| イネ科植物 Poaceae | ||||||||||||
|
||||||||||||
| DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精 ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応 植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2) 遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積 癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸 生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生 植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生 花粉の形成と構造 雌ずい群 花粉管の先端成長 自殖と他殖 フキノトウ アポミクシス 生物間相互作用 バラ科 ナシ属 蜜蜂 ブドウ科 イネ科植物 |
||||||||||||
| 目次 | ||||||||||||
| 1)イネ科の穀物(ライムギ・オートムギ) | ||||||||||||
| 2)トウモロコシ | ||||||||||||
| 3)イネ科植物(サトウキビ・トウモロコシ・キビ・アワ) | ||||||||||||
| 4)種子登熟 | ||||||||||||
| 5)陸上植物の細胞と組織 | ||||||||||||
| 6)被子植物の花粉形成過程では数多くの遺伝子が発現する | ||||||||||||
| 7)生殖細胞の決定の分子機構 | ||||||||||||
| 8)緑の革命green revolution |

1)イネ科の穀物
 |
| エーゲ海沿岸のリゾート地,「クシャダス」 |
ライムギは、カフカス・小アジア原産で、コムギより酸性土壌に強く、乾燥や寒冷な気候にも適応し、ロシア連邦やスカンジナビア半島はもとより、ドイツや東ヨーロッパなどでは主要な穀物として栽培されている。実を製粉して黒パンを作り、麦芽はウオツカやビールの原料、中世にはオオムギに代わってコムギに次ぐ第2の穀物としての地位を確立していた。
麦はデンプンを多く含んでいるが、酵母はそのデンプン自体をアルコールに変えることはできない。まず、デンプンを糖分に変える必要があり、その糖化のために必要な酵素を麦の中に作らせるため、麦を発芽させる。それを麦芽malt(mɔ́ːlt)と呼ぶ。
ライムギは、風媒花で、しかもコムギとは近縁である。互いの交配も可能である。ただし、ライムギの花粉がコムギの雌しべに受粉される場合に限り受精する。そのため、コムギの花粉が、ライムギの雌しべに受粉しても、実は稔らない。 ライムギはコムギやオオムギと違って、本来的には、他家受精植物のようで、自家受精の時は、著しく収穫量が減る。ライムギは、もともとコムギ畑に生える雑草で、自然環境の生育条件がほぼ一致する。ただ、ライムギは発芽温度が1℃~2℃と低く、低温に強いため冬作物として栽培される。そのため、コムギの生育条件の悪い畑では意図的にコムギとライムギを混栽することがある。コムギが寒冷で不作となった時季、ライムギの収穫で、ある程度補填する。
オートムギ(エンバク)は、英語名の「Oat」からの呼び名で、イネ科カラスムギ属に分類される一年草で、その種子は穀物として扱われる。原産地は地中海沿岸から肥沃な三日月地帯(ペルシア湾からチグリス川・ユーフラテス川を遡り、シリアを経てパレスチナ、エジプトへと到る半円形の地域)と、中央アジアにかけてであり、この地方には現代でも野草化したオートムギが広く自生している。
 6,000年から7,000年前の肥沃な三日月地帯の遺跡において、オートムギの栽培の痕跡は、未だ検出されていない。しかしこの地方には、やがてオートムギの野生種が自生し、コムギやオオムギ畑に入り込んで雑草として生育するようになる。雑草のオートムギの種子は、適合する環境条件下であれば発芽するが、その集団が一斉に発芽するわけではなく、発芽せずに残るものがある。これは、すべての発芽を、その環境条件だけに合わせれば、成長の過程で、一度、その条件が悪化すれば、成長途上にある全種子が死滅することに繋がる。このリスクを回避するための種よる適応だと考えられている。これは、最悪の環境を想定しながら、種の保存を大前提に達成したオートムギの進化の積み重ねによる成果である。
6,000年から7,000年前の肥沃な三日月地帯の遺跡において、オートムギの栽培の痕跡は、未だ検出されていない。しかしこの地方には、やがてオートムギの野生種が自生し、コムギやオオムギ畑に入り込んで雑草として生育するようになる。雑草のオートムギの種子は、適合する環境条件下であれば発芽するが、その集団が一斉に発芽するわけではなく、発芽せずに残るものがある。これは、すべての発芽を、その環境条件だけに合わせれば、成長の過程で、一度、その条件が悪化すれば、成長途上にある全種子が死滅することに繋がる。このリスクを回避するための種よる適応だと考えられている。これは、最悪の環境を想定しながら、種の保存を大前提に達成したオートムギの進化の積み重ねによる成果である。オートムギの栽培植物の多くは、発芽や収穫の効率を上げるため、それぞれの独自の休眠性が奪われ、好適な条件下であれば一斉に発芽するように品種改良されている。その採集の生産性が向上したために、約5,000 年前には、中央ヨーロッパで主要な作物になっている。 栽培は秋蒔きと春蒔きとがある。オートムギは冷涼を好むも、ライムギほど耐寒性は高くない。寒冷地では、凍害により越年できないことが多い。そのため、通常、温暖な土地では秋蒔き、寒冷地では春蒔きを行うことが多い。
オートムギは、寒冷で高緯度の痩地で栽培されることが多く、そのため世界的には春蒔きによる生産が主流となっている。また、オートムギは、ムギ類の中でも乾燥に最も弱く、特に湿潤を好み、生育には多量の水を必要とする。生育期に乾燥が激しくなると収穫量に悪影響を及ぼす。腐植土を好むが、生育地の幅は広い。酸性に強く、酸性土壌で広く生育するが、アルカリ性土壌にも耐えられる。よく成長するが、そのため分、倒伏しやすい。
オートムギは、調理脱穀したオートグローツが基本となる。大きくわけるとオートグローツには、加熱したものと、加熱しないものの2種類がある。加熱したものを粉砕せずに、あらく砕いたオートグローツを蒸してから、平らに引き延ばし、乾燥させて圧扁したものがロールドオーツである。加熱処理されており薄くなっているので調理しやすい。約4倍量の水を加え、5分ぐらい煮て、ロールドオーツが膨らみ水分がなくなったら完成、ハチミツやブラウンシュガーと相性が良いようだ。ドライフルーツやフルーツグラノーラなどを加えて食べても美味しい。
オートミルは脱穀されているから調理しやすい。通常、小麦とは異なり精白されず、胚芽が残るため、タンパク質が多く、食物繊維・鉄分・カルシウム・ビタミンB1・ミネラルなどの栄養素が豊富に含まれている。脱穀しただけであるため、用途次第であるが、焙煎・粉砕・圧扁・加熱などの調理で消化されやすくなる。
ロールドオーツを更に食べやすくするために細かく砕いたクイックオーツを、粥のように、鍋に入れて水や牛乳に浸して、焦げ付かないようにかき回し、全体に火を通せば、クリーミーで、甘さが際立ち、美味しく食べられる。クイックオーツをもっと簡単に調理しやすくしたのが、インスタントオーツ、基本的に味付けされているのでそのまま食べることも可能で、調理する場合でも簡単なアレンジもしやすい。
目次へ
2)トウモロコシ
 トウモロコシは、イネやコムギと並ぶ世界三大穀物に数えられる。南米アンデス山麓原産のイネ科トウモロコシ属の一年生植物で、種実(「しゅじつ」とは、硬い皮や殻の中の食用果実)を食料や飼料にし、葉や茎を飼料にする。主成分はデンプンで、タンパク質・脂質・糖質をバランスよく含んでいる。
トウモロコシは、イネやコムギと並ぶ世界三大穀物に数えられる。南米アンデス山麓原産のイネ科トウモロコシ属の一年生植物で、種実(「しゅじつ」とは、硬い皮や殻の中の食用果実)を食料や飼料にし、葉や茎を飼料にする。主成分はデンプンで、タンパク質・脂質・糖質をバランスよく含んでいる。コーン油(トウモロコシ油)は、トウモロコシの胚芽から得られる。コーンスターチcornstarchを生産する時の副産物である。そのコーンスターチは、トウモロコシの穀粒から精製されたデンプンで、原料のトウモロコシを、亜硫酸液に浸し、柔らかくなったところで粗挽(あらび)きし、胚芽分離機にかけて、デンプン・タンパク質・繊維質の混合物と胚芽とに分ける。次に脱水し乾燥させて製品にする。
良質なコーン油は、胚芽に水蒸気を当て、圧搾する。色は通常淡い黄色で、濃いオレンジ色の場合、胚芽だけでなく、種子からもオイルが摂られている場合がある。コーン油は、ビタミンEが豊富なので、酸化には比較的強い。しかも、大豆油と同様、植物油なので、リノール酸を豊富に含むので、加熱調理にはあまり向かない。極めて、良質のサラダ油なので、ドレッシングなど温めないものと混ぜて利用したほうがよい。コーン油は、血中コレステロールを下げる作用があり、動脈硬化などのリスクを低下させる。
油は三大栄養素の1つ、脂質の仲間、脂質はエネルギー源になるだけではない、脳の約65%は脂質であるばかりか、神経の伝達機能を維持するためにも脂質は欠かせない。細胞の膜や体の様々な働きを調節するホルモンに似た物質などの材料になる。
リノール酸は、必須脂肪酸、ただ取りすぎると免疫細胞が働きにくくなり、アトピー性皮膚炎や花粉症などのアレルギー性炎症疾患を引き起こす。また、動脈硬化や心臓病などを誘発する。植物系のリノール酸は、菓子・パン・マヨネーズ・カップ麺・総菜など加工食品やファストフードに含まれる。これらの「見えない油」が、知らず知らずのうちに摂取過多を招く。
トウモロコシの考古学的史料によれば、野生祖先種は紀元前5,000年ころにはメキシコに分布し、その栽培型が成立したのは紀元前3,000年ころと見られている。北アメリカへの伝播も古く、ニュー・メキシコ州では紀元前2,300年の遺跡から出土している。
トウモロコシ栽培は、
メキシコ西部・北部 →北米南西部 → 北米東部 → カナダへ伝播、
南米へのルートは、メキシコ南部・東部 → グアテマラ → カリブ諸島 → 南アメリカ低地 → アンデス高地と伝播したようだ。
マヤ文明やアステカ文明では、トウモロコシが大規模に栽培され、両文明の根幹を支えていた。
ヨーロッパへの導入は、1492年、クリストファー・コロンブスがアメリカ大陸を発見した際、現地のカリブ人が栽培していたトウモロコシを持ち帰った。ヨーロッパには、ほぼ即座に栽培面積が広がり、1,500年には、スペイン南部のアンダルシア州セビリアにおいて栽培植物としての記録が残っている。
日本には、天正7(1,579)年に中国を経由して、ポルトガル船が長崎に持ち込んだのが最初で、そのため「唐キビ」「南蛮キビ」とも呼ばれた。明治初年には、アメリカから北海道に入り、北海道で盛んに栽培された。
トウモロコシの茎は直立し、高さ2~4m、節は14~16のものが多く、殆ど分枝しない。葉は互生し、葉身は大きいものでは幅5~10cm、長さは約60cm線形で、先端が垂れ下がり、中脈は太く白い。茎頂には10数本に枝分かれした雄花序(ゆうかじょ)を付ける。雌雄異花で雌花は夏に花を開く。雄花序は、茎の先端に抽出して雄穂(ゆうすい)と呼び、雌花序は茎の中部の2~4節の葉腋(ようえき)に生じ雌穂(しすい)と呼ぶ。そのうちの1~2個だけが熟する。これが食用になる。外側には数枚の苞があり、中の太い芯の上に10~20列の果実が規則正しく並ぶ。果実の付け根には、薄い鱗片があるが、これが苞類や外花頴(がいかえい)が退化したものである。
トウモロコシは、先端の花序に、栄養的な優先権がある。しかし、頂端に雄花序が付いていても、雄花序はそれほど養分を必要としないため、その下の雌花序への栄養は十分残されている。
 雄穂は、通常の品種では、長い穂軸(すいじく=芯;しん)から十数本の枝柄が分かれ、各枝柄に多数の雄性の小穂(しょうすい)が対(つい)になってつく。各小穂は2小花からなり、小花には3本の雄しべがある。
雄穂は、通常の品種では、長い穂軸(すいじく=芯;しん)から十数本の枝柄が分かれ、各枝柄に多数の雄性の小穂(しょうすい)が対(つい)になってつく。各小穂は2小花からなり、小花には3本の雄しべがある。トウモロコシの雄花序と雌花序は相同器官で、容易に入れ替わる。トウモロコシの黒穂菌(くろぼきん;黒穂菌が寄生して多くは穂、時に茎葉に黒色の病斑が生じ、黒色の粉が充満する)に侵されたトウモロコシでは、成長ホルモンの分泌に異常が生じ、しばしば茎頂に雄花序でなく雌花序が付く。
トウモロコシ体内のホルモンの分布により、茎頂に雄花序が付き、そのホルモンが下方に下りて、ある限界値を下回ると雌花序が付くようになる。花序の性が、根からの距離により決まる現象のように見えることになる。
雌穂は何枚もの包葉(トウモロコシの果実を包む皮)に包まれた円柱形で、穂軸の表面には雌小穂が縦に10~20列並ぶ。小穂は2小花よりなるが、下位の小花は不稔(ふねん)で、上位の小花のみが結実する。最初の雌花序が成熟して、その穀粒が膨らんでいく間、その枝の基部から出る二次枝の雌花序の成長は抑制される。
トウモロコシの進化の過程で、メキシコ南部の不安定な気候では、乾燥が続くことが予想され、最初の雌花序だけを成熟させ、下方に副次的にある花序は不稔になっていった。この適応進化の過程に加えて、人類がアメリカ大陸に渡った 11,000年前からは、1個だけの雌花序だけが、より大きな穀粒となる一方、その他の雌花序の発現は抑制される品種を選抜することによって、やがてそれが退化し、側枝の節間(せっかん)は短縮されるようになった。
この側枝の節間の短縮により、茎頂から下ってくるホルモンの境界領域の末端に、雄性から雌性に変化する「性的ジャンプ」が生じるようになった。人間による選抜の積み重ねによって、トウモロコシは途方もない多様な地方品種を持つ穀粒作物に急速に進化した。
上位小花の雌しべの花柱と柱頭は長く糸状に伸び、それを絹糸(けんし)と呼ぶ。これが各小花より伸び出して、開花時には雌穂全体の絹糸が束状に包葉の先から現れる。その本数はすべて苞に包まれた子房とつながり、実の数と一致する。
同じ個体では、茎頂の雄花が開いて花粉を飛ばしている時、雌花の花柱(トウモロコシの毛)は未だ伸びてはいない。その後、2日ほどして熟し伸長する。即ち雄性先熟の仕組みで、風媒花のトウモロコシは他家受精をする。開花後、雌穂は長さが14日目ころまで、太さは20日目ころまで発達する。開花後約50日で完熟する。種子の色は黄・白・赤茶・紫・青などがある。
目次へ
3)イネ科植物(サトウキビ・トウモロコシ・キビ・アワ)
十分吸水したイネの種子は、10~15℃の温度があれば発芽し、1本の種子根を地中に伸ばし、同時に幼芽を地上に出して主茎とし、その主茎を支える鞘葉(しようよう)から第1葉、第2葉と順次展葉していく。
酒精飲料は、エチルアルコ ールを含んだ飲料の総称であるが、その多くはイネ科の穀粒を原料としている。サトウキビは、強風や水不足に対して大変強い作物で、台風や旱魃などの厳しい自然環境にある鹿児島県南西諸島及び沖縄県においては、貴重な基幹作物であり、農家の約7割が栽培している。
サトウキビの栽培には、春に植え付けを行い翌年の冬に収穫する春植、夏に植え付けを行い翌々年の冬に収穫する夏植、冬の収穫後の株から出る芽を育て、翌年の冬に再度収穫する株出(かぶだし)の3種類の方法がある。生産地では、1月から3月頃がサトウキビ収穫の最盛期となり、この時期には、家族総出で刈り取りを行う光景も見られる。生産地の島々にある製糖工場では、サトウキビを原料として原料糖を製造しており、サトウキビを満載したトラックが島中を行き交い、製糖工場の周辺では甘い香りが立ち込める。
製糖工場では、工場内のボイラーで発電する。その燃料には、原料糖の製造工程で生じる「バガス」と呼ばれるサトウキビの搾りかすが利用されており、必要な電力のほとんどが賄われている。しかも、余った「バガス」は、畑の肥料や家畜の飼料にも利用されている。
その原料糖は、消費地に近い精製糖工場に運ばれて、さらに純度が高められ、白い結晶の「砂糖」になる。
トウモロコシは、食用油(コーン油)やコーンスターチ(トウモロコシの穀粒から精製されたデンプン)を製造するための工業用作物であるが、肉類・牛乳生産のための牧畜飼料作物として、青刈用と種実を用いる実取用とに分けられる。他の牧草の大部分も、イネ科植物である。
イネ科植物は、芝生からタケ・ササ類まで、形態も変化に富み、用途も広い。世界には約600属1万種以上分布し、日本では127属約500種が知られている。単子葉植物のイネ科の殆どは、イネのように細長い葉と茎(稈)を持った草本であるが、イネ科タケ亜科のうち、タケは、木本のように茎が木質化する種の総称である。ササ類の殆ども、稈が何年も生き新しい枝を出す木本型の植物である。通常の双子葉植物の木本の幹と異なり、その稈は一定以上の太さにならない。二次肥大成長するための形成層を欠くためである。これは草本の特徴である。イネ科の稈は原則として中空で、節があり葉をつける。
葉は葉身と葉鞘(ようしょう)からなる。細長い扁平な葉身は容易に分かるが、葉身の基部を巻いて稈の節間を包んでいる部分が葉鞘である。葉身と葉鞘の境目には小さな襟のような葉舌(ようぜつ)がある。
イネ科植物の葉は、茎を取り囲んだ筒状の鞘となっており、「葉鞘」と呼ぶ。その筒状の葉鞘が、やがて葉に移行する。この葉と葉鞘の移行部を観察してみると、葉は風に吹かれて揺れ動き、結構強い力がかかるに違いない。この葉鞘が葉につながるが、その部分は特殊な形をしている。これは、風などによる葉にかかる力から、葉を守るためのものと思われる。この力のかかる部分は肥厚し幅広くなっているが、この部分を「葉耳(ようじ)」と呼ぶ。茎は節から送り出されるように伸長するので、茎に固定するわけにはいかない。この力のかかる部分は肥厚して葉をしっかりと支えている。この肥厚部分が幅広くなると、「葉耳(ようじ)」と呼ばれる構造になる。画像の白い部分がそれである。葉鞘の先端部分には、薄い襟のような膜状の構造がある、これを「葉舌(ようぜつ)」と呼んでいる。これらの構造物は種を同定するポイントでもある。
1.葉の付け根に筒状の膜(葉舌)があるか?あるとしたらそれは長いか、短いか?
2.葉の付け根にふくらみ (葉耳) があるか?この部分の毛の量は多いか、少ないか?
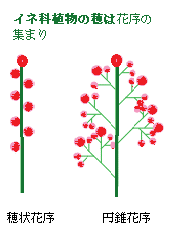 花には雄しべ、雌しべの他に花被が退化した鱗被(りんぴ)という小片がある。雄しべは、イネやタケ・ササ類では、6本だが、多くの種では3本に減っている。イネ科植物の穂は花序の集まりであり、真の花序は枝の先についている小穂である。イネ科・カヤツリグサ科植物の花穂で、花序の最小の単位となる穂状の枝に付く花の集まりである小穂は、イネ科の分類や種の同定には、最も重要な部分である。小穂の中軸は小軸と呼ばれ、その上に頴(えい)と呼ばれる鱗片が2列に並ぶ。
花には雄しべ、雌しべの他に花被が退化した鱗被(りんぴ)という小片がある。雄しべは、イネやタケ・ササ類では、6本だが、多くの種では3本に減っている。イネ科植物の穂は花序の集まりであり、真の花序は枝の先についている小穂である。イネ科・カヤツリグサ科植物の花穂で、花序の最小の単位となる穂状の枝に付く花の集まりである小穂は、イネ科の分類や種の同定には、最も重要な部分である。小穂の中軸は小軸と呼ばれ、その上に頴(えい)と呼ばれる鱗片が2列に並ぶ。頴は、イネ科以外の植物では、苞に相当する器官である。最下の2枚の頴の内側は空で苞頴(ほうえい)と呼ぶ。3枚目の頴は外花頴(がいかえい)と呼ばれ、その内側には1枚の内花頴(ないかえい)がある。外花頴と内花頴の間に本当の花がはいっており、外花頴、内花頴と一個の花は一体となっているので、この3つを合わせて小花(しようか)という。頴の名称は、専門家の間でも不統一である。
小穂は、多くのイネ科植物では、円錐花序に配列しているが、コムギやカモジグサでは穂状花序を作る。
トウモロコシなどのキビ亜科では、小穂は原則として2小花、時には1小花に退化し、さらに小型になって数個から多数が密生する一つの小穂群を作る。その小穂群が円錐状・穂状・総状などの花序を形づくる。キビ亜科では、こうしたの花序の構成単位である小穂群を「総(そう)」と呼んでいる。
キビは、イネ科の一年生草本作物で大型の円錐花序を持ち、小穂の長さは4~4.5mm。草丈は約1m。葉は幅広の剣状。寒さや乾燥に強く、栽培期間が短く、山間部の雑穀として品種の保全と開発が望まれている。夏から秋にかけ、茎頂に多数の花穂をつけ、実ると垂れる。実は2mmほどの扁球形で、黄色か黄白色で、品種によっては褐色のものもある。和名は実の色の「黄実(きみ)」が転訛したと言う。 栽培キビの原産地は、未だ定説はない。中央~東アジアの温帯地域とも、あるいはインド・パキスタンには近縁の自生種が分布するからとも言われている。中国山西省のキビは、トウモロコシ・ジャガイモ・コーリャン・アワなどと並ぶ主要な農産物になっている。世界の穀物としての栽培面積では、コムギ・イネ・トウモロコシ・オオムギに次いで第5位である。
『漢書』巻94の匈奴伝に、紀元前84年、天候不順で匈奴の「穀稼(穀物)、孰(みのら)ず」とある。遊牧民の匈奴であっても、農耕は、枢要な生業であった。近年、極東ロシアのザバイカルのブリヤート地域にある匈奴の40の墓から123粒のキビの種が、そしてウランバートル北方約100kmのノイン・ウラ山中にある匈奴墓からもキビの種が出土した。ロシアの東シベリア、ブリャーチヤ共和国にあるバイカル湖西岸のイヴォルガ遺跡から、製鉄遺跡群が発掘調査され、犂先・シャベル・鍬などの農具類が出土している。共伴する殆どの土器が漢代中国の灰陶に酷似しており、遊牧民の匈奴国家においても農耕は重要な生業であった。
太古の遊牧民の生業に不可欠な農耕を支えてきたキビは、石器時代から穀物の代表作物であった。紀元前8世紀は、ヨーロッパにも伝播し、特に遊牧民を主体とする諸民族集団であるスラブ民族の農耕にとって不可欠な存在となっていた。
しかし日本には、五穀の中でも最も遅く伝来している。今では、日本への伝来は、華北から朝鮮を経て伝わったと推定されている。
日本書記には持統天皇即位7(693)年の春、
「丙午、詔令天下、勸殖桑紵梨栗蕪菁等草木、以助五穀(3月17日、詔(みことのり)があり、天下に、桑・紵(からむし;繊維素材となる植物)・梨・栗・蕪菁(=ブセイ/かぶら;蕪;菁=すずな;かぶの別称)などの草木を植えるよう奨励し、五穀(米・稗・粟・麦・豆)を補うよう布告した)。」
「日本書記」第30巻の最後が、持統天皇紀であり、その布告の5穀は、米・稗・粟・麦・豆であるが、キビは含まれていない。米・稗・粟・麦・豆よりも遅れて栽培されたか、あるいは重要度が低かったようだ。
黍(きみ)を詠んだ歌は、万葉集には1首だけで
「梨棗 黍に粟つぎ 延ふ葛の 後も逢はむと 葵花咲く」
梨(なし)・棗(なつめ)・黍(きみ)に粟(あは)つぎ、延(は)ふ葛(くず)の、後(のち)も逢はむと、葵(あふひ)花咲く
「梨棗(りそう)とは離早で、愛している人と離れている、の意」「黍(きみ)に粟(あは)つぎ」は「君に逢はつぎ」、「葵(あふひ)」は「逢う日」。
古代に食用となった植物ばかりで、見事な恋歌が詠まれている。
フユアオイの古名は「葵」、多年草で冬でも枯れないのでその名がある。春から秋にかけて白や淡紅色の小花を、次々と咲かせ若葉や花柄は、苦み・えぐ味なく食用に、実は利尿に効ありとされている有用の植物である。
キビは、中国・日本・朝鮮などで常食する5種の主要な穀物である。現代の日本では、米・黍・粟・麦・豆を5穀とするが、黍の代りに稗(ひえ)をあげる説も多い。中国や朝鮮などでも5穀の定め方に諸説あって一定しないのと同じである。
日本では5穀の一つとされキビの茎は家畜の飼料になる。少なくとも、新石器時代からの人類の食用と飼料穀物であり続け、中国の華北地方では、アワとともに古代の主要穀物であった。
日本のキビは、縄文時代に渡来したアワより遅く、弥生時代に渡来したと考えられている。しかし、日本では、キビの品種改良は大きく行われたことがなく、各地の環境に適した在来種が栽培されてきたようだ。縄文時代の集落遺跡から、キビのプラントオパールの検出が予想される。
 イネ科の一年草のアワの茎は、1m未満のものから2mを超すものまである。日本には縄文時代に朝鮮半島を経て渡来した。最も古い栽培植物の1つで、縄文時代には既に栽培され、かつては日本でも広く栽培されいた。1株から1~2本の茎が直立し、茎先から円柱形の総状花序が出て、短い小穂が円柱状に並ぶ。長さ10~40cm、小穂は密生して、しかも分枝は短縮しているから小穂は隙間なく密着する。そのため、花序の中軸は外からは見えない。成熟すると、小穂は長さ2mmほどの黄色の実に熟し垂れ下がる。
イネ科の一年草のアワの茎は、1m未満のものから2mを超すものまである。日本には縄文時代に朝鮮半島を経て渡来した。最も古い栽培植物の1つで、縄文時代には既に栽培され、かつては日本でも広く栽培されいた。1株から1~2本の茎が直立し、茎先から円柱形の総状花序が出て、短い小穂が円柱状に並ぶ。長さ10~40cm、小穂は密生して、しかも分枝は短縮しているから小穂は隙間なく密着する。そのため、花序の中軸は外からは見えない。成熟すると、小穂は長さ2mmほどの黄色の実に熟し垂れ下がる。 鳴子きれて 粟の穗垂るゝ みのり哉 (正岡子規)
(鳴子とは、田畑の作物を荒す鳥を追払うためのおどし具で、小さい板に細い竹管を糸で吊り並べたもの。)
温暖で乾燥した風土を好み、生育期間が3 ~ 5ヶ月と短いために、高地や高緯度地域でも栽培することができる。
穎果(えいか)は、痩果の一種で、果皮が成熟後に乾燥して種子を覆う種皮(珠皮)と密着する実で、穀果ともいう。イネ科植物の種子の特徴で、イネの実で言えば、玄米を精白するときに、胚芽の部分と同時に、籾殻(もみがら)として取り除かれる果皮にあたる。ちなみに、胚芽米は籾殻を除去する際、「胚芽」を残した精米方法で、精米が難しく、販売されている種類も少ないので、価格は少し高くなる。ただ、胚芽は米が発芽をする部分であるため、多くの栄養素が含まれている。
アワの穎果は、花序の穂に密につき、「冬の野狐の尾」のように垂れ下がる。日本では最近、日常食には殆ど利用されなくなった。アワは穂の大きさなどで、オオアワとコアワに分類される。日本で栽培されているアワの殆どはオオアワである。中国ではオオアワは粱で、コアワが粟と分類されるが、日本ではアワ全体に粟の字を用いる。日本で栽培されるのは、オオアワである。原産地は東アジア地域とされ、エノコログサから分化したと考えられる。エノコログサ最大の変異植物である。
エノコログサは、世界の温帯に広く分布する一年草で、草丈は50cm以下、海岸の砂地、荒れた道端や山野でも普通に見られる。エノコログサの和名は、円柱状の花序を犬の尻尾に見立てて、漢字の「狗尾草(えのころぐさ)」を当てたことによる。むしろ「猫じゃらし」の名で通用している。世界の温帯に広く分布する。
アワの原種は、日本では縄文時代から栽培され、品種の分化が進んでいる。日本へは、朝鮮を経て最も古く渡来し、ヒエと並んで我が国最古の作物で、水田稲作が伝来する以前には、主食であったと考えられている。乾燥に強く土質を選ばず、栽培期間は90~130日と短期で、寒冷地でも栽培は可能である。
花序の長さは20cmにもなり、熟しても果実は、簡単には壊れず、これが収穫後の作業を容易にする。
ヨーロッパには石器時代に伝わっており、イタリア・ドイツ・ハンガリーなどでは古くから栽培されている。中国では黄河流域で紀元前2,700年ころ栽培されたとする記録がある。原産地は東~中央アジアと推定されている。かつては、ドイツからスイス・イタリア・ウクライナ・ ロシア・ルーマニア、・ギリシアに至るヨーロッ パの主要雑穀は、キビとアワであった。
ヨーロッパ南部にトウモロコシ、北部にジャガイモが新大陸から伝播して以降、雑穀はこの500年ほどで急激に栽培面積を減らした。それでも、アメリカへは初期移民がヨーロッパから伝えて以来、少量栽培されていたが、1,849年から栽培が奨励され、20世紀にはアメリカの雑穀の90%を占めるに至った。現代では、日本を含め先進国では、小鳥の餌に利用されている。
生育期間が短いので、古代から高冷地ではもっとも重要な穀物の一つとなり、明治後期には20万ヘクタール余りも栽培されていた。大正以降は減り続け、現在では郷土料理用などに特別に栽培される程度で、菓子や小鳥の餌に常用されるものは、殆どが輸入品である。品種は多いが、栽培上は春アワと夏アワとに分ける。春アワは北海道や東北地方に適し、5月に種を播く。夏アワは西南日本の暖地で栽培され、6~7月に種を播く。
利用面からは、粘り気の少ないウルチアワと粘り気が強いモチアワとに分けられる。米と混ぜて粟飯とする場合はウルチ性が適し、粟団子や粟餅、菓子などにする場合はモチ性が適している。大阪名物の「粟おこし」は、アワを煎って飴で固めたもの、台湾では、モチアワが、アワ酒やアワ餅に加工され神事との関連が深いようだ。
エノコログサの基本種は、世界の温帯に広く分布し、日本全国のほか、朝鮮・台湾・中国南部の海岸の砂地に生育する、草丈50cm以下の小型の1年生草本をいう。海岸の砂地や岩の割れ目などで、基部から放射線状に広がり生える、草丈15cm以下の、しかも海岸型であれば、変種とされハマエノコロと呼ばれる。
林付近に生育するオオエノコロは、エノコログサとアワ(栽培作物)の交雑種であるが、その交雑種は多く形態も多種で、いろいろなタイプがある。
小穂の構造は、軸の基部に2枚1対の包頴があり、下から順に外包頴(がいほうえい)、内包頴と呼ぶ。その上にさらに2枚の頴があってこれを外花頴(がいかえい)と内花頴という。この外花頴と内花頴にはさまれて3本の雄しべと1本の雌しべがあり、これが本来の花に相当する。外花頴・内花頴・雄しべ・雌しべでできる小花は、1小穂上に1個のものから多数並ぶものまで様々である。
イネ科の植物の小穂の外側の先端にある棘状の突起を芒(のぎ)と呼ぶ。その芒の棘が内向きに生え、皮膚や衣服にひっかかり種子の散布を助ける。
目次へ
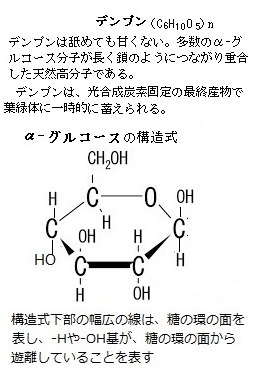
 植物の光合成産物は、エネルギー代謝や器官形成、単糖(糖類であるが、それ以上に加水分解できない糖の総称で、グルコース・フルクトース・アラビノース・マンノース・ガラクトース・リボースなどがある)やショ糖・デンプン・フルクトースなどの貯蔵炭水化物や貯蔵タンパク質、貯蔵脂質などの合成などに利用される。
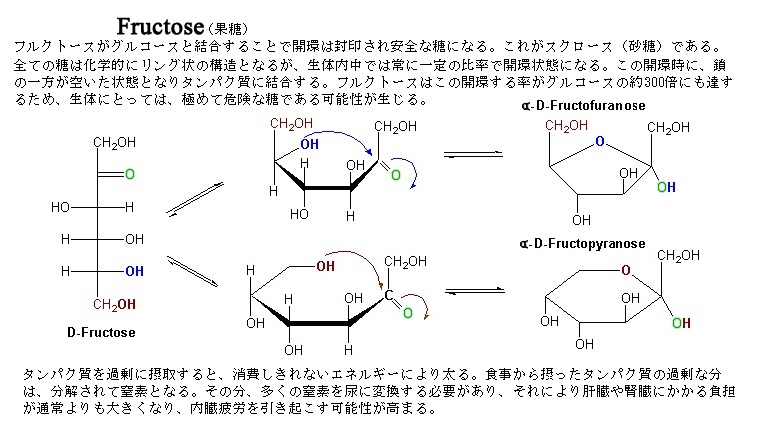
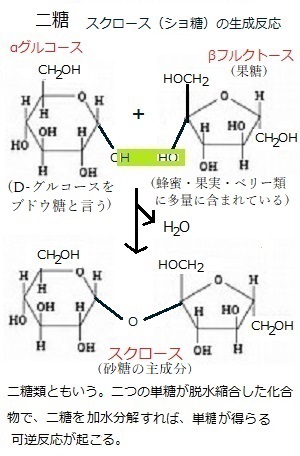
植物の光合成産物は、エネルギー代謝や器官形成、単糖(糖類であるが、それ以上に加水分解できない糖の総称で、グルコース・フルクトース・アラビノース・マンノース・ガラクトース・リボースなどがある)やショ糖・デンプン・フルクトースなどの貯蔵炭水化物や貯蔵タンパク質、貯蔵脂質などの合成などに利用される。そのため、一般に糖などの転流物質の濃度はソース側で高く、シンク側で低いことから、これにもとづく篩部輸送の機構として、ソース側の膨圧(植物細胞の細胞壁はかなり硬いので、小さな体積変化が大きな膨圧変化を生じさせる)により、ソース側の細胞内溶液が篩部を通してシンク側に押し出されるとされる、圧流説が提唱されている。光合成産物の転流形態は、基本的にはショ糖である。 ショ糖は、スクロースsucroseとも言う。工業製品である砂糖の主成分である。サトウキビやサトウダイコン(テンサイ)・ソルガム(モロコシ)などの多くの植物によって合成されるグルコースglucose(D-グルコース)とフラクトースfructose(果糖)が、脱水縮合した二糖類である。 フルクトースは、水によく溶けて、果実によく含まれるため果糖とも呼ばれる 。
化学式 C6H12O6、通常、D-フルクトース(ブドウ糖)のことをさす。広く植物中に存在する。フルクトースは単糖類の一つで、蜂蜜や果実・ベリー類に含まれている。フルクトースはグルコースと同じく還元性をもつ。それはケトン基のあとにCH2OHがついているためである。フルクトースは、単糖類の中でも最も甘い。分子量はグルコースと同じく180。
多くの果実では、デンプンが登熟する最中に、グルコースやフルクトースに変換される。また、果実には、クエン酸やリンゴ酸も多く含まれる。糖や酸は、味覚の重要な要素であるが、動物の食べごろの時季になると、殆どの液果では、その登熟に合わせて、糖や酸の化学変化により、口当たりがよくなり、香りの化合物も放出される。
フルクトースは、動物の精液の主要な糖で、精子のエネルギー源となる。 サトウキビは茎の柔細胞にショ糖sucroseを、リンゴなどの果実は果肉細胞にショ糖や果糖を高濃度に蓄積する。これらの糖は細胞の中の「液胞」に隔離・蓄積され、時には液胞内の糖濃度が1 M近くにもなる。液胞の中に糖を高濃度に隔離・蓄積するには、液胞の膜に糖の能動輸送体が必要だが、未だその遺伝子は特定されていない。液胞に糖を隔離することにより細胞質ゾルcytosolの糖濃度は、液胞より低く保たれている。
それでも数百mM(モル濃度とは、溶液1L中に溶けている溶質のモル数 [mol]で表した濃度)になる。 しかし、ウリ科やシソ科などはショ糖のほかにラフィノースやスタキオースなどのオリゴ糖を、またセリ科やバラ科の植物はショ糖に加えてマンニトールやソルビトールなどの糖アルコールを転流する。また、これらの転流糖類の篩管内における濃度は比較的高く、ショ糖転流型の植物では篩管液のショ糖濃度が1 Mを上回ることもある。このような篩管における高度な糖の集積には種々の糖輸送単体、即ち、ショ糖トランスポーターsucrose transporterなどが寄与している。現在知られている多くのショ糖トランスポーターは、伴細胞(ばんさいぼう;植物の篩管に付随する柔細胞で、篩管に平行して並んでいる)もしくは篩要素に局在している。
目次へ
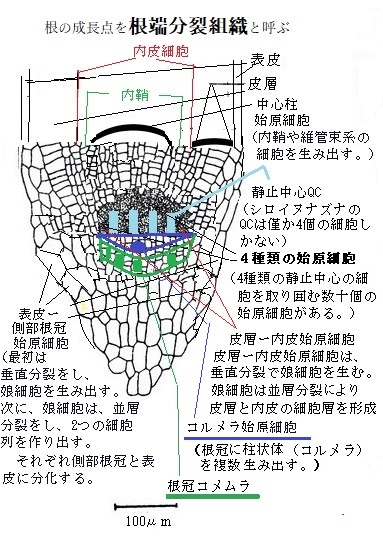
5)陸上植物の細胞と組織
 海で生まれた植物の先祖にとって、水は当たり前のように活用できた。それが一度、陸上に進出すると貴重な資源となった。陸上植物は、水が不足がちな陸上環境に適応するために、その根と茎と葉を進化させた。根は植物を固定し、水と栄養を吸収する。茎は背を伸ばし、葉を多く茂らせたが、水の運搬だけでなく、栄養を全身に満遍なく送り込む仕組みが必要になった。
海で生まれた植物の先祖にとって、水は当たり前のように活用できた。それが一度、陸上に進出すると貴重な資源となった。陸上植物は、水が不足がちな陸上環境に適応するために、その根と茎と葉を進化させた。根は植物を固定し、水と栄養を吸収する。茎は背を伸ばし、葉を多く茂らせたが、水の運搬だけでなく、栄養を全身に満遍なく送り込む仕組みが必要になった。葉は光エネルギーを得て、水を分解して水素と酸素を発生させ、二酸化炭素をATPやデンプンなど有機化合物に固定する。
陸上の二酸化炭素は豊富でも、水は僅かな水蒸気としてあるのみで、湿った地面に含まれた水分は必須である。植物が大型化するにつれ、根と葉の距離は遠ざかる。そのため根で吸収した水やミネラルを葉に送り、その一方、葉で合成したエネルギー源を含めた光合成産物を根に送る効率的な長距離輸送システムを進化させた。隣接する、もしくは長距離輸送する器官間の物質輸送は、通常は通導組織を経由する。
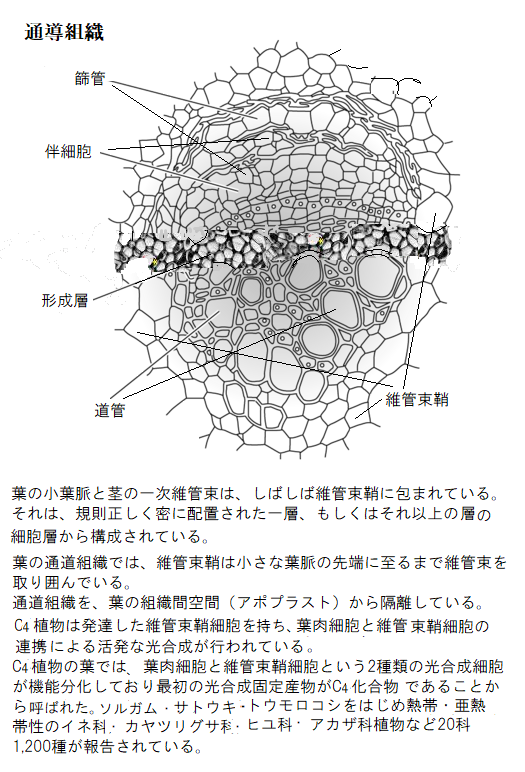 植物の成長は、細胞拡大に続き、特殊化した細胞への分化を伴う。表皮組織・基本組織・維管束組織は、すべての植物器官に見られる主要な組織系である。
植物の成長は、細胞拡大に続き、特殊化した細胞への分化を伴う。表皮組織・基本組織・維管束組織は、すべての植物器官に見られる主要な組織系である。表皮組織は植物の外側に1〜数層の細胞層からなる表皮と言う保護層を形成する。
基本組織は、植物体の大部分を占め、主として柔組織からなるが、茎や根の髄や皮層、葉の葉肉を含み、植物内を三次元的に充填する組織である。形態的にも生理的にも多種多様な組織を含む。双子葉植物の茎のように維管束が環状に配列する場合、基本組織は維管束の外側は皮層、内側は髄とよばれる。根では皮層だけで髄を欠く場合が多いが、根の皮層の内側に内皮と内鞘があり、さらにその内側に維管束形成層と一次木部と一次篩部が茎とは異なる形態で存在する。また葉の基本組織は葉肉(ようにく)と呼ばれ、もっぱら光合成を行う。 葉の場合は、表と裏を、「向軸面adaxial surface」と「背軸面abaxial surface」と言う。
葉の基本組織は向軸側にある細長い形をした柵状組織と背軸側の不規則な形をした海綿状組織の2種類からなる。
海綿状組織は細胞間に空隙が多い。中葉を介して隣接する細胞と接着している面が細胞壁表面の一部に過ぎないからである。
この構造により、光合成や呼吸の際の二酸化炭素と酸素のガス交換が可能なる。
柵状組織と海綿状組織の両細胞型には、葉緑体が多数存在し、細胞の縁辺部にならんでいるが、光条件の変化に応じて細胞内を移動することができる。
これらの細胞には、多数の球状のペルオキシソームperoxisomeや、数µmにまで伸びて枝分かれするミトコンドリアも存在している。
このペルオキシソームという細胞小器官は、小胞体とは独立して形成され、葉緑体やミトコンドリアと密着している。ペルオキシソームで、接する葉緑体の光呼吸の産物であるグリコール酸glycolic acid(CH2(OH)COOH)が酸化されグリオキシル酸ができる。この反応の過程で、他の化合物を容易に酸化し分解する過酸化水素H2O2発生し、ペルオキシソームが過剰に持つタンパク質である酵素のカタラーゼcatalaseによって水と酸素に分解される。
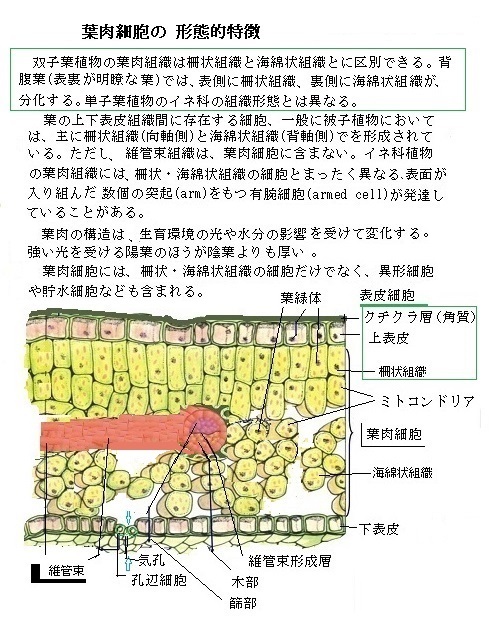 葉肉細胞のミトコンドリアと小胞体ネットワークは、葉緑体の近くに密着している。そのため、呼吸や光合成の際に産出される代謝中間体の効率的な受け渡しが容易になる。
葉肉細胞のミトコンドリアと小胞体ネットワークは、葉緑体の近くに密着している。そのため、呼吸や光合成の際に産出される代謝中間体の効率的な受け渡しが容易になる。これらの葉肉細胞は、環境さえ整えれば、他細胞へと分化できるので、柔組織の1つと言える。
柔組織は、薄い一次細胞壁を持つ基本組織系の細胞の1つであるため、分裂組織で作られても分裂活性は維持され、他の基本組織系や維管束組織系の細胞に分化できる。
若い柔細胞は、大き目の液胞やネットワーク状の多数の小さな液胞を持つ。これらが、やがて巨大な中央液胞へと成長する。
ニチニチソウの海綿状組織と柵状組織の葉肉細胞は、異形細胞idioblast(固有芽細胞)と言う特殊な性質を備えた細胞に分化する。この異形細胞は、ビンクリスチャンやビンブラスチャンと言う重要な抗がん剤を含む。草食動物に対抗する防御物質とみられる。
ビンブラスチャンは、タンパク質の重合体である微小管における、その重合を阻害する働きをする。
細胞質は、細胞骨格cytoskeleton(cyto;細胞、skeleton;骨格)と呼ばれる繊維状のタンパク質集合体により三次元的なネットワークとして構成されている。諸々の細胞小器官が立体的に配置される場や、その細胞小器官や他の細胞骨格構成因子の運動を支える足場を提供し、有糸分裂・減数分裂・細胞質分裂・細胞壁の構築・細胞の形態維持・細胞分化などに重要な役割を果たしている。
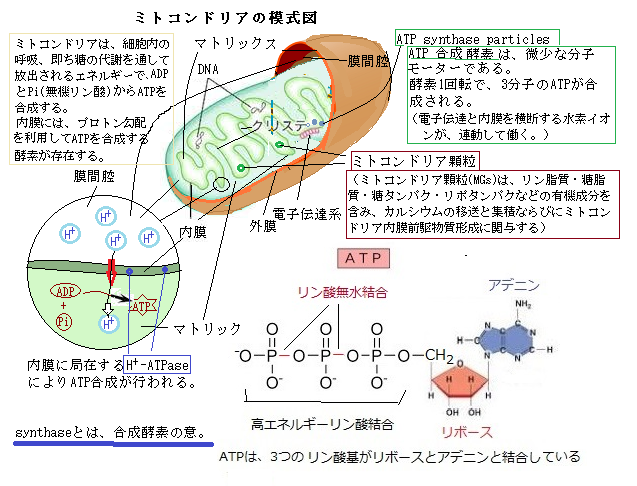 植物には、主要な細胞骨格構成因子として微小管とアクア繊維の2種類がある。その両者は高分子タンパク質で構成され、それぞれは決まった直径の球状からなり、それらが密に様々に重なるため、その長さが数µmになるものも生じる。
植物には、主要な細胞骨格構成因子として微小管とアクア繊維の2種類がある。その両者は高分子タンパク質で構成され、それぞれは決まった直径の球状からなり、それらが密に様々に重なるため、その長さが数µmになるものも生じる。一本の微小管microtubuleは、それを構成するチューブリンtubulinと呼ばれる球状タンパク質が数十万個重なる円柱状の重合体からなる。外径25nmの中空な円柱であるが、生物種や細胞型により形状は異なる。
チューブリンのサブユニットは、細胞内で遊離した状態で蓄えられているが、ビンブラスチャンは、微小管を非チューブ状の凝集体に重合させることで細胞分裂を阻害する。
異形細胞の近くにある柔細胞が、この毒性のアルカロイド生合成の中間体を供給する。各柔細胞は、それぞれ異なる中間体を供給しているため、柔細胞それぞれが化学的役割の分業を担っていることがわかる。これらのアルカロイドは、異形細胞の特殊化した液胞の中に蓄えられ、その毒性が細胞全体に散らないよう隔離されている。
柔組織は、肥厚した一次細胞壁を持ちながら伸長しつづけ基本組織系に分化できる。セロリの茎の筋などを構成する厚角細胞は、厚い層状の細胞壁を持ち、セロリのパリパリとした食感生み出している。
ゴムノキ・ケシ・リーフレタス・タンポポなどに見られる乳管laticiferのネットワークは、乳白色の乳液latexを運搬する機能がある。乳管を構成する乳細胞の一次細胞壁は、不均一で肥厚しているが伸長を続けることができる。ニチニチソウの葉の乳管は、異形細胞と同様にアルカロイドを含み柵状柔細胞と接している。
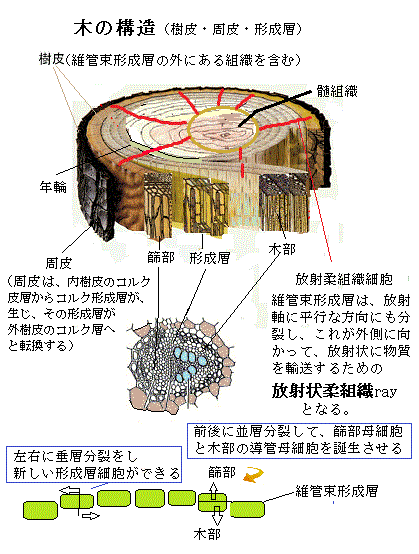 維管束組織は、植物の長軸に沿った水や溶質の輸送や移動を担う組織で、木部と篩部からなる。最初は、一次木部と二次木部からなる維管束の中の成長域である「維管束内形成層」として分断されているが、成長する過程で維管束形成層vascular
cambiumとして連結される。
維管束組織は、植物の長軸に沿った水や溶質の輸送や移動を担う組織で、木部と篩部からなる。最初は、一次木部と二次木部からなる維管束の中の成長域である「維管束内形成層」として分断されているが、成長する過程で維管束形成層vascular
cambiumとして連結される。維管束形成層が環状に繋がると、内側方向で分裂した細胞が二次木部を形成して、その外側には二次篩部を形成する。木部と篩部は、通道細胞・柔組織・肥厚した細胞壁を持つ繊維細胞(長い先細りの厚壁細胞で、維管束植物に強度を与える)で形成される。
植物の内側から外側へ向かう放射軸に沿った成長を二次成長と呼び、形成層cambiumと言う分裂組織がこれを行う。木質部分を造り出す形成層は、維管束形成層と呼ばれ、一次成長時に作られた木部と篩部の間に生じる。維管束形成層の細胞は、伸長軸に平行に分裂を繰り返し、茎や根の外側または内側に向かい細胞を供する。
維管束形成層は、また放射軸と平行しても分裂し、これが外側に向かって放射状に物質を輸送するための放射状柔組織rayとなる。篩管細胞と導管細胞に加えて維管束形成層は、放射柔組織細胞とも呼ばれ、茎の中で側方への物質輸送や、冬期の休眠時のような不適な環境にある時、物質を貯蔵する際の通り道となる柔細胞をも作る。放射柔組織細胞は、単列放射組織あるいは多裂放射組織となって、篩部・形成層・木部を貫く放射組織である。
維管束形成層の分裂により生じる細胞のうち、内側に形成されるのが、水や養分を土壌から植物体の様々な器官へ輸送するための二次木部secondary xylem(zάɪləm)に分化する。
晩材は、木材の一成長輪のうち、成長期の後期に形成される部分をいう。この時期に形成層の活動が衰え、針葉樹では細胞壁が厚く直径の小さな仮道管が、また広葉樹では細い道管がそれぞれ形成される。したがって、この部分は密度が高く、しばしば濃い色にみえる。温帯に生育する樹木では、晩材の形成される時期が夏季の後半にあたるため、夏材(かざい)(または「なつざい」)とも呼ばれる。また、秋材(しゅうざい)の呼称もあるが、形成時期を考えると、この語は不適当とされる。
温帯地域の木は、形成層の働きによる茎の肥大生長が年輪を形成する。春から夏にかけての生長期に形成される早材部分と、秋から冬にかけて色が濃く高密度に形成される晩材部分の、それぞれの細胞の大きさと細胞膜の厚さなどが異なるため、各1年間の肥大生長部分が明瞭に区別でき、これが同心円状の模様となる。 維管束形成層に由来する細胞のうち、外側へと移動した細胞は、二次篩部に分化する。二次篩部も光合成産物を葉から他の器官へ輸送する。二次篩部には篩部繊維phloem fiberが付随しており、これが他の繊維と同様に茎の強度を形成する。
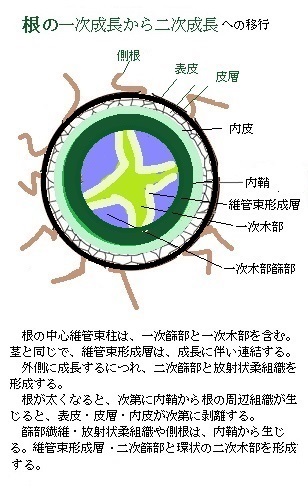 根の中心維管束柱は、一次篩部と一次木部、及び維管束形成層を備えている。維管束形成層は、茎と同様に成長に伴い連結され、外側に成長して二次篩部と放射状柔組織を形成する。それにより根が太くなるにつれ内鞘から根の周辺組織が生じるため、表皮・皮層・内皮は剥離する。
根の中心維管束柱は、一次篩部と一次木部、及び維管束形成層を備えている。維管束形成層は、茎と同様に成長に伴い連結され、外側に成長して二次篩部と放射状柔組織を形成する。それにより根が太くなるにつれ内鞘から根の周辺組織が生じるため、表皮・皮層・内皮は剥離する。裸子植物と多くの真正双子葉植物では、篩部組織や放射線柔組織、及び側根は内鞘細胞から生じ、維管束形成層は二次篩部と環状の二次木部を形成する。イネ科植物では、側根原基は篩部に接する内鞘細胞とその外側の内皮細胞で形成される。 大多数の植物では、内鞘細胞の伸長軸に垂直な分裂面での重層分裂に続いて、平行な分裂面での並層分裂が起きる。こうして側根原基細胞は、細胞分裂を重ね伸長をしながら、皮層と表皮の細胞列を貫き、新しい側根を外に伸ばす。
植物では、隣り合う細胞の細胞壁が中葉middle lamellaと呼ばれる薄い膜を介して接着されている。それで植物細胞の移動は起こりえないため、植物の発生は、細胞分裂と細胞成長のパターンに限定される。
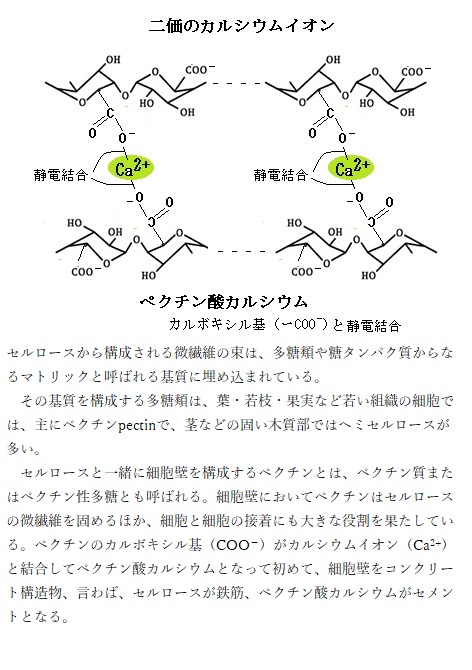
陸上植物の細胞には、セルロースとキシログルカンなどのヘミセルロースやペクチンの3つのグループからなる多糖類と構造タンパク質から構成される一次細胞壁primary cell wallと、セルロー スとキシランなどのヘミセルロース、そしてリグニンを主要な構成成分とする二次細胞壁secondary cell wallの2種類が存在する。一次細胞壁はまだ成長過程にある細胞膜の外側に生じ、通常、1µm未満の薄い構造で、二次細胞壁は、細胞の成長がほぼ終了した後に、維管束組織や表皮組織などの一次細胞壁の内側に沈着する。
一次細胞壁より分厚く強固で、陸上植物が地面に対して垂直に成長できる物理的な強度や、維管束組織における水の輸送と表皮組織における水分の損失防止と言った植物の陸上化・大型化に必要な機能を備えている。その頑丈さ耐久性は、堅い膠状(にかわじょう)の物質であるリグニンが、セルロースとヘミセルロースに架橋することで、その強度の根幹を保っている。
維管束は篩部と木部と言う大きな2つの部分からなる。木部の役割は植物の体を支え、水を根から葉へ送り、光合成産物の形成に貢献する。シダ植物と裸子植物では、仮導管がその役割を担い、被子植物では分厚い二次細胞壁と壁孔を持つ繊維細胞から成る「木部繊維細胞(木部を構成する細胞のうち、導管と柔組織を除く細長い細胞)」が、幹の強度を高め植物体を支持し、導管による水の吸い上げをたすける。木部繊維細胞が、パルプの原料となる。
 木部柔組織は、デンプンや油脂などの貯蔵組織であり、その細胞膜は、溶質の流入と流出に働くキャリアーcarrierでもある。裸子植物やシダ植物の木部は、その木部柔組織と仮導管だけから構成されるため、蒸散により駆動される水輸送と植物体の支持など両方を担わなくてはならず、そのため仮導管は大きな直径を持つことは無理であった。被子植物の木部の方は、それぞれの機能に特化した導管と木部繊維組織があるため、直径の大きな導管を作ることができることにより、多くの水輸送を可能にした。
木部柔組織は、デンプンや油脂などの貯蔵組織であり、その細胞膜は、溶質の流入と流出に働くキャリアーcarrierでもある。裸子植物やシダ植物の木部は、その木部柔組織と仮導管だけから構成されるため、蒸散により駆動される水輸送と植物体の支持など両方を担わなくてはならず、そのため仮導管は大きな直径を持つことは無理であった。被子植物の木部の方は、それぞれの機能に特化した導管と木部繊維組織があるため、直径の大きな導管を作ることができることにより、多くの水輸送を可能にした。木部を構成する多くの細胞は、導管が作られる過程で死んでしまう。導管や仮導管は、水の通路であるから縦に積み重なった細胞が死に、細胞内部の原形質がなくなり、上下の細胞壁が開孔し一本のパイプのようになる。陸上植物の茎や幹の内側には木部があり、外側には篩部が形成される。 篩部には、篩管があり、光合成産物である炭水化物を植物体中に送り届ける役割がある。篩管は生きた細胞であり、その細胞の上下に篩(ふるい)のような穴が多数開いており、それが篩管と言う名前の由来になった。
篩管を作る細胞は生きているが、核を持たない特殊な構造をしているため篩管細胞の隣に、篩管細胞が生きていくための必要なタンパク質を供給する伴細胞を伴う。
篩部輸送と木部輸送とに大別されるが、光合成産物の転流はもっぱら前者で行われるが、栄養塩類の転流は両者で行われる。生命活動に必須の元素のうち窒素・リン・硫黄・カリウム・カルシウム・マグネシウムのほか、鉄・マンガン・銅・亜鉛・モリブデンなどの元素は周囲の土壌や水に溶けた珪酸塩・硝酸塩・アンモニウム塩・リン酸塩などの形で摂取される。
これらの塩(アルカリと酸がくっついたもの)は、水溶液中ではイオンの状態に分かれて存在し、植物が栄養として吸収する時はこのイオンの形である。 代表的な硝酸塩である硝酸カルシウムは、普通の地下水の中では、水に良く溶けるので、地下水に溶けて電離して、プラスの電荷を帯びたカルシウムイオン(Ca2+)とCa2+の2倍量のマイナスの電荷を持った硝酸イオン(NO3-)と分かれてとして存在している。
細胞内のカルシウムイオン濃度は、細胞外に比べて殆どが低く抑えられている。外界から刺激を受けると、一転して細胞内のCa2+濃度が上昇し、それが情報を伝達するシグナルとなる。例えば、人間 が痛い、冷たいなどを感じた時、それぞれの感覚をつかさどる細胞内のCa2+の濃度の上昇がシグナルとなっている。 Ca2+は、主に二つの経路で細胞質へ運ばれている。一つが細胞の外側から運ばれる経路、もう一つが、細胞内に存在する小器官である小胞体や液胞から遊離する経路である。
小胞体や液胞は脂質でできた膜に包まれており、細胞内カルシウムイオン・ストアとも呼ば れている。細胞内小器官の一部は、Ca2+をその内腔(細胞小器官のさらに内側の空間)に貯蔵し一定の条件下でカルシウムイオンを細胞質に放出する。そのCa2+貯蔵庫および供給源としてカルシウムシグナルに関与する細胞内小器官を、細胞内カルシウムストアと総称する。
 フォトトロピンは、青色光を吸収する植物の青色光受容体の一つで、シロイヌナズナにはフォトトロピン1(phot1)とフォトトロピン2(phot2)の二つが存在することが知られている。phot1とphot2
ともシロイヌナズナの青色光による光屈性・葉緑体定位運動・気孔の開口をひき起こす。いずれも植物が様々な光環境下に適応するための重要な反応である。
フォトトロピンは、青色光を吸収する植物の青色光受容体の一つで、シロイヌナズナにはフォトトロピン1(phot1)とフォトトロピン2(phot2)の二つが存在することが知られている。phot1とphot2
ともシロイヌナズナの青色光による光屈性・葉緑体定位運動・気孔の開口をひき起こす。いずれも植物が様々な光環境下に適応するための重要な反応である。
Ca2+がphot1とphot2に共通するシグナルであるが、phot1とphot2 によるCa2+運搬機構は、それぞれ異なっていることが明らかになっている。
篩部は複合組織であり、植物全体へ糖や他の有機化合物の輸送を担う細胞全体を篩要素sive eelement(sív;ふるい、こしき)と呼ぶ。それは、複数の異なる形態を含む幅広い細胞を含み、典型的な被子植物に見られる高度に分化した篩管要素sievetube elementや、裸子植物の比較的特化していない篩細胞sieve cellをも含めて篩要素と言う。篩要素に加えて、伴細胞companioncellと柔細胞parenchyma cellがある。
篩要素は、成熟すると核をふくむ原形質のほとんどを失い、互いに連なって篩管sieve tubeを形成し、転流物質の流路として機能する。篩管の両端部分の細胞壁には篩孔sievepore(pˈɔɚ)と呼ぶ多数の小孔をもった篩板(しばん)sieve plateがあって、その孔の開いた仕切りと言える細胞壁は、篩い(ふるい)状になっており、物質が透過できる様になり、隣接する篩管同士と連絡している。つまり、細長い篩管細胞が縦方向につながり、細胞の中身がつながった形をしている。
篩管要素には、1つもしくは複数の伴細胞が隣接している。実は、単一の母細胞の細胞分裂により、篩管要素と伴細胞に分化する。裸子植物には、伴細胞がない代わりにタンパク細胞albuminous cell(albuminous;ælbjúːmənəsとはタンパク質を含むの意味)と言うものがあって伴細胞に似た働きをしていると考えられている。 篩管細胞の中に僅かに残る原形質は、細胞の側面に沿って存在し、物の輸送を妨げないと言う。それでも10年、100年と生きている。細胞が生きて行くのには、核の遺伝子の遺伝情報を解読して作られたタンパク質の存在が不可欠で、核のない篩管細胞では、このタンパク質は作れない。そのため、必要なタンパク質は、隣接する伴細胞で作られて、篩管細胞に送られているようだ。
篩管細胞と伴細胞では、たくさんの小さな孔が細胞壁を貫いている。多くの場合、伴細胞の方が、複雑に枝分かれしている構造になっている。この原形質連絡の多数の繋がりは、この2つの細胞の機能的関係を物語っている。
伴細胞で作られたタンパク質はこの原形質連絡を通って、タンパク質を作れない篩管細胞に運ばれている。さらに、成熟葉の光合成産物を合成する葉肉細胞(ようにくー)から、その葉の小維管束の篩要素への光合成産物の輸送にも、重要な役割を果たしている。葉の葉肉細胞で合成された光合成産物は直接篩管の中には入れず、先ず伴細胞に入り、そこから、これも原形質連絡を通って篩管の中に送り込まれている。
生合成能力を失った篩管細胞が、必要とするタンパク質合成や様々な代謝機能などを、伴細胞が、肩代りしていると考えられている。さらに、伴細胞の多くのミトコンドリアが、篩要素へATP形態のエネルギーを供給しているようだ。
柔細胞は生きている細胞で、原形質を有している。有機養分の貯蔵組織や分泌組織などを主に構成し、養分を合成・分解・貯蔵し放出する。細胞壁は一次細胞壁から成り、薄いのが一般的である。篩部柔細胞は、二次篩部の場合には二次木部と同様に、縦に長い細胞からなる紡錘形柔組織と等径的な細胞からなる放射組織からなる。
輸送細胞transfer cellは、伴細胞に似ているが、指のような突起がある細胞壁を複数持ち、細胞内部に向けて肥厚する点が異なっている。この肥厚は、篩要素と接しない部位の細胞壁で、特に目立つことから、細胞膜の表面積を増大させ、アポプラストを経由して膜を横切る溶質が、細胞膜を通過できる輸送能を高めていると見られる。
 輸送細胞と篩要素の間には、原形質連絡があるが、輸送細胞と周辺の篩要素以外の細胞の間には、原形質連絡は稀にしか見られない。篩要素と輸送細胞のシンプラストは、周辺細胞のシンプラストとは完全ではないが、ほぼ隔離状態にある。
輸送細胞と篩要素の間には、原形質連絡があるが、輸送細胞と周辺の篩要素以外の細胞の間には、原形質連絡は稀にしか見られない。篩要素と輸送細胞のシンプラストは、周辺細胞のシンプラストとは完全ではないが、ほぼ隔離状態にある。輸送細胞は、篩部転流径路上にある節、ソース組織の篩部、篩要素以降の積み下ろし径路でよく見られる。
木部柔細胞xylem(zάɪləm) parenchyma cellは、輸送細胞に変化することがある。アポプラストの一部である木部を移行する溶質を吸収し、その輸送先を変えるためと考えられている。
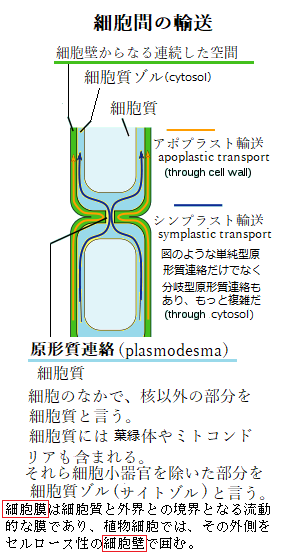
アポプラストは、植物組織において細胞膜よりも外側の細胞壁や細胞間の空間の総体、及び死んだ細胞の内腔、例えば木部道管や道管繊維などが連続した系で、その径路で、水が根の皮層を横切る時、膜を横断する必要がなく、連続して移動できる領域を言う。言わば、水溶液(アポプラスト液)で満たされた空間の総体で、アポプラスト輸送apoplast transportとは、アポプラストと呼ばれる「細胞壁からなる連続した空間」を通る分子の移動のことである。隣接した細胞群の互いに繋がった細胞壁を通す物質の移動になり、分子は、細胞膜を横切ることなく植物体内を移動できる。通常、植物組織の全体積の5~20%が細胞壁である。植物体内におけるアポプラストは、水や溶質の輸送路であり生化学反応の場でもある。特にその細胞壁は単なる物質の移動経路としてのみならず、物質代謝の場としても機能している。 アポプラストは、植食性昆虫や動物による食害に、全身的に抵抗する。その物理的損傷に応答して、植物組織の障壁を分解する加水分解酵素が植物側から放出される。このような酵素による生成物の殆どは、アポプラストに存在しており、damage associated molecular pattern(DAMP;損傷関連分子パターン)と呼ばれる内生のエリシターelicitor(自然免疫を活性化させる自己生成する誘導因子)として機能して、広範囲の生物に対する抵抗性を誘導する。 成長中の果実や生育地が高塩環境であれば、アポプラストの溶質濃度が、高いにもかかわらず、アポプラストの溶質ポテンシャルは、-0.1~-0MPaの範囲にあることが多い。
シンプラストsymplastとは、アポプラストと対をなす輸送路である。植物細胞の細胞膜内で、隣接した細胞同士を繋ぐ、直径40~50nmの管により高頻度で連結されている連続体を言う。細胞質ゾル(サイトゾル)同士は、分岐した原形質連絡や、枝分かれのない原形質連絡によって、植物細胞間では、複数にわたる繋がりが、それぞれ連続体を形成している。 シンプラストにより、細胞間の低分子輸送をはじめ、溶質の移動が、拡散によって容易に行われると考えられている。それをシンプラスト輸送symplastictransportと呼ぶ。その原形質連絡は、タンパク質や核酸など、様々な巨大分子の輸送経路となり、隣接する細胞間の情報交換を仲介している。 アポプラスト輸送とシンプラスト輸送は、いずれも維管束系で重要な役割を担っている。
目次へ
 体細胞はもとより、いずれは配偶子となる生殖系列細胞も含めて二倍体である。しかも、母親由来の染色体と父親由来の染色体が1セットずつ、計2セットを持っている。
体細胞はもとより、いずれは配偶子となる生殖系列細胞も含めて二倍体である。しかも、母親由来の染色体と父親由来の染色体が1セットずつ、計2セットを持っている。二倍体生物の有性生殖に直接かかわる生殖細胞(germ cell)、即ち配偶子(gamete;gǽmi t)は、殆どの細胞と異なり一倍体であるため、ヒトであれば、各配偶子には23本の染色体が1組あるだけだ。しかも、殆どの生物は、雄と雌で異なる新たな遺伝子を含む配偶子を作り上げている。動物の配偶子は、大型で運動性のない卵と、小型であるが活発に運動する精子がある。その違いは、卵が大量の細胞質を抱えているが、精子にはそれが殆どないことによる。この特異な2種類の一倍体である配偶子が、融合して二倍体の細胞の受精卵、即ち接合子(zygote;zái out)となり、やがて新たな遺伝子を受け継ぐ個体を誕生させる。そのため2種類の配偶子は、染色体を正確に半分に分ける特殊な細胞分裂をして作られる。このような細胞分裂を減数分裂と言う。
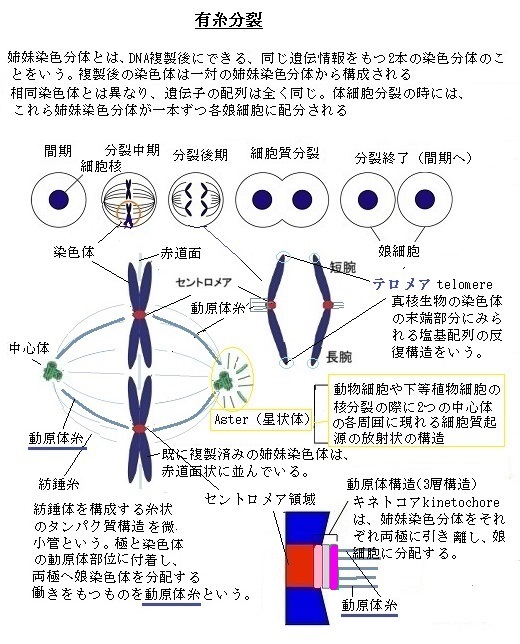 減数分裂では、1回のDNA複製に続いて2回の細胞分裂が起こる。二倍体細胞が通常の細胞分裂で分裂する場合は、持っている2セットの染色体を前もって2倍にする。これにより、その細胞分裂で新たに誕生する2つの娘細胞それぞれが、完全な母方染色体1セットと父方染色体1セットからなる、ヒトの染色体セットを受け継ぐことになる。
減数分裂では、1回のDNA複製に続いて2回の細胞分裂が起こる。二倍体細胞が通常の細胞分裂で分裂する場合は、持っている2セットの染色体を前もって2倍にする。これにより、その細胞分裂で新たに誕生する2つの娘細胞それぞれが、完全な母方染色体1セットと父方染色体1セットからなる、ヒトの染色体セットを受け継ぐことになる。減数分裂では、通常2セット分ある全染色体が、最終的に1セットになり、それが精子と卵からなる配偶子を作る。その前に起こる染色体倍化の現象は、通常の細胞と同様で1回である。
生殖系列細胞は、その後独自の過程を経る。DNAの複製を行なうことなく、連続して2回の細胞分裂が起き、むしろ最終的には、細胞内の染色体数を半減させて4個の一倍体細胞である配偶子を作る。
生物40億年の進化の歴史は、苛烈な自然選択に、無駄なく合理的に応えられる生命体にしか、その存続を認めなかったことを物語る。それ以後の種の存続のためにも、不可欠な突然変異が必ず生じる仕組みを、進化を担保する原動力としている。
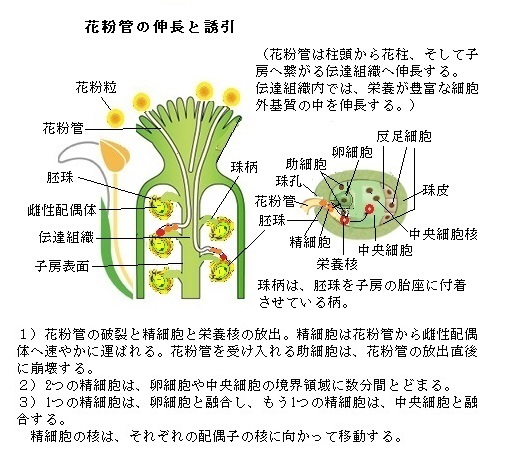
被子植物の花粉形成過程では数多くの遺伝子が発現することが、近年の遺伝子発現解析によって明らかになた。ゲノム解析、マイクロアレイMicroarray法(アレイ Arrayとは整列とか配列の意。微小なガラスチップの上にDNAの塩基配列を合成し貼り付け、多量の生物学的データを構築する技術)やRNA-Seq(sequencing塩基配列決定)法(RNA分子の遺伝子配列における変異、即ち一塩基多型Single Nucleotide Polymorphism、即ちゲノムの塩基配列の中の同一部位に2通りの塩基配列があり、最も多いのが塩基1個の置換であり、これをSNPと呼ぶ)・短い欠失・挿入変異など、発現変動を網羅的かつ定量的に解析するなど遺伝子発現解析技術の進展により、植物が持つ約3万個以上の遺伝子の約半数が葯や花粉で発現し、さらにその10分 の1程度は花粉特異的に発現すると見られている。例えば、イネが持つ遺伝子総数3万2千個のうち、約2万個もの遺伝子が発現して花粉と葯で働いている。イネの花粉研究に関しては、農業上重要な形質である種子不稔を示す突然変異体を解析することにより、主に葯壁で発現する遺伝子の情報が蓄積されてきている。
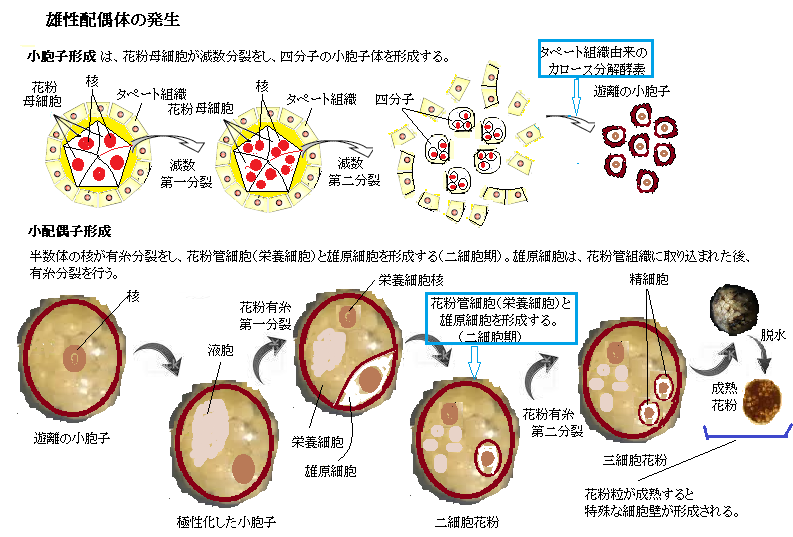
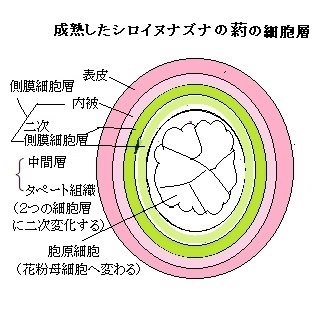 よく晴れた風の強い日に、雄しべから小さくて軽い花粉が大量に放出され、あたり一面に散布する。 その前に、雄しべの先端にある葯の中で細胞が分裂を繰り返し、多数の「花粉母細胞」が作られる。その「花粉母細胞」は、酵素の作用で2度行なう減数分裂によって、「花粉四分子」と呼ばれる4つの未熟な花粉・小胞子になる。
やがて 小胞子の核は細胞の一端に移動し、その場で半数性の体細胞分裂(小胞子分裂)をおこなう。この分裂は大きさの異なる2つの細胞を生じる典型的な不等細胞分裂であり、大細胞は栄養細胞(花粉管細胞ともよぶ)、小細胞は雄原細胞である。やがて雄原細胞は栄養細胞質中に入り、その細胞内に細胞膜も備えた細胞になる。雄原細胞は細胞分裂をおこない、2つの精細胞が生じ、被子植物特有の重複受精にいたる。イネやシロイヌナズナでは、雄原細胞分裂が開花前の花粉内で起こるが、ユリやタバコなどでは、精細胞の形成は受粉後の花粉管中で起こる。
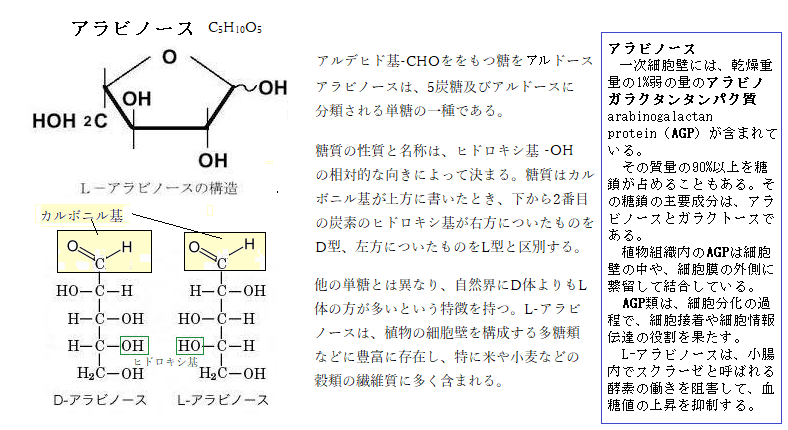
イネで働くトランスポゾンtransposon(ゲノム上の位置を転移する塩基配列で、動く遺伝子、転移因子を言う)と呼ぶ遺伝子(Tos17)を活性化させると、染色体上にランダムに転移し、転移した場所の遺伝子が分断され、突然変異体を生じる。Tos17 の転移による遺伝子破壊が花粉変異の原因であることをつきとめ、関連する遺伝子を同定すると、そのうち1つは、葯で優先的に発現する遺伝子であり、細胞壁成分のアラビノースarabinoseをリン酸化する新規のアラビノキナーゼarabinokinaseをコードしていた。
よく晴れた風の強い日に、雄しべから小さくて軽い花粉が大量に放出され、あたり一面に散布する。 その前に、雄しべの先端にある葯の中で細胞が分裂を繰り返し、多数の「花粉母細胞」が作られる。その「花粉母細胞」は、酵素の作用で2度行なう減数分裂によって、「花粉四分子」と呼ばれる4つの未熟な花粉・小胞子になる。
やがて 小胞子の核は細胞の一端に移動し、その場で半数性の体細胞分裂(小胞子分裂)をおこなう。この分裂は大きさの異なる2つの細胞を生じる典型的な不等細胞分裂であり、大細胞は栄養細胞(花粉管細胞ともよぶ)、小細胞は雄原細胞である。やがて雄原細胞は栄養細胞質中に入り、その細胞内に細胞膜も備えた細胞になる。雄原細胞は細胞分裂をおこない、2つの精細胞が生じ、被子植物特有の重複受精にいたる。イネやシロイヌナズナでは、雄原細胞分裂が開花前の花粉内で起こるが、ユリやタバコなどでは、精細胞の形成は受粉後の花粉管中で起こる。
イネで働くトランスポゾンtransposon(ゲノム上の位置を転移する塩基配列で、動く遺伝子、転移因子を言う)と呼ぶ遺伝子(Tos17)を活性化させると、染色体上にランダムに転移し、転移した場所の遺伝子が分断され、突然変異体を生じる。Tos17 の転移による遺伝子破壊が花粉変異の原因であることをつきとめ、関連する遺伝子を同定すると、そのうち1つは、葯で優先的に発現する遺伝子であり、細胞壁成分のアラビノースarabinoseをリン酸化する新規のアラビノキナーゼarabinokinaseをコードしていた。花粉形成過程では2度の細胞分裂がおこるなど、花粉が生長する際に細胞壁の代謝が活発に起こるため、この酵素遺伝子が重要な働きをしていると考えられる。イネ以外の多くの植物でも花粉形成に関わる類似のアラビノキナーゼが存在していると思える。
キナーゼkinaseは、リン酸化を触媒する酵素で、タンパク質などの基質にATPに由来するリン酸を付加するすることにより、その分子の性質を変える。その上、あるタンパク質が受容体の機能を備え、シグナルを受容するとそれが信号となる。他の分子をリン酸化することにより、他の分子に伝達される。そのタンパク質を受容体キナーゼreceptorkinaseと呼ぶ。
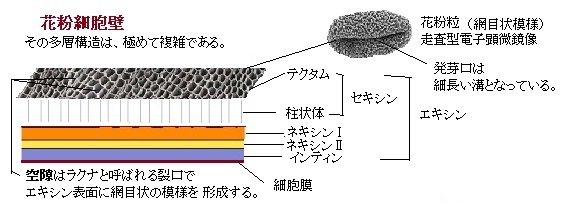 植物が持つ約3万個の遺伝子の約半数が葯や花粉で発現し、さらにその十分 の一程度は、花粉だけに特定され発現されている。イネの花粉突然変異体collapsed
abnormal pollen1 (cap1)の開花期の変異花粉は、細胞質成分の殆どを失い、花粉外壁exinのみになる。花粉変異の原因遺伝子CAP1は、アミノ酸からなるL-アラビノキナーゼ様タンパク質をコードしている。正常なイネでは、CAP1遺伝子は花粉と葯の内壁で発現し、花粉形成の二細胞期、即ち花粉管細胞(栄養細胞)と雄原細胞が形成される際が、最もその遺伝子の発現量が高くなる。イ ネの花粉形成では、その二細胞期にデンプンの蓄積が始まり、開花期の成熟した花粉の花粉管細胞質はデンプン粒で満たされている。
植物が持つ約3万個の遺伝子の約半数が葯や花粉で発現し、さらにその十分 の一程度は、花粉だけに特定され発現されている。イネの花粉突然変異体collapsed
abnormal pollen1 (cap1)の開花期の変異花粉は、細胞質成分の殆どを失い、花粉外壁exinのみになる。花粉変異の原因遺伝子CAP1は、アミノ酸からなるL-アラビノキナーゼ様タンパク質をコードしている。正常なイネでは、CAP1遺伝子は花粉と葯の内壁で発現し、花粉形成の二細胞期、即ち花粉管細胞(栄養細胞)と雄原細胞が形成される際が、最もその遺伝子の発現量が高くなる。イ ネの花粉形成では、その二細胞期にデンプンの蓄積が始まり、開花期の成熟した花粉の花粉管細胞質はデンプン粒で満たされている。アラビノースは、5炭糖及びアルドースに分類される単糖の一種である。 他の単糖とは異なり、自然界にD体よりもL体の方が多いという特徴を持つ。細胞壁などに結合型で存在する。単糖とはヒドロキシ基-OHを2個以上と、アルデヒド基-CHO、あるいはケトン基-CO-のいずれかをもつ化合物である、アルデヒド基とケトン基をまとめてカルボニル基と呼ぶ。アルデヒド基をもつ糖をアルドース、ケトン基をもつ糖をケトースと総称する。組成式はC5H10O5。
糖質の性質と名称は、ヒドロキシ基の相対的な向きによって決まる。糖質はカルボニル基を上方に書いたとき、下から2番目の炭素のヒドロキシ基が右方についたものをD型、左方についたものをL型と区別する。他の単糖とは異なり、自然界にD体よりもL体の方が多いという特徴を持つ。L-アラビノースは、植物の細胞壁を構成する多糖類などに豊富に存在し、特に米や小麦などの穀類の繊維質に多く含まれる。
目次へ
三次元的に整然と細胞が配置される雌性配偶体のパターン形成は、核の位置が重要である。
葉は、構造的には、左右相称であり、基部・先端部軸及び向背軸(表・裏軸)という軸性を保持する器官である。シロイヌナズナの AS2 遺伝子は、これら3つの基本骨格にそった葉の形成に必要である。この遺伝子に変異が生じると、基本骨格にそった形態形成が、すべて不全となる。AS2 タンパク質は、42個のタンパク質、ASL1~ASL 41を含む 植物に特有なAS2/LOB ファミリーに属する。トウモロコシのAS2/LOBファミリータンパク質をコードする遺伝子の変異体では、胚嚢において核および細胞の数が増加する。
植物ホルモンとしてオーキシンauxin・サイトカイニン・ジベレリン・アブシジン酸など10種類程度しか知られていない。植物がこの少ない数のホルモンを駆使して、個体の秩序を保っている。
オーキシンの働きは複雑で、植物では、各器官において、その成長に対するオーキシンの最適濃度(感受性)が異なっている。茎は、オーキシンが比較的高濃度な状態で成長が促進される。そのためか、根は茎の最適濃度においては成長が抑制され、低濃度で成長の促進がみられる。同様に、頂芽と側芽においてもオーキシンに対する最適濃度が異なり、頂芽の成長が促進されるオーキシン濃度の状態では、側芽の成長が抑制される。それにより頂芽優勢がおこると考えられている。側芽におけるオーキシンの最適濃度は、頂芽のそれよりも、もっと低い。
そのオーキシンの合成類似物であるモルフォゲンmorphogenと呼ばれる分子の濃度勾配は、シロイヌナズナの雌性配偶体において、その勾配に応じ細胞が分化し、その位置取りが行われているようだ。モルフォゲンは、位置情報を与える上で鍵となる役割を果たす。モルフォゲンの合成・輸送・分解の組み合わせにより、モルフォゲン分子は、組織内で濃度勾配を形成して分布する。それにより一連の濃度依存的な反応を引き起こす。多くの器官では、発生・変態・再生の際に、局在する発生源からモルフォゲン分子の濃度勾配を位置情報として発し、その濃度勾配に依存して細胞の発生が決定されている。というのも、オーキシンは、細胞から隣の細胞へエネルギー依存的に極性を持って輸送される唯一の植物ホルモンであるからだ。その細胞間のオーキシンの極性輸送は、シンプラストを介することなく、その過程全体で代謝エネルギーが必要であるが、オーキシンは細胞膜から出て細胞壁に広まり、隣の細胞膜を通って隣の細胞に入る。
オーキシンは植物の細胞分裂・形態形成・屈性現象など、様々な生理現象の制御にかかわる低分子ホルモンである。もっとも有名で、かつ重要な植物ホルモンである。近年では、オーキシンの細胞外への排出輸送体である「PIN」の研究が進み、オー 胞の外へ汲み出す。この繰り返しによりオーキシンは一定の方向へ運ばれる。
オーキシンは「PIN」のある側に多く流れ、その先でオーキシン濃度が高くなることから、オーキシンの輸送方向の決定には「PIN」が重要な役割を果たしているようだ。茎や根で見られる屈性は、オーキシンの「極性輸送」によって茎や根の内部のオーキシン濃度に勾配が生じ、その結果、成長速度が左右・上下に差が生じることで起きる現象である。
オーキシンの「極性輸送」は、重力の影響を受けない。しかし植物は重力を感じて曲がる性質も持っている。根は重力に導かれ下へ下へと伸びる。茎は重力と反対方向へ立ち上がる。この植物が重力を感知し屈曲する仕組みを「重力屈性」という。
オーキシンは、形成途中の胚嚢の内部に濃度勾配を形成していることや、オーキシン合成、あるいはオーキシン応答にかかわる遺伝子発現を胚嚢において増減させると卵細胞のマーカーが発現するようになる。
多細胞動物の個体は、様々な種類の細胞で構成されている。例えば、ヒトでは、60兆の細胞に分化した内臓・神経・血球・皮膚など、形も機能も異なった体細胞が存在する。これらの細胞は全て一個の受精卵から派生し、細胞分裂を繰り返して、当初は同じ性質を持っていたはずが、分化して専門化した形態と機能を備えた細胞になる。この過程は通常では後戻りができない。
多細胞植物の細胞も、受精卵から分化するが、体細胞を適当な条件で培養すると、初期化のような現象が起こり(脱分化)、細胞分裂を経てもう一度分化させ(再分化)、さらに個体を再形成することもできる。植物細胞の持つこの能力を分化全能性と言う。
動物の受精卵は分化を繰り返して成長し、最初は同じ性質を持っていたはずの細胞が数を増やすにつれ特殊化して様々な性質の細胞になる。この分裂成長の各段階で細胞核に遺伝子(DNA)の特定の部位に、置換基などが結合する修飾などにより、ある特定の遺伝子しか働かなくなる。分化とは、遺伝子の発現が特定化される過程と言える。これは原則として一方通行のプロセスであるが、植物は、動物よりリセットされやすい。なお、この脱分化という分化のリセットによる初期化は、遺伝子群の発現を制御することにより起こる。また、再分化に関しても、同様に遺伝子機能によって行われる。
分化全能性totipotencyとは、個体を構成する様々な種類の細胞のいずれも分化することができる潜在能力を言う。動物でも植物でも、すべての細胞の起源となる受精卵は、明らかに分化全能性を持っている。次の受精卵を生み出す生殖系列の細胞も、分化全能性を保持している。
それ以外の体細胞の分化全能性であるが、植物においては、体細胞が分化しても、必ずしも分化全能性は失われない。1960年代には、数々の細胞培養実験の結果から、既に明確になっていた。
例えば、葉・茎・根・花などの一部を取り出して、ある条件下で無菌的に培養すると、それらの組織を構成している細胞が分裂をはじめ細胞が増殖する。これらの細胞はもとの分化した細胞ではなく、未分化の状態のもので、ただ細胞の塊のようなもので、これをカルスと呼んだ。このように一度分化した細胞が未分化の状態になることを「脱分化」と言う。
通常の「胚」は配偶子(精細胞と卵細胞)が受精してできた接合子(受精卵)が分裂を繰り返して形成され、種子の中に納まる。「不定胚」は、その植物体の体細胞が、接合子と同じような過程を経ず、直接に形成され、胚と同等のものになる。「不定胚」からも、培養によって完全な個体にまで育てることができる。カルスと「不定胚」の基本的な違いは、前者は未分化のものであり、後者は分化したものである。
例えば、ニンジンの細胞から不定胚を経て個体の全部を再生させた実験などで、植物体細胞が分化全能性を持つことが証明されている。分化全能性のより厳密な証明は、その後タバコ葉肉プロトプラストprotoplast(細胞壁を酵素処理により取り除いた細胞)から植物体を再生することによってなされた。「proto-」とは「最初の」「主要な」「原始の」、「plast」とは「形成されたもの」(連結形)。
プロトプラスト融合protoplast fusionは、異なるゲノム情報を持つ2つの植物細胞を融合させる方法である。植物の病原菌である「アグロバクテリウム」は、この細菌に感染した植物の成長ホルモンを過剰に生産させ、クラウンゴールcrown gallと呼ばれる瘤状の腫瘍を作らせ、果樹などの農作物の収量を減少させる深刻な問題を起こす。その一方では、「アグロバクテリウム」が持つ遺伝子の一部により、植物細胞の形質を転換できる。改変した「アグロバクテリウム」は、植物に感染させることもできる。葉の小片を植物体から切り離し「アグロバクテリウム」の培養液で共培養し、その後、植物細胞を洗浄して組織培養のための培地上で培養する。植物ホルモンのオーキシンとサイトカイニンを用いて、それぞれの培養組織から根とシュートを生じさせる。この方法で、成長した植物体が作られる。現在、栽培されている普通小麦 Triticum aestivumは、3つの異なる先祖種間の他家受粉による異種6倍体で、古典的なランダムな遺伝的組換えに依存しながらも、全く新たな植物種を.生み出していた。
裸子植物と被子植物を合わせて「種子植物」と呼ぶ。種子を作るということは植物が持つ遺伝的な継承である。通常、植物体は母方由来と父方由来の2種の染色体を1セットにする。それで2倍体と言う。植物の成長の大前提にあるのが、細胞分裂でDNAの複製を伴う。細胞分裂では、1セットの染色体が倍加し、その同じ遺伝情報を持つ染色体が倍化する。それを姉妹染色分体と呼ぶが、細胞分裂後期には、正確に2等分される。したがって、娘細胞の染色体セットは2倍体のまま変わらない。しかし、自然の条件でも頻度は少ないが、発生の初期に倍加した染色体セットが2等分されないで4本の染色体をもった細胞ができ、これが植物体になることがある。それが4倍体であるが、植物種としては同一で、4倍体植物はその後も正常な減数分裂をし、受精し種子形成をしても、細胞分裂の基本形態は変わらないため、次世代にも4倍体が保たれる。
ところが、2倍体植物と4倍体植物とが交配すると2倍体植物の卵子・精子(1倍体)と4倍体植物の精子・卵子(2倍体)とが融合するので3倍体となる。それでも、通常通りの細胞分裂が行われ、3倍体の植物体は正常通り成長し花を咲かせる。しかも、植物種としても同一のままである。ところが3倍体では、卵子・精子形成の際に行われる減数分裂が正常に行われないので、正常な配偶子ができない。そのため種子を形成するための受精・胚発生ができないため種子は生じない。
コムギの品種は、環境特異的で、もっとも広く利用されている品種でさえも、特定の地理的かつ自然的な環境に適応するため、その潜在能力の殆どは、それまでの栽培慣行に全面的に依拠するようになる。2倍体のコムギ種は14本の染色体がある。4倍体のフタツブコムギ(現在のパン小麦の原種にあたる古代小麦)や現代のデュラムコムギ種(マカロニコムギとも呼ばれ、マカロニやスパゲッティなどに合う原料としてパンコムギに次ぐ生産量をあげている。タンパク質含量が高い)は28本の染色体を有している。4倍体のコムギ種は、2つの2倍体コムギ種間の稀に生じる自然交雑によってできた種である。また、一般的な6倍体の普通小麦は42本の染色体を有している。即ち、ゲノム2倍体とゲノム4倍体が組み合わされて、ゲノム6倍体雑種ができあがった。一般的な6倍体のパンコムギ種も、42本の染色体を有している。
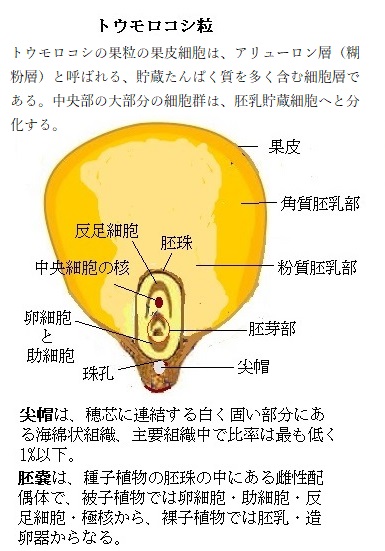 様々な生物のゲノムを解析する際に重要なことは、DNAの長さと、その中にある遺伝子の数である。今日のトウモロコシの品種は、中央アメリカに分布しているイネ科Zea属のテオシントとして知られる近縁野生種が、数百年にわたり選抜育種され栽培化されたものの子孫である。育種と栽培化によりトウモロコシの形態は大きく変化し、果実や果粒も大きくなり味もよく、何よりも格段に糖度が増した。
様々な生物のゲノムを解析する際に重要なことは、DNAの長さと、その中にある遺伝子の数である。今日のトウモロコシの品種は、中央アメリカに分布しているイネ科Zea属のテオシントとして知られる近縁野生種が、数百年にわたり選抜育種され栽培化されたものの子孫である。育種と栽培化によりトウモロコシの形態は大きく変化し、果実や果粒も大きくなり味もよく、何よりも格段に糖度が増した。古典的な栽培育種では、2つの品種を他家受粉し、その子孫から望ましい形質を持つ株を選抜し、その系統を生育させる。その積み重ねを継続する過程で、折角の両親の優れた遺伝的寄与も減数分裂で混ざり合うため、その望ましくない形質を取り除くため、選抜された系統間で反復して「戻し交配backcross」を長い期間行わなければならない。
アグロバクテリウムを介した形質転換では、望ましい遺伝子だけを目的の植物に導入する。その植物を、遺伝的に改変された生物GMO(genetically modified organism)と呼ぶ。
GMO供与遺伝子は、交配が不可能な受け取り側にも様々な遺伝的要素として繋ぐことができ、それにより遺伝子に新たな利用法が付加されるようになった。Modifiedとは「修正された」と言う意味である。ゲノムに入ったGMO供与遺伝子を、導入遺伝子transgeneと呼ぶ。
通常、遺伝子の機能を調べるには、正常遺伝子を改変遺伝子で置き換え、正常タンパク質が欠けたままの状態で、変異タンパク質の機能を調べる。最初に標的遺伝子を置き換えてから、その細胞を発生過程にある細胞の胚に移植すると変異マウスができるようになる。そのマウスから生じる異常障害から、正常タンパク質の役割が推測される。
そのために先ずは、ES細胞を培養して、遺伝子操作で改変した遺伝子のDNA断片を、その多数のES細胞に導入する。ごく少数のES細胞に限られるが、相同組換えにより、正常な遺伝子が改変遺伝子に置き換えられた細胞が生まれる。この少数の細胞を同定して培養し、コロニーを作らせる。この一対の標的遺伝子の内、一方だけが、改変遺伝子に置き換わったES細胞になる。このES細胞を、マウスのごく初期の胚に注入すると、ES細胞は、成長中の胚に取り込まれ、次に生まれた世代のマウスの体細胞の一部が、改変遺伝子を持つようになる。それにより生殖系列細胞にも改変遺伝子を持つマウスが稀に生まれ、それらを正常マウスと交配すると、その子孫の中に、生殖系列細胞も含んだ全細胞が改変遺伝子を持つマウスが生まれる。これらをノックインマウスknock in mouseと呼ぶ。このノックインマウス同士の交配で、生体中の細胞に、一対の各染色体双方に改変遺伝子を持つ子孫が得られる可能性が起こる。これにより、疾病と遺伝子の関係や、特定のタンパク質や酵素の生成が生体に与える影響など、細胞や生体における遺伝子の機能解析が強力に進めるようになった。
一般には、初期発生の間に個々の動物細胞の分化能力は次第に限定され、分化全能性は失われていくと見る。しかし近年では、動物細胞の分化全能性が維持もしくは喪失される際の分子機構が解析され、分化全能性の研究は、既に、植物分野を超え、特に再生医療を見据えた、今後の胚性幹細胞embryonic stem cells(ES細胞)の分化全能性や多分化能の研究成果が、大いに期待されるにようになった。
「embryonic」とは「胎生の」、「stem cell」は「幹細胞」の意味、「幹細胞」とは、自分とまったく同じ能力を持った細胞に分裂することができる能力(自己複製能)と様々な細胞に分化する能力(多分化能)を持つ特殊な細胞である。
ES細胞は、多能性細胞とも呼ばれる。受精卵が初期に分裂してできる100個程度の細胞の塊をばらばらにほぐして、体外で培養を続けることによって作られる。ただ、受精卵を壊すという倫理的問題や免疫拒絶が避けられないという技術的問題で臨床には利用しにくい。この難点を回避しようと、山中教授は体細胞を初期化して、多能性を持つ細胞を作る技術を開発した。皮膚など成体の細胞に、いくつかの遺伝子を人工的に導入して作った多能性幹細胞induced Pluripotent Stem cellは、iPS細胞と略される。培養して増殖させることができ、適当な因子を加えることなどで体を構成する殆どの細胞ができるため、この名がある。「Pluripotent(plùːrípətənt)」とは「多能性」の意味。
植物の分化全能性の研究も、植物分子生物学の恩恵を受け、この10数年の間に著しく進展し新たな実用段階に入っている。
目次へ
イネ科植物の在来品種の場合、一定以上の肥料を投入すると収量が絶対的に低下する。それは在来品種の場合、丈が伸びすぎて倒伏が起こり、肥料の増投が収量の増加に結びつかないからである。
背が伸びない「矮性」は、自然界で競合する植物にとって、光合成に致命的な不利となるが、栽培品種としては、むしろ有利に働く。背が低いと倒れにくい、更に収量が増える。丈が伸びれば、そのために多くの栄養とエネルギーが消費される。
「矮性」の品種では、栄養やエネルギーを果実や種子の方に集中して投入でき、通常の品種より収穫量が増える。この「矮性」の利点は大きく、栽培作物では、背を低くするため「矮性剤」を使う。その「矮性剤」の実体は、植物ホルモンのシベレリンという生合成阻害剤である。
「ジベレリン」の主な働きは、茎の背丈を伸ばすことである。しかし、ブドウ農家にとっても「ジベレリン」とは、ブドウを種無しにし、ブドウの粒を大きくする薬品でもある。ブドウの花が咲いた頃、コップに注いだ「ジベレリン」の溶液にぶどうの花を一房一房に浸してゆく作業を続ける。一般的に、この作業を「ジベレリン処理」と言う。ブドウ農家にとって、この処理は、ブドウを種無しにし、ブドウの粒を大きくする重要な作業になる。
1,985年、イギリスの2人の研究者が、植物の種子にジベレリンが含まれていることを発見し、それが皮切りに、様々な植物が「シベレリン」を生成していることが明らかになり、その結果、「ジベレリン」は植物ホルモンであることが分かった。「ジベレリン」を生成する際に重要な働きをする遺伝子や「ジベレリン」が植物の体内で生合成される径路など、詳細に解明されている。
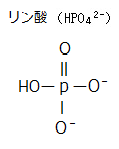 植物の情報伝達の迅速な応答は、タンパク質のリン酸化を介して行われる。葉緑体や色素体では、特異的なキナーゼが、ATPのリン酸基を、デンプン代謝に関与する酵素の特定アミノ酸であるα-アミラーゼに転移させている。
植物の情報伝達の迅速な応答は、タンパク質のリン酸化を介して行われる。葉緑体や色素体では、特異的なキナーゼが、ATPのリン酸基を、デンプン代謝に関与する酵素の特定アミノ酸であるα-アミラーゼに転移させている。「ジベレリン受容体」を欠損するイネの変異体は、そのα-アミラーゼを合成できない。胚でジベレリンが合成されると、ジベレリンは種子の皮の糊粉層へ移動し、糊粉層でアミラーゼが合成される。合成されたアミラーゼは胚乳の貯蔵澱粉を分解し、胚の成長に使われる。糊粉層は、イネ科の植物の種子の皮の部分にある、肥厚した一次細胞壁を持つ特殊な分泌組織で、その細胞にアリューロン果粒(アリューロンとは、ギリシャ語で“粉”を意味する)と名付けられたタンパク質を貯蔵する液胞が詰まっていることに由来する。
その糊粉層細胞膜にあるタンパク質に「ジベレリン」が結合しているようだ。細胞膜に局在する「ジベレリン受容体」の多様性は、糊粉層細胞には、α-アミラーゼの誘導を導くCa2+非依存性の情報伝達経路と、その分泌を制御するCa2+依存性の情報伝達経路がある。 イネやトウモロコシ、エンドウやインゲンマメには、背丈が伸びない「矮性」と呼ばれる品種があり、それらの品種には「ジベレリン」を作る遺伝子に変異があり、これらの品種にも外から「ジベレルリン」を与えれば背丈が伸びることが知られた。
 ジベレルリンが作物の背丈を伸ばすのは、オーキシンと同じで、「伸長成長」、即ち「吸水成長」をひきおこすためと考えられている。ジベレルリンが、「SAUR遺伝子small-auxin-up
RNA(オーキシン早期応答遺伝子)」の発現を促進することから、オーキシンと同じように「酸成長」のメカニズムで、細胞壁をゆるめ、吸水を促進していると見られている。SAUR 遺伝子はダイズ・タバコ・トウモロコシなどの様々な植物に存在し、シロイヌナズナでは78個、イネでは58個のSAUR
遺伝子が同定されている。
ジベレルリンが作物の背丈を伸ばすのは、オーキシンと同じで、「伸長成長」、即ち「吸水成長」をひきおこすためと考えられている。ジベレルリンが、「SAUR遺伝子small-auxin-up
RNA(オーキシン早期応答遺伝子)」の発現を促進することから、オーキシンと同じように「酸成長」のメカニズムで、細胞壁をゆるめ、吸水を促進していると見られている。SAUR 遺伝子はダイズ・タバコ・トウモロコシなどの様々な植物に存在し、シロイヌナズナでは78個、イネでは58個のSAUR
遺伝子が同定されている。酸性に傾いた細胞壁では、「エクスパンシン」と言うタンパク質が活性化し、細胞壁は、繊維状のセルロースをはじめとする多糖類が、リグニン(主として中間層や細胞壁にセルロースやヘミセルロースと結合して植物体の強化に役立つ木質素。木化した植物体の主成分の一つ)と結合して強度を保持するが、「エクスパンシン」が活性化すると、これらの多糖類の結合が緩められ細胞壁が伸びやすくなる。
細胞伸長は若い細胞で起こる。植物細胞の成長は、吸水による容積拡大が大きな比重を占めるので、細胞が吸水して膨圧を高め、細胞壁を少しずつ押し広げる、そして細胞壁の合成が進み、軸方向に伸長成長する。オーキシンは細胞壁の伸展性を増加させる。
しかし細胞壁は細胞壁の合成の蓄積により、やがて伸展性を失い、細胞伸長は停止する。
オーキシンとジベレリンは、細胞伸長を促進する点では似ているが、そのどちらかでも作れなくなった変異体では矮小化する。これにより、オーキシンとジベレリンは、共同して細胞伸長を促進していことが分かる。
矮性のコムギやイネは、かつて人類の食糧難を救った歴史がある。1,940(昭和15)年代~60年代にかけて、穀物の大量増産を実現した「緑の革命」と呼ばれる農業革命である。当時、世界人口は急激な伸びを示し、1,900年の推定人口は、20億人であったが、1,950(昭和25)年に25億人、1,960(昭和35)年に30億人を超え、人口増加に見合う食料増産を達成することが世界的な課題となっていた。
1960年代に入ると、アメリカをはじめとする先進国の農業研究所では、トウモロコシ・小麦・イネなどの品種改良、特に収穫量の多い改良品種の開発が進められた。
最初は、化学肥料や農薬の投入による増産が試された。コムギやイネなどは、化学肥料を投与し過ぎて、茎の丈が不必要に高くなり、ついには倒伏が多発した。野生の植物は、光合成に必要な光を獲得するために、周囲の諸々の植物と丈の長さを競うことで優位に立とうとする。豊富な栄養は、最初に背丈を伸ばすことに使うのが、野生環境に生きる陸上植物の宿命である。
しかし、同じ田畑で同じ栽培作物が同時に育てられるなら、丈の高さにエネルギーと栄養を集中させることは、むしろ無駄である。投じられる肥料は、食料増産と種子の品質向上に向けられるべきで、矮小化のままの方が、かえって収穫量は安定し増加する。
国連は世界人口について、
「今後数十年で、耕作地を大規模に拡大せずに、食料の生産量を大幅に増産しなければならないが、直近の数十年て見ると、世界の小麦・コメ・トウモロコシなどの主要生産物の単収増加率は、1960年代や1970年代に比べて大きく鈍化している。問題は、単収の増加率が今後数十年における需要の伸びに追いついているか、ということである」
2,050年には世界の人口増に見合う食料生産が今のままではできず、世界的な飢餓もありうるとの見通しを立てている。
そこで、導入された主な高収量品種(High Yield Varieties: HYVs)として、メキシコシティー郊外でアメリカ合衆国の農学者ノーマン・E・ボーローグ博士らによって開発されたメキシコ小麦と呼ばれる短稈やコムギの多収穫品種群、フィリピン・マニラ郊外の国際稲研究所(IRRI)で開発されたイネ品種IR8などが挙げられる。これらの短稈品種は、植物体の背丈が低くなるが、穂の長さへの影響が少ない性質(半矮性)から選抜し導入したものである。半矮性の導入によって作物が倒伏しにくくなり、施肥に応じた収量の増加と気候条件に左右されにくい安定生産が実現した。なお、高収量品種を、近代品種と近年では言い換えられている。かつては、高収量品種と呼ばれたが、生産環境に関わりなく常に高収量を実現できるわけではないためである。
緑の革命に寄与した他の要因として、灌漑設備の整備・病害虫の防除技術の向上・農作業の機械化が挙げられる。『緑の革命』という用語は、1,968年に米国国際開発庁のWilliam S. Gaudによって造語されたものである。また、同一内容のことを「種子・肥料革命seed‐fertilizer revolution」と呼ぶ人々もいる。
穀物の多収穫品種を育成し、灌漑・肥料・農薬・農業機械などの技術革新で発展途上国の伝統的農法を脱し、急激な食糧増産がはかられ、米国のロックフェラーやフォード両財団の援助で、1,962年に、フィリピンのマニラ郊外にある国際稲作研究所(IRRI)では、1965年にいわゆるミラクル・ライスMiracle Rice(奇跡の米)とよばれるIR‐8が開発され、その翌年に同じくIR‐5を公表した。しかし、第一世代の近代品種は病虫害に弱く、環境不良地帯ではとりわけ収量性が低いという欠陥があった。
その近代品種Iは、急激に普及地域を拡大したが、病虫害による深刻な被害が続出した。それで、近代品種に病虫害抵抗性を付与する研究が1,970 年代初期に盛んになった。1,976年に開発された病虫害抵抗品種IR36にその成果があらわれた。その後の近代品種も、病虫害抵抗性を備えた品種であり、IR36を始めとする第二世代の近代品種IIの開発は、収量を増大させただけでなく、収量の安定によって農家の経営に大きく寄与した。
その詳細をたどれば、1,963年には、メキシコに、国際トウモロコシ・コムギ改良センターが開設され、メキシコ小麦とよばれる多収穫品種が開発されたのを契機に、画期的な多収穫短稈のイネとコムギの品種が育成され、インドやパキスタンなどの熱帯アジアやメキシコなどに急速に普及させたのが始まりのようだ。
緑の革命による農業の発展は、インドでは、小麦地帯である北西のパンジャーブとハリヤーナー両州や、その東のウッタル・プラデーシュ州西部を中心に1,967年から始まった。改良品種の作付け・灌漑設備の拡大・灌漑のためのディーゼルエンジンや電力の使用・化学肥料や農薬の投入・トラクターの使用などにより、米と小麦、特に後者の生産が上昇した。近代的農業は、かなり高度なレベルに発達し、これが、農民の経済状況の改善、ひいては社会的地位の向上につながり、現在、そこは世界有数の灌漑農業地帯となっている。雨期作の米と10~4月の乾季を作期とする小麦の相互乗入れによる米麦二毛作の拡大、主穀と綿花以外の重要農作物としてサトウキビの導入などが見られる。
台湾種とジャワ種を交配した新品種IR‐8は、収量を従来の品種の約2倍にすることができたが、収穫量は意外にも伸びず、味の点でもなお改良の余地があった。また、大量の肥料や農薬の散布、灌漑設備や農機具の充実など、近代的農業技術の導入を前提とするものであり、多額の資本投下を必要とした。そのため、新品種を導入できる農民や地域が限られ、特に、灌漑排水の不整備や肥料不足など、発展途上地域では、すぐに成果をあげられなかった。そのため農村内部の階層間、地域間の所得格差を拡大させた。また、化学肥料と農薬の大量投与による環境汚染や、新品種が短稈であるため雨期のデルタ地帯に適さないという欠点など、多くの批判を浴びた。
一方、アジアやアフリカの開発途上国における深刻な食糧不足の対策は喫緊だが、「第二の緑の革命」は、未だ大いなる課題のままの状況にあった。1980年代後半から、アフリカにおいて多収穫の新品種の導入が始まっている。また、新しいバイオテクノロジーによる多収穫品種の開発も行われている。その一方では、遺伝子組換えなどの技術に対する反発や、商品化された新品種を企業が独占するなどの問題点も指摘されている。
しかし、1,960年代に進められた稲・小麦などの多収穫品種の開発と、その導入によってもたらされた開発途上国における農業技術の革新、化学肥料の使用などによる穀物収穫量の飛躍的な増大により、国際連合食糧農業機関Food and Agriculture Organization(FAO)によると、アジアの穀類生産量は1,960年からの40年間で3倍に増え、1,969年にアジアの途上国で人口の42%を占めていた栄養失調比率は、2,000年に16%まで低下した。
米国の農学者ノーマン・E・ボーローグ博士は、インドや中国などでコムギなどの多収穫品種を中心とした新しい農業技術を開発し、穀物の大幅な増産を指導した。メキシコでも3倍もの生産量の向上を達成した。世界の食糧不足の改善に尽くし、「緑の革命」を実現したとして、1,970年にノーベル平和賞が与えられた。
ボーローグ博士は、稲塚権次郎が昭和10(1935)年に世に送り出した日本産のコムギの新品種「農林10号」を親に用いて背の低い丈夫なコムギを作った。これが奇跡のムギである。その後、コメやその他の穀物の「奇跡の品種」が続出し、世界の「緑の革命」の引き金となった。
日本のコメでは、近年、広域適応性を持つ関東以西に広く普及した日本晴や、食味優良という点で、北陸・東北地方を中心に広く栽培されているコシヒカリやササニシキなど、良食味品種の育成やブランド化が全国各地で盛んに行われ、産地間の競争が激化している。年々進む温暖化に備えるため、暑さに強いコメの育成に傾斜し、高温に弱いコシヒカリの後を狙うコメが、続々とデビューしている。
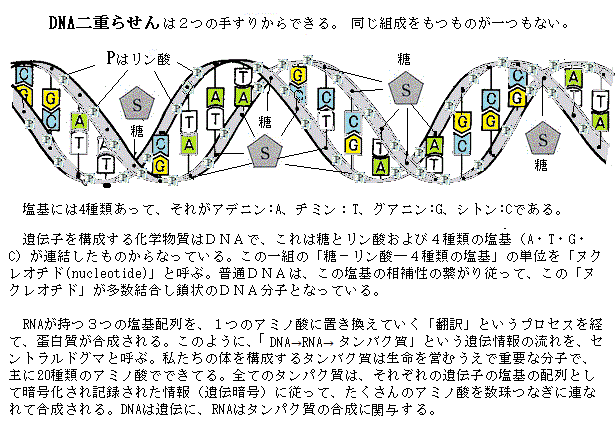 この時代の品種改良は、品種を掛け合わせる「交配」と言う古代からの手法で達成されてきた。それも当然で、遺伝子の本体はDNAであり、その「二重らせん」の立体構造が解明されたのは、ジェームズ・ワトソンとフランシス・クリックによる、1953(昭和28)年に発表された論文を待たねばならなかった。しかも、微生物のDNAの配列が、ようやく解読されたのが1,970年代の半ば過ぎ、植物の研究に遺伝子やDNAを解析する手法が用いられるようになったのは、20世紀の終わり頃であった。
この時代の品種改良は、品種を掛け合わせる「交配」と言う古代からの手法で達成されてきた。それも当然で、遺伝子の本体はDNAであり、その「二重らせん」の立体構造が解明されたのは、ジェームズ・ワトソンとフランシス・クリックによる、1953(昭和28)年に発表された論文を待たねばならなかった。しかも、微生物のDNAの配列が、ようやく解読されたのが1,970年代の半ば過ぎ、植物の研究に遺伝子やDNAを解析する手法が用いられるようになったのは、20世紀の終わり頃であった。ところが、「緑の革命」で開発された新品種の遺伝情報を、現代の研究手法で調べると、いずれも、作物の収量を増加させるジベレリンやブラシノステロイド(近年、植物ホルモンの一種として認められた、細胞伸長・細胞分裂・屈曲促進・木部分化などの関与する)などの重要な植物ホルモンにおける、生合成や信号伝達に関与する遺伝子が壊れた変異体であることが明らかになった。
遺伝学の基本法則を発見したメンデルが、実験の際に用いた矮性のエンドウも、後年に残るそのエンドウの遺伝子を調べると、ジベレリンの生合成に関与する酵素の遺伝子が壊れた変異体であることが分かった。
2,017年9月15日、国連が発表した世界の食料安全保障と栄養の現状に関する最新の年次報告書によると、10年以上、着実に減少を続けてきた世界の飢餓人口が、2,016年は再び増加に転じ、世界の人口の11%に当たる8億1500万人に達している。同時に、様々な形の栄養不良が世界中の何百万人もの人々の健康を脅かしている、とある。
人類が英知を傾けてきた「緑の革命」の成果を無にするほどの、人類の残忍さと独善が地球上に跋扈している。
目次へ