| Top 車山高原 車山高原お知らせ 車山ブログ
|
|

DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性
| 目次 |
| 1)細胞体積と膨圧の関係 |
| 2)細胞膜の水透過性 |
| 3)水ポテンシャルΨ勾配 |
 |
 |
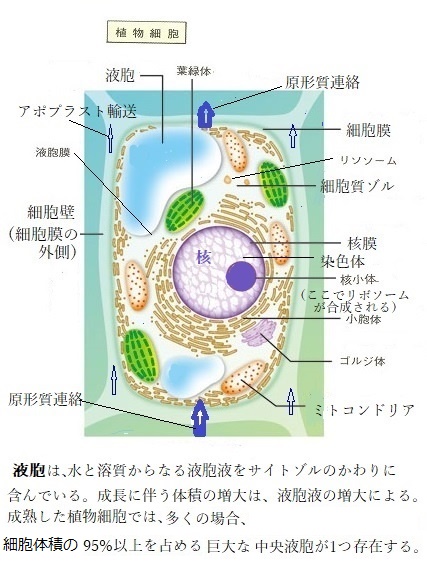
細胞体積の小さな変化が、膨圧に大きな影響を及ぼす。その植物細胞の水ポテンシャルは、光合成に付随して起こる蒸散によって大きく変化する。一方、細胞壁は、植物細胞の体積を一定に保つ大きな役割がある。
植物細胞は、硬い細胞壁でまもられ、細胞の水ポテンシャルΨの変化は、一般的に大きな圧ポテンシャルΨp(水の自由エネルギーに対する静水圧の効果を表す;膨圧)変化を伴うが、Ψpが0より大きいければ、その膨圧によって細胞の体積変化はごくわずかな範囲に留まる。
殆どの細胞の膨圧は、細胞体積が10〜15%減少すると0に近づく。非常に硬い細胞壁を持つ細胞では、膨圧が消失しても細胞体積の減少は小さい。一方、サボテンの茎の中にある水を貯蔵する細胞のように、極端に柔軟な細胞壁を持つ細胞では、膨圧消失による体積の変化は大きくなる。大きな体積弾性率εを持つ細胞の細胞壁は硬く、体積変化が同程度ならば、体積弾性率が小さく柔らかい細胞壁を持つ細胞に比べて大きな膨圧変化が生じる。
体積弾性率(ε;ギリシャ語でイプシロン)は、圧ポテンシャルΨpと細胞体積の関係により計算される。「ε」は、体積変化に対する圧ポテンシャルの変化を言う。
一般に大きなεを持つ細胞の細胞壁は硬く、柔軟性に乏しいため、水分の低下による膨圧変化が大きく、反対にεが小さい細胞の細胞壁は、柔軟性が高いため、水分の低下による膨圧変化も小さいとされている。細胞壁の物性は、植物種や細胞種により異なるため、それにより水不足の細胞体積への影響も大きく異なる。
綿(Gossypium hirsutum)の葉の水ポテンシャルは、含水量が減少し始めると、間もなく急激に低下する。ところが、水ポテンシャルに比べて、溶質ポテンシャルsolute potentialの方は殆ど変化しない。しかし、細胞体積が90%以下に減少すると、その関係は逆転する。水ポテンシャルの低下は、細胞の溶質ポテンシャルΨsの低下に起因し、膨圧は殆ど変化していない。
サボテンは、乾燥地帯を生殖地とする茎が多肉化した植物である。サボテンの茎は、外側の光合成層と内部の水を溜める非光合成組織から構成される。
乾燥すると、これらの2つ組織群の水ポテンシャルは、平衡もしくはそれに近い状態が保たれるが、水は内部の細胞から先に失われていく。
ウチワサボテンの一種Opuntia ficus-indicaの詳しい研究により、貯水細胞は光合成細胞と比べ体積は大きく、細胞壁は薄くしなやかで、体積弾性率εが小さい。εが小さい細胞の細胞壁は、柔軟性が高いため、水分の低下による膨圧変化も小さいとされている。水ポテンシャルが低下すると、貯水細胞は光合成細胞よりも大量の水を失う。
その上、水溶性の糖が、不溶性の澱粉粒へ重合することなどにより、乾燥にさらされた貯水細胞の溶質濃度を減少させる。多くの細胞は、乾燥に応答して溶質を蓄積し、部分的にでも細胞の水分消失を防ごうとする。
しかし、サボテンでは、貯水細胞が、柔軟な細胞壁と、乾燥条件では溶質濃度を減少させる性質を合わせてもち、まず貯水細胞から水を引き出させるようにする。こうして光合成組織では、十分な水分が保持される。
目次へ
 Ψg項は、重力ポテンシャルgravitational potentialと呼ばれる、重力による単位質量あたりの位置エネルギーを言う。重力ポテンシャルは、重力に作用されて得られる位置エネルギーであるため、基準状態の水からの高さ(h)・水の密度(Pw)・重力加速度(g)に依存しているため、Ψg項は、Ψg=Pw g h の式で表される。
Ψg項は、重力ポテンシャルgravitational potentialと呼ばれる、重力による単位質量あたりの位置エネルギーを言う。重力ポテンシャルは、重力に作用されて得られる位置エネルギーであるため、基準状態の水からの高さ(h)・水の密度(Pw)・重力加速度(g)に依存しているため、Ψg項は、Ψg=Pw g h の式で表される。基準状態のPw g hの値は、0.01 MPam-1である。即ち、10mの水の上昇は、水ポテンシャル0.1Mpaの増加に対応する。しかしながら、隣り合う細胞間の重力項Ψgの差は、溶質ポテンシャルΨsや圧ポテンシャルΨpと比べると無視できるほど小さい。重力項は、細胞レベルの水輸送では考慮されないことが多い。そのため簡略化されて、Ψ=Ψs+Ψpで、水ポテンシャルは測定される。
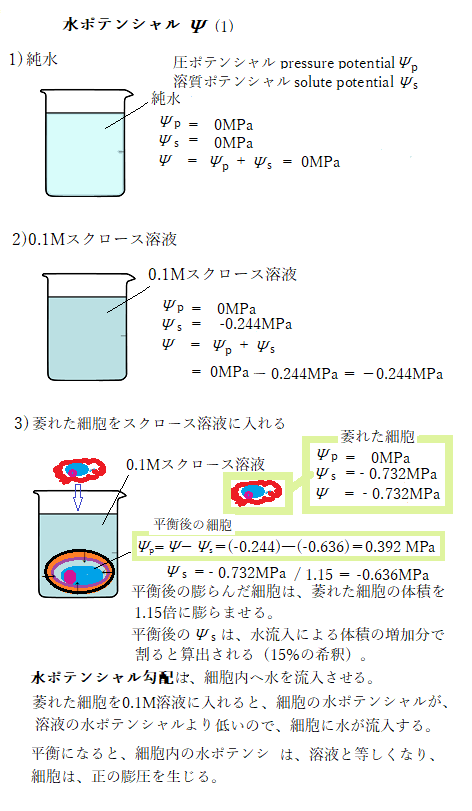
20℃の純水に満たされたビーカーの水は、大気と接しているので、大気圧と一緒で、水の圧ポテンシャルpressure potentialΨpは、0Mpaである。純水のため溶質は含まれていないので、溶質ポテンシャルsolute potentialもΨs=0Mpaである。また、ビーカーの位置も基準の高さに等しいと定義すると、重力ポテンシャルも、Ψg=0Mpaとなり、水ポテンシャルは、Ψ=Ψs+Ψp となり0Mpaとなる。
純水にスクロースsucrose(ショ糖C12H22O11分子量342)を0.1M(モル濃度とは、溶液1Lあたりに、どれだけの溶質solute[mol]が含まれているかを、mol/Lの単位で示す)の濃度に溶かす。
これにより溶液の浸透ポテンシャルosmotic potentialΨsは、-0.244Mpaに下がるため、水ポテンシャルΨも-0.244Mpaに低下する。
溶質soluteは、水を薄めることになり、純水より水の自由エネルギーを減少させる。
植物体内における水移動は、水ポテンシャル勾配(ΔΨ)を原動力とする。例えば、植物細胞が細胞膜の外側から細胞膜の内側へ水を吸収する時、周囲の水が標準状態の純水であれば、Ψ = Ψs+ Ψp= 0 + 0 = 0の状態である。しかし、細胞質は溶質である。純水にスクロースsucroseを0.1Mの濃度で溶かすと、溶液の浸透ポテンシャルΨsは、-0.244MPaに下がり、水ポテンシャルも-0.244Mpaとなり、水ポテンシャル勾配(ΔΨ)は負になる。
これが、溶質である細胞内がΨ= Ψs+ Ψp = -0.244+ 0 = -0.244となれば、この水ポテンシャル勾配を原動力として、水は細胞外に移動する。この水移動がしばらく続き、細胞内の溶質濃度が、3Mとなり、細胞内の正の静水圧(膨圧)が全くない状態まで萎れたとする。萎れた細胞は、大気と接したままであるため、大気圧と同じで、圧ポテンシャルΨpは、依然として0Mpaである。この状態の細胞内の溶質濃度となれば、溶質(浸透)ポテンシャルΨsは、-0.732Mpaに相当する。したがって、細胞の水ポテンシャルは、-0.732Mpaとなる。
この細胞を0.1Mのスクロース溶液が入ったビーカーに入れると、スクロース溶液の水ポテンシャルは、-0.244Mpaであるから、-0.732Mpaの細胞の水ポテンシャルより高いので、水ポテンシャルは、高い所から低い所へ流れるため、それによりスクロース溶液が細胞内へ移動する。
水が細胞内に入ると、細胞膜は膨らむまま細胞壁を圧迫する。細胞壁の弾性的性質のため細胞質に対する壁圧が強くなっていく。細胞壁は、僅かでも伸びる細胞を押し戻すことで変形を防ぐ。その結果、細胞質の圧ポテンシャルΨpが著しく増大し、負のΨSを打ち消すようになる。細胞内の水ポテンシャルΨは増加し、細胞内外の水ポテンシャル差ΔΨは減少する。やがて、細胞内の圧ポテンシャルΨpは、細胞内の水ポテンシャルをスクロース溶液の水ポテンシャルと同じ値まで増加させる。この平衡状態により水ポテンシャル差ΔΨは、0MPaとなり、正味の水輸送はとまる。
細胞壁による膨圧と細胞体積の関係は、細胞壁の弾性により決まる。一方、細胞と外環境との水の出入り速度は、細胞膜と液胞膜の水透過性により決まる。植物細胞の細胞壁はかなり硬いので、小さな体積変化が大きな膨圧変化を生じさせる。細胞内外の水ポテンシャル差ΔΨがゼロにならない限り、膜を介した正味の水の出入りは、減少しながら続く。
この平衡状態では、Ψ(細胞)=Ψ(溶液)と、水ポテンシャルはどこでも等しくなる。ビーカーの容積は、細胞体積よりはるかに大きいため、一個の細胞に吸収された程度の微少の水は、スクロース溶液の溶質濃度には、殆ど影響しない。それでスクロース溶液のΨS・Ψp・Ψは、変化しないので、その平衡状態はΨ(細胞)=Ψ(溶液)=-0.244MPaのままである。
細胞のΨpとΨSの計算には、細胞体積の変化量が加える必要がある。上記の例では、細胞体積は15%増加すると仮定する。それは、萎れた細胞の体積が1.15倍に膨らんだことなる。水が流入する際の細胞内の溶質の数が一定であると仮定すると、溶質濃度は15%希釈されたことになる。
最終的なΨSは、最初のΨSを、水流入により増加した分を加えた体積で割って算出する。
-0.732/1.15=-0.636 MPa となる。
これを Ψ = Ψs+ Ψp の式を並べ替えれば、平衡後の細胞の圧ポテンシャルが算出できる。
Ψp=Ψ−Ψs=(-0.244)―(-0.636)=0.392 MPa
水は水ポテンシャル勾配に従って細胞外にも出る。上記の植物細胞を、0.1Mスクロース溶液から、0.3Mスクロース溶液に移すと、Ψ(溶液)-0.732 MPaは、Ψ(細胞)-0.244 MPaより小さくなり、水は膨れた細胞から溶液へ移動する。
水が細胞から出ていくと細胞体積は減少する。体積が減少すると、
Ψ(細胞)=Ψ(溶液)=-0.732 Mpaになるまで、ΨpとΨは低下する。
細胞から水が流失するときにも、細胞内の溶質の数が維持されると仮定する。細胞体積は15%減少するから、溶質濃度は15%増加するとみられる。当初のΨsに細胞体積の相対量を描けると最終的なΨsが算出される。
Ψs=-0.636×1.15=-0.732 Mpaとなる。これをΨp=Ψ−Ψsに当てはめれば、細胞内の浸透ポテンシャルΨp=0 Mpaとなる。これによって、細胞の膨圧が低下し、スクロース分子が、比較的大きな細胞壁の孔を通って通過する原形質分離が起こる。次に、細胞膜全面を通して細胞質と溶液間に水ポテンシャル差が生じ、プロトプラストprotoplastは細胞壁の影響を受けずに収縮する。プロトプラストとは、細胞膜に包まれた原形質の塊で、細胞融合や高分子物質の取込みといった性質を示す。種間雑種の形成やウイルス感染の実験などに利用する。なお、plastとは、生物学では、「形成された」を意味する連結形である。
細胞が空気中で乾燥すると、原形質分離は起こらないが、細胞質と細胞壁は一体となって収縮するため、その細胞体積の減少に伴い細胞壁は機械的に変形する。
目次へ
細胞の水輸送速度は、細胞膜の水透過性に影響される。水ポテンシャルΨ勾配に従って、水は細胞の内外に移動をする。その流れの方向はΨ勾配によって決まり、水の輸送速度は駆動するΨ勾配の大きさに比例する。
しかし、環境の水ポテンシャルにさらされる細胞では、細胞内外の水ポテンシャル値が近づくと、細胞膜を介する水の移動は減少する。やがて、その速度は、指数関数的にゼロに近づく。
短い半減時間は、速い平衡を示す。速度が半減する時間は、細胞の体積当たりの表面積が大きく、高い透過性と大きない体積弾性率εを持つ硬い細胞壁のある細胞は、環境の水ポテンシャルに応じて素速く平衡に達する。細胞の半減時間の多くは1〜10秒だが、より速いものも知られている。
1つの細胞の半減時間は短く、1分未満で環境と平衡に達する。多細胞組織では、半減時間はかなり長くなる。
速度が半分に減少する時間には、計算式がある。
半減時間(t1/2)= { 0.693 / (A)(Lp)) } (V /εーΨs )
Vは細胞の体積、Aは細胞の表面積を表す。Lpは細胞膜の水透過性(水伝導係数)である。水透過性は、水がいかに速く細胞膜を横切って移動するかを表す。Lpの単位は駆動力あたり、単位時間当たり、単位面積当たりの水の体積を表し、m3m-2S-1MPa-1で示される。
細胞内への水輸送速度は、細胞内外の水ポテンシャル勾配(ΔΨ)と細胞膜の水透過性(Lp)に依存する。
細胞が水を吸収すると、水ポテンシャル勾配は、時間とともに低下し、水吸収速度が遅くなる。半減時間(t1/2)は、細胞パラメーターparameterとして、体積V・表面積A・水伝導係数Lp・体積弾性率ε、及び浸透ポテンシャルΨsを指数関数にして減少させる。
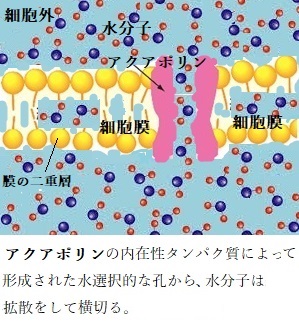 アクアポリンaquaporinは、細胞膜に存在するタンパク質の一つで、細胞へ水分子を選択的に通す細孔を持ち、細胞への水の出入りに重要な役割を果たしている。大部分の細胞では、水は細胞膜の脂質を通ることで細胞に出入りする。しかし、いくつかの上皮細胞では水の透過性が高かったため、何か別の機構が存在することが予想されていた。
アクアポリンaquaporinは、細胞膜に存在するタンパク質の一つで、細胞へ水分子を選択的に通す細孔を持ち、細胞への水の出入りに重要な役割を果たしている。大部分の細胞では、水は細胞膜の脂質を通ることで細胞に出入りする。しかし、いくつかの上皮細胞では水の透過性が高かったため、何か別の機構が存在することが予想されていた。
それが、1991年、当時ジョンズ・ホプキンス大学にいたアグレによって発見された。それまでは、脂質二重層で直接横切る拡散だけでは、生体膜を横切る水の輸送量の多さを説明できなかった。
アクアポリンは、水選択的な細孔で、膜内在性タンパク質で形成されている。水は、脂質二重層よりも速く、このチャンネルで拡散できるので、アクアポリンは、植物細胞内への水輸送を促進する。最近の研究では、水以外のCO2など様々な小分子を輸送するアクアポリンも報告されている。
陸上植物内における外気から葉緑体までのCO2の拡散は、その光合成にとって重要な要素である。無機炭素が糖に取り込まれるまで、CO2は細胞壁・細胞膜・サイトゾル、そして葉緑体包膜の4つのバリアを通過しなければならない。
葉肉細胞のアクアポリンでは、そのCO2拡散の抵抗を下げるために機能している。葉内細胞間隙のCO2の拡散抵抗は、葉肉抵抗と呼ばれる。葉が活発に光合成を行っている場合の細胞間隙のCO2濃度は、外気のCO2濃度が400ppmの時には、その60〜85%程度以下になっていなければならない。このように、CO2に対する拡散抵抗を下げることは、光合成を律速する重要な要因となっている。
アクアポリンの水選択的な孔により、膜を横切る水輸送の速度を変化させるが、その方向や駆動力は変化させられない。しかし細胞内のpHやCa2+レベルなどの生理的パラメーターに反応して、可逆的に開閉することで、植物は細胞膜の水透過性を制御できる。
植物細胞内部は、わずか脂質2分子分の厚さしかない、ごく薄い細胞膜により、変化の厳しい外部環境から、比較的安定した内部環境を維持している。細胞膜は、核や細胞質の拡散から疎水的バリアとなって守っているだけでなく、細胞が必要とする栄養を取り込む一方、細胞内物質やイオンの内向きと外向き双方の輸送transportを担い、しかも膨圧の調整も細胞壁だけに頼っているのではなく、細胞膜も常に、その制御に関与している。
細胞外から細胞内へ、あるいは細胞内の局所的輸送の多くも、膜タンパク質によって調節されている。細胞膜は、外部の物理的環境の変化や他の細胞からのシグナル分子による情報、そして攻撃を仕掛ける病原体に関する情報などを伝える役割も果たしている。
植物器官の間や、植物と環境の間で受け渡される大量の輸送も、細胞レベルでは細胞膜で調節されている。葉から根への篩管を通るスクロースの輸送(転流translocation)も、葉にある篩管細胞内の膜輸送系で調節され、篩管から根へと輸送されている。
目次へ
![]()