にある胚珠では、大胞子母細胞(2N)の減数分裂によってできた4個の娘細胞の内、3個が退化・消失して1個だけが残る。この細胞が、やがて雌性配偶体となる大胞子(1N)である。
大胞子は3回の体細胞分裂を行なって8個の半数体をもつ雌性配偶体になる。8個の核のうち1個だけが卵細胞(1N)になり、これが雌性配偶子である。残りは1Nの助細胞・中央細胞の極核・反足細胞になる。
アポミクシスは、遺伝子型が同一の子孫を残すという繁殖形態から、かつては、進化における袋小路で、可逆的に絶滅へと向かう系統の分岐点と推測されていた。
その後の系統的な解析により、アポミクシスが、進化の分岐の初期に起こった系統と、後期に起こった系統の双方で見られることや、かつてアポミクシスが起こった系統で、再び有性生殖が行われるという可逆性が明らかになると、それまでの仮説が覆った。
アポミクシスの遺伝子制御には、珠心や助細胞、及び反足細胞など雌性配偶体の正常な発達を調節する遺伝子群の発現に変化が生じたことで起こる。アポミクシスは、倍数体植物が環境適応度を上昇させてきた進化の過程で現れた、幾度もたどってきた形態形成と理解される。そのためアポミクシスの生理現象が複雑多岐となり、現段階では、明確に分類するにいたっていない。
トウモロコシのように味覚と量産性に優れた穀物の多くは、ヘテロシスheterosis(雑種強勢)の長所を生かした交雑品種である。アポミクシスの仕組の解明により、品種改良のための強力な新技術の開発が期待されている。
交雑植物では、同じ形質をもつものを、その種子から完全に再現すことができないため、優良な交雑品種の種子を作るには、その親系統の交配を毎年繰り返す必要がある。しかし、アポミクシスをF1雑種に導入できれば、親と同一の遺伝子型の種子を量産できるようになり、F2世代でのヘテロシスの喪失を防ぐことができる。この技術は、穀物の品種改良を促進する契機になることから、近年、種子のメカミズムに関する研究が激化している。
目次へ