| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
| 目次 |
| 1)陸上植物の誕生 |
| 2)有性生殖の進化 |
| 3)動物との共生 |
潮が引いた後の海岸の潮溜まりの岩の割れ目や窪みには海藻がくっついている。海で誕生した植物は、こうした上陸の手掛かりをつかみ、潮が引いても岩に固着し、環境が全く違う海から陸上へ進出する足場とした。
水辺にはゆらゆらとした糸状の藻類が繁茂している。水中には浮遊する小さな藻類が繁殖している。岩の表面にも、無数の藻類が付着している。
このような藻類たちが水中では主役である。その中に約 4.8 億年昔に水中から乾燥した陸地へ進出し、陸上植物へと進化した藻類がいた。これまでの研究の結果では、シャジクモ藻類(車軸藻類)のシャジクモ類とコレオケーテ類が陸上植物に最も近縁だと考えられている。
藻類は、水辺での生活から徐々に周りの環境に適応しつつ、大きな進化を遂げてきた。現在、地球上には、熱帯から極地まで、350万種もの多様な植物が生育している。それらの一個体ごとに、想像を絶した長い歴史と進化を経て、今日のたたずまいを作り上げてきた。
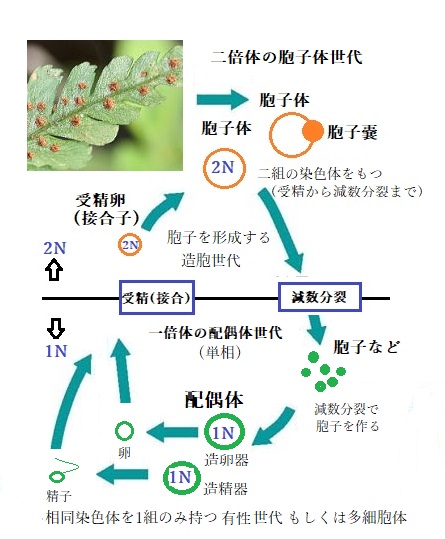
陸上植物はすべて胞子体と呼ばれる 両親に由来する 2 組の染色体を持つ2 倍体世代と、配偶体と呼ばれる1 組の染色体を持つ1倍体世代からなる世代交代を行っている。2倍体の胞子体から減数分裂をして生じた胞子が発芽して1 倍体の配偶体ができる。陸上植物の胞子体と配偶体はどちらも多細胞である。 コケ植物では胞子体より配偶体の方が大きく形態も複雑であるが、被子植物に近づくに従って大きさが逆転して胞子体の方が大きくなり形態も複雑になる。
一方、シャジクモ藻類では配偶体はあるが胞子体はない。つまり、配偶体(1倍体)が卵と精子を作り接合子(2倍体)ができるが、発芽すると体細胞分裂をせずに、すぐに減数分裂をして再び配偶体になり、胞子体を作らない。
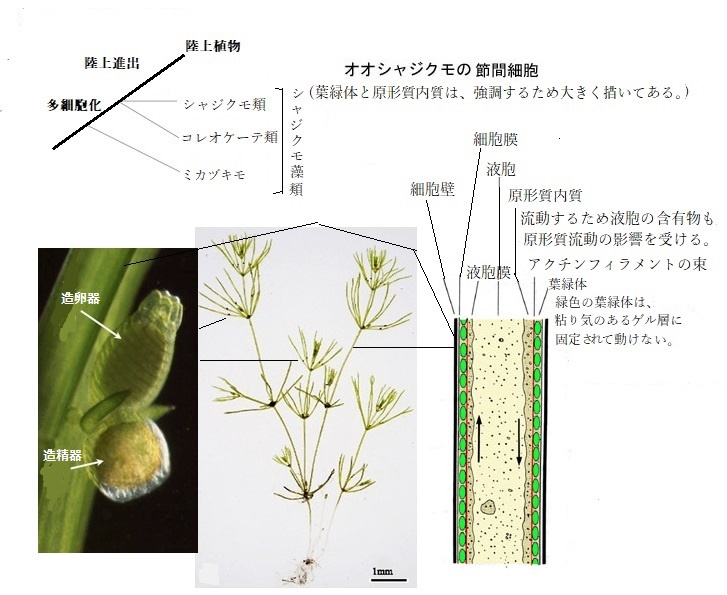
特徴的な多細胞体を形成するシャジクモ類は、高さ1 m以上になるものもいる。主軸とそこから輪生する枝、および仮根(かこん;真の根ではないが、根のように植物体を地上に固着し、水分の吸収を行う細長い細胞)からなる。主軸は節と節間の繰り返し構造からなり、節に枝を輪生させる。節は複数の節部細胞からなる柔組織であり、節間は1個の巨大な、ときに長さ15 cmに達する節間細胞からなる。節間細胞は顕著な原形質流動を示す。
軸の先端には1個の分裂細胞があり、これにより頂端成長する。原形質流動の影響により、液胞の含有物も流動する。枝の節に造精器と生卵器をつけ、卵生殖を行う。生殖器は多細胞性で、藻類としては極めて複雑な構造をしている。造精器はカロテノイド(γ-カロテン)を蓄積し、褐色を帯びる。受精卵はスポロポレニンを含む壁で覆われて休眠性の卵胞子となり、発芽時に減数分裂する。淡水域に生育し、透明度の高い湖沼では種子植物の水草よりも深い部分に密生し、車軸藻帯を形成していることがある。しかし、絶滅危惧種が多い。
上陸に成功した最初の植物は、緑藻類のシャジクモ類に似ていた。それでも体の半分は、水中で生活できる環境に適合した。潮が引いてもしっかりと岩に固着する潮だまりで、何物かに固着することから上陸が始まった。
大気は、二酸化炭素の濃度が高く、地球は高温多湿であった。この環境が水中植物の進出を乾燥から守り上陸を可能にした。実は、植物の上陸は、原始地球の環境下でのみ可能であった。その後は、再上陸の可能性は閉ざされた。現在の花咲く植物の多くは、この上陸に成功した植物の末裔である。
動物が誕生するのは、植物による食餌や酸素の供給などによって、生存可能な環境が整えられてからだ。動物と植物の出合いによって、自然界の至る所で、見事に共進化が成果をあげ、やがて、人類の登場を、約450万年前に促したと言える。
目次へ
 植物も動物も、初めは個体の一部を分裂させて、新しい個体を作る無性生殖で増殖した。やがて、雄と雌の合体という有性生殖という、複雑な繁殖方法を進化さていった。それも、最初は海の中での進化で始まった。
植物も動物も、初めは個体の一部を分裂させて、新しい個体を作る無性生殖で増殖した。やがて、雄と雌の合体という有性生殖という、複雑な繁殖方法を進化さていった。それも、最初は海の中での進化で始まった。別個体同士による細胞合体である有性生殖は、遺伝子の交換という生物の環境適応や進化のためには極めて重要であった。
有性生殖も、初期においては、雌雄差は殆どない。それが鞭毛を備えて動ける精子と、母体に寄生したまま卵細胞に分化したため、その雌雄の両性が、はっきりと分かれるようになる。
植物では、機能が異なる2つの多細胞体ができた。精子と卵子を作る配偶体と、配偶体のもとになる胞子を作る胞子体とにである。それぞれは、独立する方向へ進化し、配偶体と胞子体という2つの世代が循環する「世代交代」を、海の中で確立した。
植物は、海の中で進化し、いろいろな器官を整えたが、その中には陸上で生活する上で障害となるものもあった。特に精子は、水中を泳ぐには都合がよい構造であるが、陸上植物では、水がなければ精子は泳げないので受精ができない。
そこで、水が殆どないところでも有性生殖を可能にする進化がなされた。この陸上という新たな環境条件に適応するためには、遺伝子の交換を伴う有性生殖が重要な意味を持つ。この改造に成功した植物群が、種子植物であった。やがて種子のみならず、花を持つ植物が、有性生殖に関わる構造自体を改善し、海辺から内陸へ進出し、広大であるが不毛な大地を、生命力あふれる豊かな緑野へと改造し続けていった。
東京の小石川植物園のほぼ中央にある大イチョウ(雌株)を研究材料として、明治29(1896)年に、理学部植物学教室の助手であった平瀬作五郎が、種子植物にも精子が存在することを発見した。
イチョウには、雌株と雄株がある。5月になると、風が花粉を雌株の生殖器官である胚珠の先端にできた水滴に運ぶ。やがて水滴とともに花粉は、雌の胚珠に宿り、そこで配偶子を形成して精子を作る。そして、全ての環境条件が整う、9月を待って受精する。
平瀬作五郎は、寄生虫かと思って当時助教授だった池野成一郎に見せたが、池野は一目見るなり「精子だ」と直感したという。その後明治29年9月9日に「花粉管端より躍然精虫の遊動して活発に転々突進する状況を目撃」し、10月には、「いてふノ精虫に就テ」という論文を発表している。これが世界で初めての裸子植物における精子の発見となり、これは日本の初期の生物学者による、世界的な生物学史上の偉業と評価されている。
平瀬作五郎のイチョウ精子発見の記事は、明治29年10月20日に発行された植物学雑誌に、「運動する精子(精虫)を形成するというものであるが、前回は花粉管の中でしか見ることができなかったが、本年9月9日には精子が泳ぎだすことを見ることができ、精子であることを確証できた」とある。
これが、なぜ大発見なのかは、イチョウは、裸子植物(子房をもたず、胚珠が裸出している植物群)であるが、裸子植物や被子植物は、通常、精子を作ることはない。イチョウは、下等な植物の形質を遺存させていた。海で始まった生物が精子を作るのは、下等生物では通例であるが、陸上で繁栄を極めた高等植物では、進化の過程で失われていたはず、と考えられていた。だが、イチョウは陸上植物に進化する前の、海洋植物当時の生態を留めていた、と言う画期的な発見であった。
イチョウの受精方法は、雌の配偶体を雌株に寄生させて栄養供給させ、雄の配偶体を雄株から風媒により雌株の生殖器官である胚珠の先端に送り込んで雌の胚珠に宿り、その胚珠で配偶子を形成して精子を作る。そして、環境条件など準備万端が整う9月になって受精する。この方式によれば、受精が安定して達成される可能性が高くなる。海洋では、配偶子と胞子体を別個に発達させた植物は、陸上では、雌の配偶子を胞子体(雌株)に寄生させると言う、新たに手法に進化させた。
ところが、母体となる雌株に誕生した幼植物を、母体から放出しなければ、「種の保存」という、あらゆる生物が宿命として背負う繁殖に繋がらない。そのために、雌の配偶体を中核して種子を誕生させる形態まで進化した。配偶体は、受精後、種子を母体から切り離すことで、植物の多くは、種子を広く散布できるようになった。しかも、環境条件が悪ければ、休眠してやり過ごすことも可能になった。生殖器官の進化における最終段階が、種子と果実の発達であった。種子と果実で胚を守り、胚に適切な養分を与え、胚はそれらの養分を元手にして、発芽し芽生えを確立する。
種子植物は、全植物の約8割を占めるまでなった。大別すると、裸子植物門と、被子植物門に分かれる。
陸上植物は、やがて葉(花葉floral leaf)を変形させて、受精して種子となる胚珠を包む器となる雌しべを作り、その雌しべを生殖器官の中核とした。雌しべと雄しべを持つ植物が、被子植物であり、真の花の登場は、恐竜が地上を闊歩していた、今から1億4,000万年前と推定される。花は被子植物しか持たない。花弁はその中の特殊な器官であるから、花弁のある植物は全て被子植物と言える。被子植物の種のおよそ7割は、花に雄しべと雌しべの両方がある。
顕花植物には、裸子植物と被子植物とを含み、裸子植物の生殖器官も花と見立てる。種子はシダ植物における大胞子嚢(胞子が、その内部に生じる袋状の生殖器官)と、それを覆う構造から生じたもので、本来は胞子形成を行う葉の裏面に付着していた。裸子植物では、胞子葉(胞子を形成する葉)と区別がつきにくいものもあるため、被子植物の花だけを花と呼ぶ考えもある。英語で Flowering plant と言った場合には、被子植物のみを言う。
裸子植物には花弁がないことは確かである。「花は被子植物が作る生殖器官」という定義がある。その定義からすると、裸子植物には花がないことになる。それでは、裸子植物の花粉や胚珠を作る生殖器官をなんと呼ぶのかというと、これが実は専門用語しかない。そのため裸子植物でも、「花」あるいは「花にあたるもの」と書くことが多い。
マツやスギなどの、現生の裸子植物の多くの種子は、雌花が球果状(松かさ状)となる。現在ではイチョウ類・ソテツ類・グネツム類を除いた多数の種が球果植物に分類され、独立したグループとして扱われることもある。
ただし、裸子植物の生殖器官が花だとしても、花弁は見当たらない。胚珠を抱えた器官や花粉を抱えた器官がそれぞれ雌しべや雄しべに相当するとした場合、その外側に特別な器官があれば萼片や花弁ということになるが、それも見当たらない。
「花弁のある植物は全て被子植物に分類される」。被子植物が作る生殖器官は「花」であり、花弁は花しか作られない。花の中のどの部分を花弁と呼ぶのかは、花という構造体があった場合、その花被を構成する一番外側に並んでいる器官を萼片と定義することになっている。
キンポウゲの仲間のように、花の内側からたどっていって雌しべ、雄しべ、その外に一回りしか器官がないときは、それが黄色く色づいてきれいであっても、萼片とみなされる。花弁は、萼片の内側にもう1つ、雄しべの手前に別の器官があるとき、それを花弁と呼ぶことになっている。そのため、キンポウゲのように萼片はあるものの花弁を持たない被子植物もあり、ミズバショウの花弁のように見える純白のものは、大型化した苞片(仏炎苞)が進化したもの、ドクダミの本来の花にも、花弁も萼もなく、総苞片が支える雌しべと雄しべのみからなる。
裸子植物の花(通例に従う)は単性花で、花粉嚢や胚珠がそれぞれ集合して雄花や雌花を作る。裸子植物に属する原始的な種子植物が出現したのは古生代デボン紀の末、3億6,000万年前で,それから中生代の白亜紀中ごろに被子植物が出現するまで、2億年以上,陸上植物を代表するものであった。裸子植物の花粉媒介は、風に頼る風媒であった。
被子植物の誕生まで、蘚苔(せんたい)類のコケ植物が、陸上植物として古生代シルル紀4億3,000万年に出現してから3億年は経過していたことになる。被子植物は、受精に成功すると、新たな成長を雌しべの子房全体で関わる果実に託す。雌しべは、受精した胚珠から成長した種子を、安全かつ有効に広く散布する手段を獲得した。
雄の胞子も進化を遂げて、花粉となる。花粉を作る雄性の生殖器官を、雄しべと呼ぶ。真の花には、雄しべと雌しべの両方を備えた両性花と、雄しべだけの雄花と雌しべだけの雌花がある。受精後は雌しべは果実として成長を続けるが、雄しべは花粉を放出すれば、その役割は終わる。
 植物の生活環には、雄性配偶体(花粉)と雌性配偶体(胚嚢;卵細胞)と呼ばれる、2つの完全に独立した単相世代の個体が存在する。花自体だけが生殖器官ではない、花は雌雄の配偶体を含み、両配偶体が被子植物の真の生殖組織を作る。
植物の生活環には、雄性配偶体(花粉)と雌性配偶体(胚嚢;卵細胞)と呼ばれる、2つの完全に独立した単相世代の個体が存在する。花自体だけが生殖器官ではない、花は雌雄の配偶体を含み、両配偶体が被子植物の真の生殖組織を作る。植物は、単相世代と複相世代と言う、異なる2つの多細胞世代を交互に繰り返して世代交代を行う。
有性生殖を行う生物の状態を、染色体の数と構成で表現(核相)すると、一組の染色体をもつ状態(減数分裂から受精まで)を単相、二組の染色体をもつ状態(受精から減数分裂まで)を複相という。
生殖の過程に伴う世代交代とは、その核相(染色体数)の交代を言う。そのうちの1つの核相が、それぞれ染色体を2コピー(2Nの染色体)ずつ持つ複相の細胞からなり、もう1つの核相が、減数分裂による細胞の単相化(染色体数が2Nから1Nになること)である。
植物の生活環は、二倍体の胞子体世代と一倍体の配偶体世代という、2つの遺伝的に異なる多細胞体の発生段階から成り立っている。それぞれが世代交代の起点となり、しかも相互依存の関係にある。
植物の世代交代(alternation of generations)は、極めて多様で複雑であるが、単純な藻類から被子植物への生物進化の結晶は、生物学で言う「雄ずい」と「雌ずい」からなる花の生殖構造と、それに貢献する胞子体にあらわれている。
雄ずいは、通常、葯と花糸で構成される。雌ずいは、胚珠を納める子房と花粉が付着する柱頭と、それを支える花柱からなる。
目次へ
植物は自ら移動できないので、受粉・受精の過程は、風や昆虫・動物の生活環に依存する媒体としての運搬体に頼る。植物のこの過程は、単純な依存関係にあるのではなく、他殖(別の個体間の受精を他家受精といい、この形式による生殖を他殖という)を促進するために、形態的及び生化学的な複雑な仕組みを進化させてきた。
野の花の色は、白色系が33%と最も多く、黄色系が28%、赤色系が20%と続き、紫系と青色系を合わせ17%、その他の色が2%と言われている。植物の生殖戦略と関係があるようだ。花と昆虫は、互いに都合のよいように進化してきた。これを共進化と言う。
昆虫の目には、単眼(昆虫類にある小型の眼で、複眼とともにあるか、これに代るものとして存在する。)と呼ばれる器官があり、明暗と色彩で光を受容する。昆虫が受容出来る光の波長は人に比べて短波長よりで、300nm〜650nm (nm: ナノメーター=10-9m= 10億分の1m)の範囲だと言われている。そのため、紫外線を含む青や紫の短波長の光に、昆虫は反応しやすく、赤色のような長波長の光には反応が鈍い。ただ、人には黄色い花や白い花でも、昆虫には淡い青色に写ると見られている。
 |
| ムラサキケマンは全草にアルカロイドの痙攣毒を持つ。その種子の最 外皮にある非水溶性タンニンが、酸化すると種皮を硬くし黒く染める。 |
オレイン酸などの脂肪酸、グルタミン酸などのアミノ酸、ショ糖などの糖を含む、かなり栄養価に富んだ物質であるため、アリは好んで、餌として巣に引き込んでいく。それにより、「ムラサキケマン」の種子は、拡散する。
ムラサキケマンのようなケシ科の植物の殆どは、全草に強い有毒成分を含んでいる。誤って食べると嘔吐・昏睡・呼吸麻痺や心臓麻痺を引き起こす危険がある。それで、アリは、種子まで食べようとはしない。種子は、放置され場所で、やがて発芽する。
ただ、アゲハチョウ科のウスバシロチョウ(ウスバアゲハ)の幼虫にとって、ムラサキケマンは食草となる。ウスバシロチョウは、その毒を体内に保持して、鳥の捕食を免れている。
イチョウのように、受粉のあり方として原始的なのが、裸子植物の風媒花である。やがて昆虫や小鳥が、栄養豊かな蜜や花粉に誘引され、結果的に花粉を媒介する複雑な生殖器官を備える被子植物が主流となった。
昆虫は、蜜や花粉を餌とするため訪花する。花は、そのついでに花粉を運んでもらう。それで、別の花から運ばれた花粉が、雌しべの先の柱頭に付着する。
花冠は、花の形や彩りを変えて、昆虫や鳥を誘引する。日本には、ハナアブ・ハナバチ・ハナバエ・チョウ・ガ・カミキリムシの仲間をはじめ多数の訪花昆虫が生息する。その多種の昆虫に適合するように花も多種多様な構造となる。被子植物と昆虫は、受粉を介して相互に応答し合い、共進化の速度を速めた。被子植物との共存関係がなければ、これほど種々様々な植物や昆虫や動物などは出現しなかったであろう。
果実は食べられても種子は残る。種子は果実が食べられることで、広く散布される。雌しべの登場で、植物は動物との共生の道を歩き始めた。
現在、植物は雨の殆ど降らない砂漠にさえ進出している。サボテンやバオバプも、葉からの蒸散を抑えるなど進化した産物である。バオバプの原生種がマダガスカルに6種、オーストラリアとアフリカに1種ずつ存在する。最大のものは南アフリカ共和国北東部のリンポポ州にあり、高さ47m、直径15mである。年輪が無いため樹齢を知ることは難しいが、数千年に達すると言われる。放射年代測定は可能という。
バオバブの大木は、10トンもの水分を幹に蓄えており、乾季になると葉を落とし休眠する。葉からの蒸散を抑えるためである。休眠中は貯蔵する水分で生き伸びる。バオバブという名の由来は、アラビア語で「果実が多い」という意味であるbu hobab(ブー・フブーブ)がbaobabに変化したものといわれている。バオバブの果実は、ヘチマのように垂れ下がり、堅いが果肉は食用・調味料になる。スーパーフルーツとも呼ばれ、ビタミンCは、レモンの3倍、カルシウムも牛乳より多いと言われている。若葉を野菜として、種子からは油が採集される。
その樹形は、「巨人が幹をつかんで根を引っこ抜き、逆さまにしたようだ」と形容されるが、それはそれで納得できる表現である。その一方では、その周辺に、「サバンナの王」とも称される威風が漂う。
熱帯では、バオバブの花の蜜や花粉を食べるコウモリが、花粉を媒介している。
熱帯雨林の木の幹には、数多くのシダやランなどが着生している。進化の頂点に位置する植物の代表が、ラン科とキク科である。ラン科は、南極をのぞくすべての大陸の熱帯から亜寒帯に自生する。被子植物の中では最も後に地球上に現れた植物である。単子葉植物の科のひとつで、その多くが美しく、独特の形の花を咲かせる。被子植物の中では最も遅れて地球上登場しながら、被子植物の中で、もっとも種数の多い科となっている。
ラン科植物は、世界に700属以上15,000種、日本に75属230種がある。花は左右対称で、虫媒花の中では特異なほど効率の良い花形を発達させ、特定の昆虫によってしか受粉できないほどの共進化を見せている。また根や種子の発芽では、菌との共生が大きな役割を担う。ラン科植物の種子は、ほこりのように細かく、未成熟な胚のみで胚乳もなく、ほとんど貯蔵養分を持っていない。自然下では発芽の際に菌類が共生して栄養を供給する。さらに菌類への依存を強め、自分自身は光合成をせず、菌類にたよって生きる。
ラン科のサギソウは、日本各地の日当たりのよい湿地に生え、親株から走出枝(ランナー)を伸ばして、その先端に翌年の球根を作る。3~5枚の細長い葉を根元近くにつけ、その中心からまっすぐに茎を伸ばして、先端に1~5輪の花を咲かせる。花は純白で、唇弁(しんべん)の先端は3つに割れ、その左右の裂片には多数の深い切れ込みがある。その形が白鷺を思わせる。
ラン科の花粉は、普通の花のように花粉が1つ1つ分離せず、大きな塊となって花粉塊を作り、その花粉塊には長い柄があり、その長い柄の先が粘着体となっている。サギソウは細長い距により、長い口吻を持つスズメガに適応している。サギソウの距の長さとサギソウを訪花するスズメガの口吻の長さが、ほぼ一致している。
ただ、アザミウマ((薊馬))のようなジェネラリストタイプの送粉昆虫が、全種子生産の1/4で貢献していることも観察されている。和名の由来は、「馬出よ」などといいながら、アザミの花を振って、出てきた花粉食のアザミウマを数える、古い時代の子供達の遊びがあった。
鷺草や 風にゆらめく 片足だち (嘯山)
目次へ