| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
| 目次 |
| 1)炭化水素鎖(hydrocarbon chain) |
| 2)異化 |
| 3)リン脂質 |
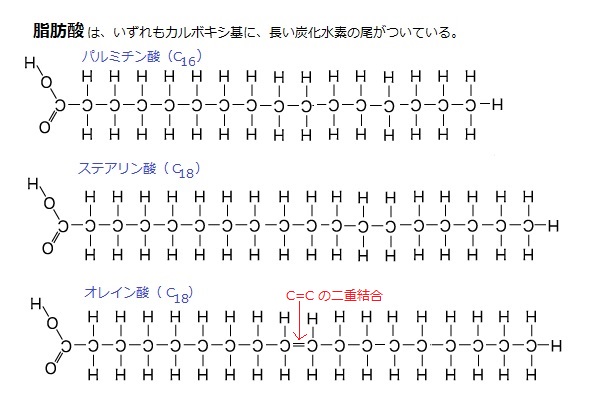
脂肪酸は、数百種類もある。炭化水素の尾部にC=Cの二重結合をもたないものを、飽和脂肪酸、二重結合があるものを不飽和脂肪酸と呼ぶ。
この二重結合は固定しているが、その両端で鎖状に折れ曲がることができる。しかも、C-Cの単結合の周りを自由に回転できる。
炭化水素鎖の飽和とは、その結合する炭素原子間に、二重結合がないため、連なる炭素が全て水素と結合しており、その鎖は最大限の水素原子をもつことを意味する。
そのため、鎖の繋がりが密に詰まることになり、折れ曲がりが生じない。それが炭化水素鎖を持つ重合体の性質を分けることになる。
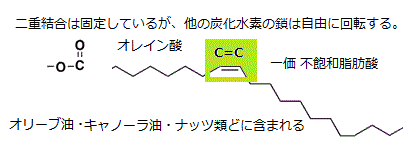
飽和脂肪酸に富むマーガリンは固いものとなる。二重結合が多い不飽和脂肪酸が豊富であれば、軟らかいマーガリンになる。脂肪酸の尾部は細胞膜にも並び、その詰まり具合が膜の流動性を決める。細胞には、様々な脂肪酸が豊富に含まれている。それぞれの炭化水素鎖の長さやC=Cの二重結合の有無と数、その位置の違いで、異なる性能を持つようになる。
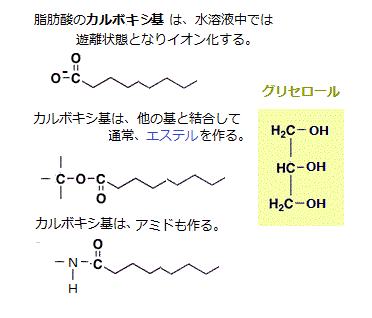
脂肪酸の多くは、食用油脂などに見られるように、いわゆる脂肪と呼ばれるトリアシルグリセロール(triacylglycerol)分子となって、細胞内で液滴(表面張力でまとまった脂肪滴)の状態で、多くの細胞質に蓄えられている。トリアシルグリセロール分子は、1個のグリセロール(glycerol)分子に3個の脂肪酸がエステル結合した化合物であることから名付けられた。ラテン語の「tri」は「数の3」である。「グリセロール」はグリセリン(glycerin)ともいい、ギリシア語のglykys(甘い)に由来する。1957年アルコールの化学名の語尾は「オール」とされ、グリセリンの正式化学名はグリセロールと国際機関で定められた。
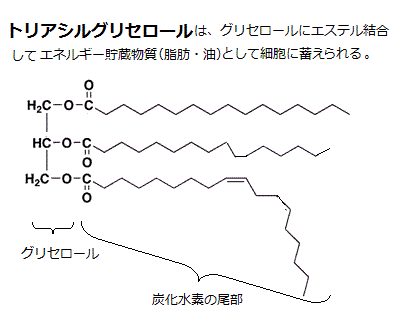
脂肪は、「三大栄養素」の一つとされ、食生活には極めて有用である。トリアシルグリセロールも食物から摂取できる。肝臓は炭水化物から脂肪酸を生成し、さらに脂肪酸とグリセロールからトリアシルグリセロールを合成する。ただし、飽和脂肪は必須ではない。流動性に欠けるため、血中のコレストロール濃度を上昇させ、動脈を塞ぐので、心臓発作や脳卒中のリスクを高める。
「中性脂肪」は、動物の脂肪組織に蓄えられる脂肪や、食品中の油脂・植物油(種子)などを構成する脂質の8から9割を占めるが、中でもトリアシルグリセロールが圧倒的に多く、特に動物の脂肪組織では95%を超えている。 構成している脂肪酸の種類によって、飽和脂肪酸が多ければ常温で個体(脂肪)であり、不飽和脂肪酸が多ければ常温でも液体(油)となる。
動物では皮下脂肪のことで、植物では種子に多く含まれ「油脂」ともいう。飽和脂肪は、ラードなどの肉の脂身やバター・クリームなど乳製品に見られ、不飽和脂肪酸はコーン油・オリーブ油のような植物油で代表されるように、植物油は室温でも液体である。
不飽和脂肪酸は、炭素原子の二重結合が一つあるモノエン脂肪酸と、二重結合が複数あるポリエン脂肪酸とがある。天然油脂として、動植物油中に含有されるグリセリドは、モノエン脂肪酸で、オリーブ油・つばき油などに多く含まれる。 リノール酸は、二重結合を2つ持っており、体内では作ることができず、食品からとらなければならない油の成分であるが、必須の脂肪酸である。血中のコレステロール値を下げ、動脈硬化の予防には有効とされている。欠乏すれば皮膚炎などを起こす。
エネルギーが、必要になると、脂肪酸がトリアシルグリセロール分子から離れて、炭素が2個ある単位体に分解される。この単位はグリコースの分解で生じるものと同じで、同じエネルギー獲得反応径路に入る。脂肪酸が分解すると、同じ重量のグルコースの約6倍のエネルギーを発生させる。そのため細胞にとって、極めて密度の高い保存食になる。
目次へ
 細胞は、生体として、化学的・物理的なグルコース(glucose;ブドウ糖)の分解などをコントロールし、日々たゆまず誕生し、成長し、分裂する活動をするために、常にその活動源となるエネルギーの支えが不可欠である。そのために、食物分子に含まれる化学総合エネルギーに頼る。
細胞は、生体として、化学的・物理的なグルコース(glucose;ブドウ糖)の分解などをコントロールし、日々たゆまず誕生し、成長し、分裂する活動をするために、常にその活動源となるエネルギーの支えが不可欠である。そのために、食物分子に含まれる化学総合エネルギーに頼る。食物分子は、細胞にとって不可欠な動力源であり、特に重要なのが「糖」である。 植物は、光合成によってCO2から糖を作り出す。動物は植物や、植物を食べて育った生物を食べて、糖や糖に化学変換される有機分子を摂取する。ところが、糖を分解してエネルギーを獲得する過程は、植物も動物も極めて似ている。
生物の細胞は、糖分子を分解し酸化して、二酸化炭素(CO2)と水(H2O)にするとき、糖に蓄えられた化学総合エネルギーから有用なエネルギーを取り出す。この過程を細胞呼吸(cell respiration)と呼ぶ。この反応で放出されるエネルギーは、ATPやNADHなどのような活性運搬体として、高エネルギー化学結合され蓄えられる。この結合は、加水分解されたとき、大量のエネルギーを放出する共有結合である。これらの運搬体は、生合成に必要な化学基や電子の供給源にもなる。
消化によってできた有機小分子は、細胞質に入り、そこで緩やかな酸化的分解がなされる。第一段階では、主に口や胃腸による細胞外で起こるが、細胞内のリソソームでも大型有機分子が消化される。 リソソームは、消化酵素を含んだ微細な袋で、不要になった細胞小器官やエンドサイトーシス(endocytosis;飲食作用)により細胞が取り込んだ巨大分子や粒子を分解する。
リソソームまでの径路は、物質の出所により異なる。細胞外の粒子は、食胞に取り込まれ、その食胞がリソソームと融合する。細胞外液と巨大分子は、小型のエンドサイトーシス小胞に取り込まれて、選択的透過性を持つエンドソームという一連の小区画をまず通過してからリソソームに送られる。エンドソームは、取り込んだ分子を選別し、一部を再利用するため細胞膜に送り返す。
膜に包まれた細胞小器官は、真核細胞内の至る所にあり、それぞれ特定の役割を果たす。細胞質内の細胞小器官の外側を満たす水溶液を「細胞質ゾル」と呼ぶ。細胞小器官であるリソソーム・小胞体・ゴルジ体などと、細胞外部の間では、絶えず物質の交換が行われている。小さなシャボン玉が大きなものから分かれて、離れた別のシャボン玉と合体するように、細胞小器官の袋から、くびれて離れた輸送小胞が、別の細胞小器官と融合して、交換を仲介する。
例えば、ゴジル体の細胞表面で、その細胞膜の一部が陥入して、くびり取られるように小胞となり、細胞膜との融合を促すシグナルが来るのを待つ。
食胞に摂取され、細胞膜に閉じ込められた細胞外の物質は、細胞膜の一部に徐々に取り囲まれ、そこが内側に陥入して首切られて、細胞内に運び入れられエンドサイトーシス小胞ができる。それには膜の成分も含まれているが、エンドソームに送られ、細胞膜に戻され再利用されるか、リソソームに送られ消化される。消化されて生じた代謝産物は、リソソームから直接細胞質に運び出され再利用される。これが「飲食作用」、即ち「エンドサイトーシス」である。これにより動物細胞は、巨大な粒子ばかりか、細胞さえ丸ごと飲み込む。この逆が「開口分泌」、則ち「エキソサイトーシス(exocytosis)」である。
細胞内の小胞は、細胞膜と融合し、その内容物を細胞外へ放出する。細胞は、細胞間の伝達に関わるシグナル伝達分子やホルモンの大部分を、このエキソサイトーシスから放出する。
細胞内に取り込まれた細胞外の粒子や分子の多くは、最後にリソソームに送られる。膜で囲まれた袋状のリソソームには、加水分解酵素が含まれており、細胞外から取り込まれた物質や老化した細胞小器官の細胞内消化を、調節しながら進めていく。リソソームには、タンパク質・核酸・オリゴ糖・脂質などを分解する40種類の加水分解酵素があり、これらの酵素は、リソソーム内部が酸性環境で活性化し、濃度がpH5で最大活性を示す。細胞質のpHは、7.2程度という。
リボソームの膜には、輸送体があって、巨大分子を分解した最終産物であるアミノ酸・糖・ヌクレオチドなどを細胞質に搬出する。細胞は、これらを外部に排出したり、細胞内で再利用したりする。 なお、ほぼ全ての真核生物の細胞質に見られる、小型の細胞小器官であるベルオキシソーム(peroxisome)にも、脂質の分解や有毒物質の破壊など様々な酸化反応に関わる酵素が含まれている。
グルコース以外の糖も、解糖系の中間体のどれかに変化した後に利用される。解糖は細胞質で起こり、6炭素のグルコースが、3炭素のピルビン酸2分子に分割され、ATPとNADHという2種類の活性運搬体を作る。 ピルビン酸分子中の電子の方が、グルコース分子中の電子より、エネルギーが低いため、ピルビン酸ができるまで、化学的再配置が次々と起こり、エネルギーが放出される。
グルコース ➡ 2ピルビン酸 + 2ATP + 2NADH
解糖で分割されるグルコース1分子あたり、ATP2分子が消費されるが、解糖後、4分子のATPが生成され、加えてNADH2分子も得られる。
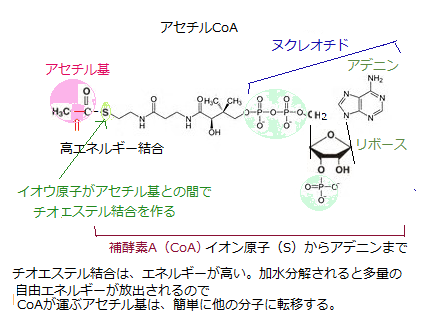 好気性生物では、このNADH分子からミトコンドリア内膜にある電子伝達系に電子が渡される。この電子伝達により、電子は高エネルギー状態から低エネルギー状態に落ちエネルギーを放出する。電子伝達系をたどった電子は、最後にO2に達し水を作る。電子を手放したNADHはNAD+に戻り、再び解糖に使われる。 その後、真核細胞の好気性代謝では、解糖で生じたピルビン酸は、細胞質からミトコンドリアの大きな内部区画・マトリックに移り、そこで巨大なピルビン酸脱水素酵素複合体によって、直ぐに脱炭酸され、廃棄物としてのCO2と、活性運搬体分子の1つであるアセチルCoAとNADHが同時に生成される。なお、ピルビン酸脱水素酵素複合体は、リボソームより大きい。
好気性生物では、このNADH分子からミトコンドリア内膜にある電子伝達系に電子が渡される。この電子伝達により、電子は高エネルギー状態から低エネルギー状態に落ちエネルギーを放出する。電子伝達系をたどった電子は、最後にO2に達し水を作る。電子を手放したNADHはNAD+に戻り、再び解糖に使われる。 その後、真核細胞の好気性代謝では、解糖で生じたピルビン酸は、細胞質からミトコンドリアの大きな内部区画・マトリックに移り、そこで巨大なピルビン酸脱水素酵素複合体によって、直ぐに脱炭酸され、廃棄物としてのCO2と、活性運搬体分子の1つであるアセチルCoAとNADHが同時に生成される。なお、ピルビン酸脱水素酵素複合体は、リボソームより大きい。 脂肪由来の脂肪酸も、主要なエネルギー源となる。段階的な酸化的分解を経て、ミトコンドリア・マトリックスで、アセチル基と結合する大量のアセチルCoAに変換される。脂肪は水に溶けず、脂肪酸誘導体・トリアシルグリセロール(triacylglycerol)となって、専門の脂肪細胞の細胞質に大きな脂肪滴を作る。脂肪細胞は、通常の細胞の数倍~10倍もの直径120 μm(1 μm = 1/1000 mm) に達する巨大な細胞であり、その内部は 1 個の大きな脂肪滴でほとんど埋め尽くされている。空腹時や運動時など、エネルギーが不足した際には、脂肪細胞に蓄えられた脂肪滴が加水分解され、脂肪酸とグリセロールのエステル結合を切断し、いずれも血中に放出される。脂肪のもつエネルギーの大半は、脂肪酸に含まれている。
 好気性生物では、このNADH分子からミトコンドリア内膜にある電子伝達系に電子が渡される。この電子伝達により、電子は高エネルギー状態から低エネルギー状態に落ちエネルギーを放出する。
好気性生物では、このNADH分子からミトコンドリア内膜にある電子伝達系に電子が渡される。この電子伝達により、電子は高エネルギー状態から低エネルギー状態に落ちエネルギーを放出する。電子伝達系をたどった電子は、最後にO2に達し水を作る。電子を手放したNADHはNAD+に戻り、再び解糖に使われる。 その後、真核細胞の好気性代謝では、解糖で生じたピルビン酸は、細胞質からミトコンドリアの大きな内部区画・マトリックに移り、そこで巨大なピルビン酸脱水素酵素複合体によって、直ちに脱炭酸され、廃棄物としてのCO2と、活性運搬体分子の1つであるアセチルCoAとNADHが同時に生成される。なお、ピルビン酸脱水素酵素複合体は、リボソームより大きい。
脂肪由来の脂肪酸も、主要なエネルギー源となる。段階的な酸化的分解を経て、ミトコンドリア・マトリックスで、アセチル基と結合する大量のアセチルCoAに変換される。その後、後述するクエン酸回路で主導的な役割を果たす。
脂肪は水に溶けず、脂肪酸誘導体・トリアシルグリセロール(triacylglycerol)となって、専門の脂肪細胞の細胞質に大きな脂肪滴を作る。脂肪細胞は、通常の細胞の数倍~10倍もの直径120 μm(1 μm = 1/1000 mm) に達する巨大な細胞であり、その内部は 1 個の大きな脂肪滴でほとんど埋め尽くされている。
空腹時や運動時など、エネルギーが不足した際には、脂肪細胞に蓄えられた脂肪滴が加水分解され、脂肪酸とグリセロールのエステル結合を切断し、いずれも血中に放出される。脂肪のもつエネルギーの大半は、脂肪酸に含まれている。
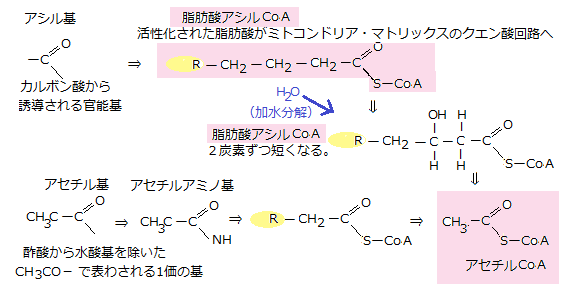
その脂肪酸からエネルギーを得る必要が生じると、リパーゼ(lipase)という酵素が、トリアシルグリセロールのグリセロールと脂肪酸の間のエステル結合を切断する。放出されたトリアシルグリセロールの炭化水素鎖の尾部・脂肪酸は、まずATPに頼る反応で、活性運搬体(アシル補酵素A)と結合して脂肪酸アシルCoAとなる。
細胞は、転移を起こしやすい、エネルギーの高い結合体である、活性運搬体を活用して基を運ぶ。活性運搬体の補酵素A(CoA)も、転移しやすいアシル基やアセチル基と結合する。それにより真核細胞のミトコンドリア・マトリックスへ運ばれ、活性化された脂肪酸アシルCoAは、4種類の酵素からなるクエン酸回路(citric acid cycle)で酸化される。
citric(sɪ́trɪk)とは、「クエン酸の」意である。回路を1周するたびに、脂肪酸アシルCoAは、2炭素ずつ短くなり、アセチルCoAが1分子生じ、それに伴い2種類の活性運搬体、NADHと別の高エネルギー電子輸送体であるFADH2、それぞれが1分子ずつ生じる。
殆どの細胞は、炭素化合物酸化の全工程のほぼ2/3を担うクエン酸回路で、アセチルCoAのアセチル基がCO2とH2Oに酸化される。その主な最終生成物は、CO2とNADHに蓄えられた高エネルギー電子で、CO2は廃棄物として排出される。NADHの高エネルギー電子は、ミトコンドリア内膜にある電子伝達系に渡され、最後はO2と結合してH2Oとなる。 クエン酸回路自体がO2を使うわけではないが、回路の進行にはO2が必要である。電子伝達系がO2を最終受容体としてNADHから電子を取り除き、サイクルを回し続けるのに必要なNAD+を再生するからである。
生命体は35億年前の地球上に、明らかに存在していた。しかし地球大気がO2を豊富に含むようになったのは、10億年前から20億年前の間と見られている。クエン酸回路のエネルギー生成反応は、生命体の誕生に関わっていなかったことになる。
クエン酸回路は、その触媒作用でアセチルCoAのアセチル基の炭素原子を、完全に酸化してCO2にする。もっとも、アセチル基はそのまま酸化されるわけでなく、4炭素分子であるオキサロ酢酸に転移して、6炭素のトリカルボン酸とクエン酸を作る。
 アセチルCoAなどの活性運搬体にある転移可能な基は、図に示すように構造分子のごく一部に過ぎない。残りの方が多い有機性の大部分は、酵素が運搬体分子を見分けられるよう、「marker」としての役割を果たす。その「marker」は、アセチルCoAのみならず運搬体分子の多くに、ヌクレオチドが含まれている。
細胞進化の初期段階、生命体の触媒は、主にRNAか、またはそれに近い分子であって、タンパク質は、その後の進化の過程で登場したようだ。
アセチルCoAなどの活性運搬体にある転移可能な基は、図に示すように構造分子のごく一部に過ぎない。残りの方が多い有機性の大部分は、酵素が運搬体分子を見分けられるよう、「marker」としての役割を果たす。その「marker」は、アセチルCoAのみならず運搬体分子の多くに、ヌクレオチドが含まれている。
細胞進化の初期段階、生命体の触媒は、主にRNAか、またはそれに近い分子であって、タンパク質は、その後の進化の過程で登場したようだ。アセチルCoAなどの運搬体分子は、原始におけるRNAの起源にまで遡れる年代に、RNA主体の酵素・リボソームを創始するに至ったと考えられる。 リボソームは、細胞内では、最も大型で複雑な構造体の一つで、2/3がRNA 、1/3がタンパク質からなり、しかも、その全体構造を作り、タンパク質を合成しているのがrRNAである。現代、触媒活性を担う酵素は、殆どがタンパク質であるが、タンパク質が生成されていない生命誕生から初期にかけて、細胞内では、触媒活性を持つ原始のRNAが、細胞内外の働きを調節していたと見られる。
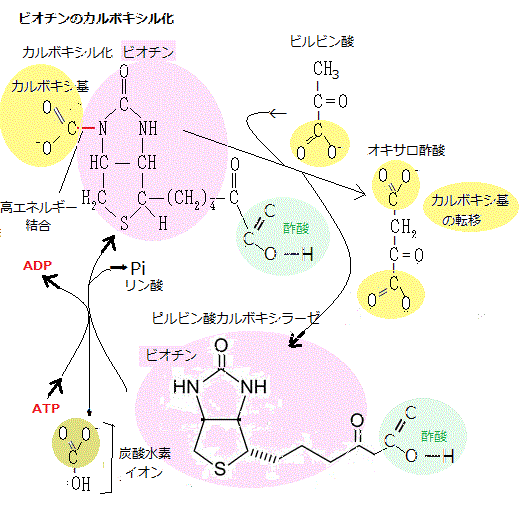 ビオチンは、水溶性ビタミンの一種で、ビタミンB7とも呼ばれるが、単にビオチンと呼ばれることも多い。生体内において、カルボキシル基転移酵素(carboxylase)の補酵素として働く役割から補酵素Rと呼ばれることもある。
ビオチンは、水溶性ビタミンの一種で、ビタミンB7とも呼ばれるが、単にビオチンと呼ばれることも多い。生体内において、カルボキシル基転移酵素(carboxylase)の補酵素として働く役割から補酵素Rと呼ばれることもある。ビルビン酸カルボキシラーゼなど多くの酵素の構成単位となっている。そのビオチンのような活性運搬体は、通常、ATPの加水分解と共役する反応で作られる。運ばれた基が、生合成に関わるときのエネルギーも、元々はATPを作り出す異化反応から生成される。特に、巨大分子の核酸・タンパク質・多糖などの合成でも似た反応をたどる。
ビルビン酸カルボキシラーゼなどの多くの酵素に存在するビチオンは、カルボキシ化されると、カルボキシ基を他の分子に転移させる。図では、カルボキシ基が酢酸に転移され、クエン酸回路に必要なオキサロ酢酸が生成されている。
他の分子にビオチオンを使ってカルボキシ基を移す酵素もある。カルボキシル化ビオチンの合成には、ATPの加水分解のエネルギーを使う。これは多くの活性運搬体に共通する特徴である。
真核細胞の好気性代謝では、解糖で生じたピルビン酸は、ミトコンドリア・マトリックスに取り入れられる。そこで3酵素からなる巨大なピルビン酸脱水素酵素複合体によって、直ぐに脱炭酸される。その結果、NADH・アセチルCoAと、廃棄物としてのCO2が生成される。
解糖で生じたピルビン酸と同様に、脂肪由来の脂肪酸も、ミトコンドリア・マトリックスで、アセチルCoAに変わる。脂肪酸は、CoAとの共有結合でまず活性化され、反応回路を1周するたびに、カルボキシ末端から炭素を2個ずつ切り取られ、やがて完全に分解される。1周ごとに1分子のアセチルCoAが生じ、それに伴い2種類の活性運搬体NADHと別の高エネルギー電子輸送体であるFADHができる。 ピルビン酸と脂肪のみならず、アミノ酸の一部も細胞質からミトコンドリア・マトリックスに入り、アセチルCoAか、クエン酸回路の中間体のどれかに変化する。真核生物のエネルギーを獲得する異化過程では、糖・脂肪・タンパク質のどれから始まっても、ミトコンドリアに集まる。
異化はアセチルCoAの生成で終わるわけではなく、植物分子のアセチルCoAへの変換では、分子に蓄えられたエネルギーのごく一部が取り出され、ATP・NADH・FADH2になるだけで、大部分は、まだアセチルCoAに留まっている。細胞呼吸の次の段階は、クエン酸回路で、これによりミトコンドリア・マトリックスで、アセチルCoAのアセチル基がCO2とH2Oに酸化される。
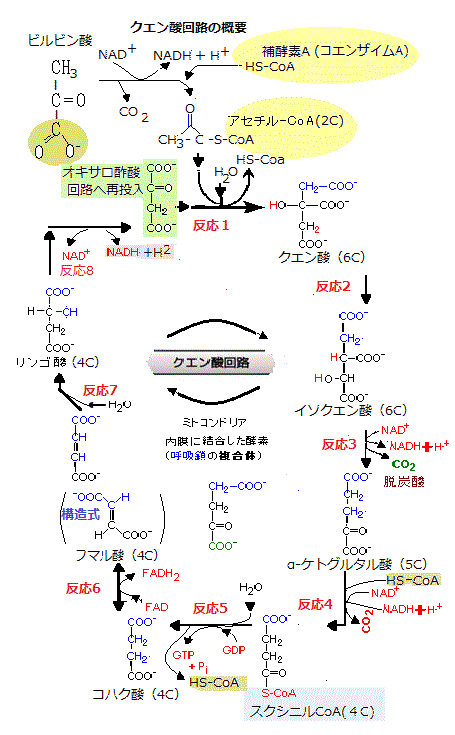
クエン酸分子は、徐々に酸化され、解糖の際と同様に、その酸化エネルギーによって活性運搬体が作られる。クエン酸回路は、食物由来のピルビン酸からできたアセチルCoAのアセチル基の炭素原子を、その触媒作用で、完全に酸化してCO2にする。ただ、アセチル基は、その状態のまま酸化されるのではなく、全体として8個の反応を通過する回路がある。
その回路を1周すると、NADHが3分子、GTPが1分子、FADH2が1分子生じ、他にCO2が2分子放出され、4炭素分子であるオキサロ酢酸に転移し、6炭素のクエン酸(3つのカルボキシル基をもつ化合物という意味でトルカルボン酸)を作る。これが回路の名前の由来であり、これによりクエン酸回路という。
殆どの細胞では、クエン酸回路が、炭素化合物を酸化させる全工程のほぼ2/3を担う。その主な最終生成物は、CO2とNADH(C21H29N7O14P2)として蓄えられる高エネルギー電子で、そのCO2は廃棄物として排出される。
NADHの高エネルギー電子は、ミトコンドリア内膜にある電子伝達系に渡され、最終的にはO2と結合してH2Oになる。 左図でも分かるように、ミトコンドニア・マトリックスで起こるクエン酸回路自体がO2を使うことはないが、その回路進行の過程でO2は不可欠な存在となる。電子伝達系がO2を最終受容体にしてNADHから電子を取り除いて、NAD+(C21H27N7O14P2)を再生し、そのサイクルを回し続けるからである。
地球の生命体は、35億年前には、既に存在していたようだ。しかし、大気中がO2で満たされるようになるのは、10億年前から20億年前と言う。生命体が、クエン酸回路から得られるエネルギー生成の条件が、漸くその間に整ったと言える。
クエン酸分子は、徐々に酸化され、解糖の際と同様に、その酸化エネルギーによって活性運搬体が作られる。このクエン酸回路の8個の反応により、最後にはオキサロ酢酸が再生され、再び次のサイクルに投入される。
次のサイクルに入ると、1周ごとに3分子のNADHが生じるほかにCO2が2分子放出され、FADH2(還元型フラビンアデニンジヌクレオチド:C27H33N9O15P2)1分子からFADが、リボヌクレオシド三リン酸のGTP(グアノシン三リン酸)1分子からGDPが作られる。
GTPはATPによく似た分子で、回路が1周するたびに末端のリン酸基をADPに渡しATPを作る。FADH2はNADHと同様、高エネルギー電子と水素の運搬体である。NADHやFADH2の転移しやすい高エネルギー電子に蓄えられたエネルギーは、酸化的リン酸化によるATP合成に使われる。酸化的リン酸化はミトコンドリア内膜で起こり、食物の酸化的異化のうちで大気中のO2を直接必要とする唯一の過程である。 クエン酸回路については、反応を進めるのに必要な大気中のO2が、廃棄物として放出されるCO2と結合していると誤解されがちだが、実際は、クエン酸回路に入ったアセチル基をCO2にするのに必要な酸素原子は、大気中のO2ではなくH2Oから供給される。1周ごとに3分子の水が分解され、最終的にはその酸素原子の一部がCO2の生成に使われる。大気中のO2は、電子伝達系で還元されH2Oになる、吐き出されるCO2に直接取り込まれることはない。
解糖系やクエン酸回路の異化反応では、細胞のエネルギーだけでなく、様々な有機分子を作るための構成材料も作られる。解糖やクエン酸回路で作られる中間体の多くは、同化径路(anabolic pathways; anabolic; `ænəbάlɪk; 同化作用の)に組み入れられ、一連の酵素触媒反応によってアミノ酸・ヌクレオチド・脂質やその他の細胞が必要とする有機小分子に変わる。例えば、クエン酸回路で作られるオキサロ酢酸とα―ケトグルタミン酸は、ミトコンドリア・マトリックスから細胞に戻され、それぞれアスパラギン酸をはじめ多くの必須分子生成の前駆物質となる。この過程は複雑で、主要な異化反応から枝分かれした、いくつもの径路に繋がっている。
転移したアセチル基は、この反応でCO2に酸化され、その際大量のNADHが生じる。最後に、NADHの高エネルギー電子は、ミトコンドリア内膜にある酵素群、電子伝達系をたどるのだが、ここで電子の移動に伴って放出されるエネルギーが酸化的リン酸化、すなわちATPの生成と分子酸素(O2ガス)の消費という過程を引き起こす。
酸化によって放出されるエネルギーで、細胞の殆どのATPを作るのが、この最終段階である。 糖や脂肪の分解で取り出されるエネルギーが、ATPという細胞内で使うのに便利な化学エネルギーの塊に変換される。全体としてグルコースや脂肪酸のH2OとCO2への分解で、理論的に得られるエネルギーの約半分が捕捉され、 エネルギー的に起こりにくい ADP + P1 ➡ ATP の反応を進めるのに使われる。
これに対して、自動車エンジンなどの燃焼エンジンでは、燃料の利用可能エネルギーのうちせいぜい20%しか有効に使われていない。どちらの場合も、残りのエネルギーは、熱として放出され、動物の場合は体を温めている。 細胞1個には、いつも109個(1億個)のATPが存在し、多くの細胞では、1~2分ごとにすべてのATPが消費され入れ替わる。ヒトは、安静にしていても、24時間で自分の体重分のATP分子を加水分解する。
目次へ
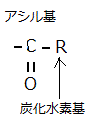
脂肪酸やトリアシルグリセロール(triacylglycerol)など脂肪酸誘導体は、脂質(lipid)の仲間である。トリアシルグリセロールの名前の由来は、脂肪酸の尾部3本が、グリセロールに、エステル結合した置換基がアシル基であるため、トリ・アシル・グリセロール=3分子・脂肪酸・グリセロールという意味に分解できるからである。殆どの脂肪酸は、カルボキシ基(―COOH)を介して、他の分子と共有結合している。カルボキシ基は、細胞などの水溶液中では、イオン化し、カルボン酸(―COO)として振舞う。アシル基(―C=O)は、カルボン酸から誘導された官能基である。
トリアシルグリセロール化合物は、脂肪の保存形態として存在し、動物組織の脂肪滴(飽和脂肪酸)や植物油(主に不飽和脂肪酸)の主成分となる。 脂質は、タンパク質・糖質とともに生体中に存在する物質で、水に溶けにくいが、脂肪やベンゼンのような有機溶媒に溶けやすい、長鎖脂肪酸やその誘導体・類似体で、生体を構成する一群の生体分子と、定義されている。
脂質には、脂肪酸のように長い炭化水素鎖となって繋がるものと、ステロイドのように環状構造を形成するものとがある。
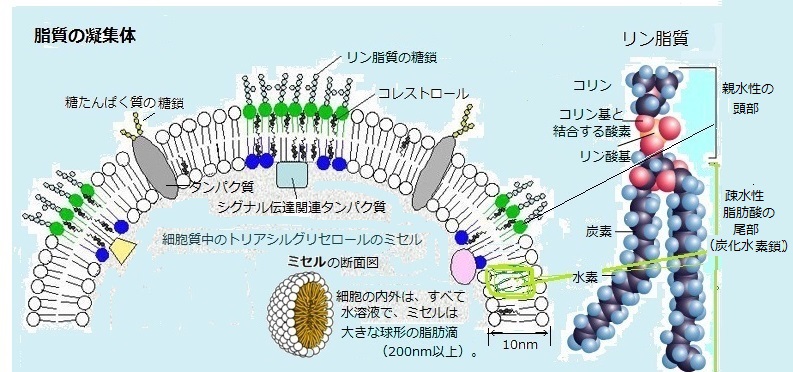
 lipid bilayer)を作ることである。「bilayer(báilèiər)」とは、二重層・二分子膜の意で、この薄い膜を作る脂質は、1個の分子が2つの全く異なる性質を備えている。主にリン脂質からなり、そのリン脂質が集合して細胞膜を作り細胞を包むが、特に真核細胞では、細胞内膜が細胞質ゾルと様々な小器官を取り囲んでいる。動物細胞内では、核とミトコンドリアだけが、2枚の膜で包まれている。
殆どのリン脂質は、脂肪酸とグリセロールが主体の小分子で、トリアシルグリセロールに似ている。
lipid bilayer)を作ることである。「bilayer(báilèiər)」とは、二重層・二分子膜の意で、この薄い膜を作る脂質は、1個の分子が2つの全く異なる性質を備えている。主にリン脂質からなり、そのリン脂質が集合して細胞膜を作り細胞を包むが、特に真核細胞では、細胞内膜が細胞質ゾルと様々な小器官を取り囲んでいる。動物細胞内では、核とミトコンドリアだけが、2枚の膜で包まれている。
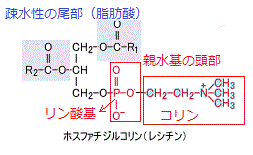
殆どのリン脂質は、脂肪酸とグリセロールが主体の小分子で、トリアシルグリセロールに似ている。グリセロール分子は、リン酸基に結合した親水性の頭部と2本の炭化水素鎖からなる疎水性の尾部を繋ぐ。グリセロールのーOH基が、脂肪酸の炭化水素鎖の一端にあるーCOOH基に結合しているのは、3個ではなく2個である。その不飽和脂肪酸にある炭化水素鎖の一方にある折れ曲がりは、C=Cの二重結合があるところで生じる。グリセロールの残りの1個のーOH基は、親水性のリン酸基に結合している。
ホスファチジルコリン(phosphatidylcholine)では、そのリン酸基に、さらにコリンのような親水性の小分子が結合し、親水性の頭部をなしている。ホスファチジルコリンは、細胞の膜で最もよく見られるリン脂質で、脳の神経細胞膜の主成分でもある。
リン脂質は、炭化水素鎖が構成する2本の脂肪酸からなる疎水性の尾部と、リン酸基を含む親水性の頭部を持ち、両親媒性が強い。この独特な両親媒性の構成と形により、リン脂質は、ほぼ全体が疎水性となるトリアシルグリセロールとは、物理的・化学的に、その特性が異なる。
細胞膜には、リン脂質に加え糖脂質など、他の脂質が様々な量で含まれる。 糖脂質は、リン酸基の代わりに1個以上の糖を持つ。リン酸質と同様に、2本の長い炭化水素鎖からなる疎水性部分と、1個若しくは複数の糖が含まれる極性部分からなる。 リン脂質が水中で膜を作るのは、両親媒性という性質による。リン脂質が水面に広がって単分子層を作るとき、疎水性の尾部を構成する分子や疎水基をもつ分子などは、水溶液中にあると、水との接触を避けるために水と接する面積を減らそうと、互いに会合する疎水力が働く。
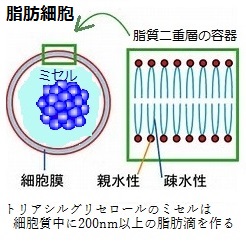
水中では、このような分子層が2枚、尾部同士が合わさってくっつき、リン脂質によるサンドイッチができる。これが脂質二重層で、リン脂質と糖脂質は、細胞の膜を構成する容器を形成する。 トリアシルグリセロールなどの脂肪酸分子の凝集体にも、親水性の頭部と疎水性の尾部があり、脂質二重層により形成される細胞の膜内では、脂肪酸の誘導体は、疎水力によって集合し、もっと大きな凝集体を作る。分子量(分子の相対的な質量)が大きな分子ほど分子間力が大きくなる。その分子間力により多数の分子の小塊を作る、これをミセル(micelle)と呼ぶ。 脂肪酸の分子内に親水性部の頭部と疎水性の尾部を合わせ持つ両親媒性物質であれば、真核生物の細胞の内外は、すべてが水性環境であるため、脂肪酸の誘導体は疎水力によって中心に集合し、親水性の頭部は、その外円を構成する。細胞の溶液中では、数十から百数十分子が集まって、最も大きな凝集体となる。脂肪酸は、グリセロールにエステル結合して、「トリアシルグリセロール」となり、エネルギー貯蔵物質である脂肪や油として細胞に蓄えられる。トリアシルグリセロールでは、大きなといっても、細胞質の中で200nmか、それ以上の脂肪滴を作っている。