| Top 車山高原 車山高原お知らせ 車山ブログ
|
 植物における根は、肥料成分や水を吸収するための構造であると共に、植物体を支える役割を果たす。樹木では深く広く土壌に侵入する。草の根の方は、それほど深くない。 その形は植物の種類によってもある程度決まっているが、地形や土質によって変化する。根の細胞のための栄養分は、地上部から師管を通じて送られる。 根を構成する細胞も呼吸をする。酸素を吸収し、二酸化炭素を排出する。ガス交換は一般には根の表面で行われる。土壌間隙から酸素を直接取り込んでいる。地上植物の根では、このような状態で、酸素呼吸には十分である。 しかし、根の表面全体が水と直接接し、水に溶解している酸素を吸収するような条件では、根表面に接する水の酸素濃度はすぐに低下する。 酸素の供給は、拡散に頼ることになる。水の中での酸素拡散の速度は遅いため土壌からの酸素の供給がほとんどなくなり、根は柔細胞間隙からだけの酸素供給に頼ることになる。したがって、湿地や水中では根は深く侵入できないことが多い。 湿地に森林が成立しにくいのはそのためである。一部の植物では呼吸根を出し、あるいは、根の内部に空気の通る管を形成してこれに対応している。 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力
| 目次 |
| 1)重力が関係する表面張力と濡れ性 |
| 2)土壌中の水 |
| 3)根の構造と機能 |
| 4)キャビテーション |
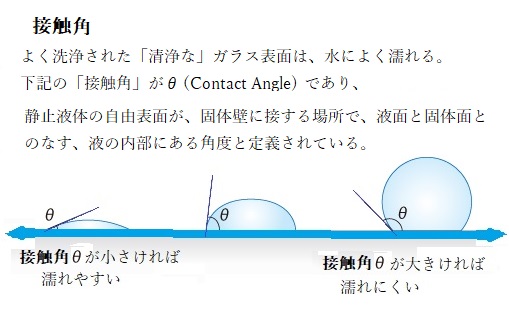 一般的に表面張力と濡れ性の関係が、液体と固体との濡れの現象にどのようにして関わるのか?
一般的に表面張力と濡れ性の関係が、液体と固体との濡れの現象にどのようにして関わるのか?濡れ性、すなわち固体表面上の液滴の接触角θを支配しているものは、固体の表面張力γS、液体の表面張力γL、固体と液体の界面張力γSLの3のバランスで決る。
これを数式で表したものが、イギリスの物理学者 トマス・ヤングに因んだヤングの式Young's equationである。
γS = γL ・cosθ + γSL (θ;シータ)
一般的に表面張力とぬれ性の関係は、液体の表面張力より、固体の表面張力の方が強い時、接触角が小さい(ぬれやすい)。液体の表面張力の方が、固体の表面張力より強いとき、接触角が大きくなる(ぬれにくい)。この関係は接着の制御など、様々なアプリケーションに活用されている。
コップの縁すれすれのところまで水を注ぎ、さらにゆっくりと注いでいくと、水面が盛り上がる。この現象が、表面張力による液面の盛り上がりであることはよく知られている。 水の質量により、地球上では重力が働く。重力は、物体の位置をできるだけ低いところに保とうと作用する。水をゆっくり注いで水面の高さが徐々に高くなっているときにも、重力は常に働いている。それで、注がれた水は、その水面の高さをできるだけ低い位置に留めようとする。さらに水が注がれると、水面の高さをできるだけ低く保つようにするが、コップの中には水が満たされていて水の行き場はない。やがてこぼれる。この状態では、重力が水面を上方から押さえつけていると考えてもよい。 しかし実際には、水を注いでいった時に、コップの縁からすぐにこぼれるのではなく、ある程度の高さまで水面が盛り上がる。
水面を押さえつけようとする重力の作用に抗して、別の力が水の表面に作用するのが表面張力である。水を注いでいった時に、水の表面張力によってある程度の高さまでは水面が盛り上がるが、さらに注ぎ続けると水はコップの縁からこぼれてしまう。水面が高くなると、水面を押さえつけようとする重力の作用がさらに大きくなって、表面張力では、これを支えきれなくなるためである。
表面張力は、水だけでなく、一般の液体の表面にも作用し、さらに固体の表面にも作用する。 液体の表面とは気体/液体の界面、固体の表面とは気体/固体の界面のことであるから、「液体の表面張力」をもう少し正確にいえば、「気体/液体の界面にはたらく界面張力」ということになる。
「固体の表面張力」も同様に、「気体/固体の界面にはたらく界面張力」ということになる。界面はこのほか、液体/液体の界面・液体/固体の界面・固体/固体の界面があり、それぞれの界面には界面張力が働く。
ところが現実には、上記の関係が成り立たないケースもある。このような場合には、表面自由エネルギーの成分分けの概念を導入した手法が有効となる。
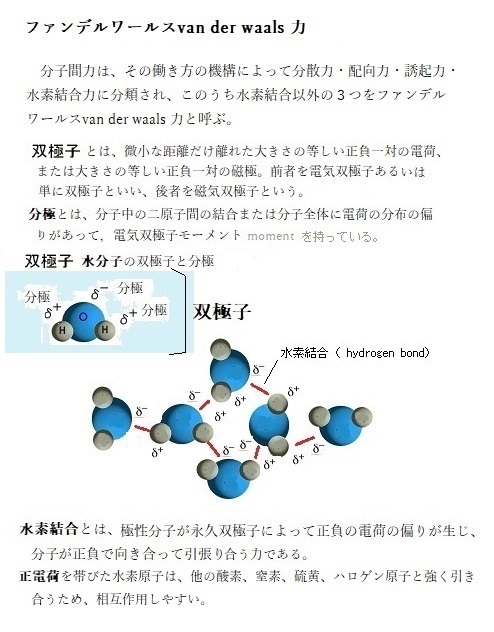
分子間力は、その働き方の機構によって分散力・配向力・誘起力・水素結合力に分類され、このうち水素結合以外の3つをファンデルワールスvan der waals 力と呼ぶ。オランダの物理学者ヨハネス・ファン・デル・ワールスの名前から付いたファンデルワールス引力は、水素結合よりも弱いが、2個の原子が引き合えば、両原子のファンデルワールス半径の和に等しくなるまで結合するため、多数の原子が集まり、表面形状がぴったり合う巨大分子同士の結合が可能となる。
そのため重要な機能を持つようになる。実際の原子は、非常に小さい原子核とその周囲を取り巻く電子雲からなる非常に疎な構造を持つ。しかし、原子がある半径以内では堅いものと想定することで様々な用途に応用できる。
ファンデルワールス力半径は、単体の結晶をつくる元素について、隣接する原子同士の距離を2で割ることで算出される。ファンデルワールス半径は、ファンデルワールス力で原子、または分子が凝集した時の、互いの距離から計測され、これ以上近寄ることは出来ないという位置に凝集した状態で安定していれば固体になる。その結晶構造と格子間隔が分かれば、ファンデルワールス半径は、算出できる。
原子半径・イオン半径・共有結合半径なども同じ発想である。水素の共有結合半径はH2分子の結合距離の半分であり、その結合距離は実験的に測定される。原子同士の距離は、X線回折装置などを利用して計測する。 X線回折装置は、結晶性物質に原子間距離と同程度の波長を持つ単色X線(一つの波長のみから成るX線)を照射した際、X線が原子の周りにある電子によって散乱、干渉した結果起こる回折を、解析することを測定原理としている。この回折情報を用いることにより、分子の三次元構造や結晶性物質の構成成分の同定や定量、結晶サイズなどが測定可能となる。
表面自由エネルギーが分子間力の働きに起因する以上、ここでもその機構に基づいた分類が考えられる。これが表面自由エネルギーの成分分け( γ = γd + γp + γh )の概念につながり、表面張力の大きさだけで、ぬれの関係が説明できないような現象の解明に役立つ。
 分子間力と表面自由エネルギーの成分分けの関係を示すと、
分子間力と表面自由エネルギーの成分分けの関係を示すと、「分散力」
無極性分子(中性分子)には電荷の偏りがないが、電子は電子雲からなる非常に疎な構造を持ち、絶え間なく動いているため、ある瞬間をとらえた時、電荷の偏りが生じる。この瞬間的な双極子の誘起により誘起双極子として分極する。誘起双極子induced dipole(ˈdaɪˌpoʊl)が、他の分子も誘起双極子として誘起させ相互作用をする。全ての分子は、常に振動することから、瞬間的に電荷の偏りが生じて誘起双極子が発生する。この分子同士が引張り合う力を分散力と呼ぶ(表面自由エネルギー分散成分γd)。
「配向力(はいこうりょく)」
この力は双極子-双極子の配向に基づくもので、双極子間の電荷の偏りに依存した静電的な力である。結晶中のイオンのように静止している電荷が作る電場を静電場と言う。
双極子モーメント(永久双極子permanent dipole moment)を有する極性分子間に、正負の電荷の偏りが生じ、正負で向き合って引張りあう引力を配向力orientation forceと言う。一般に、分子間の配向は、温度が上昇すると、熱運動の増大により乱れてくるので、配向力は高温ほど小さくなる(表面自由エネルギー双極子成分γp)。
「誘起力」
永久双極子と誘起双極子の相互作用を言う。永久双極子モーメントをもつ極性分子が他の分子に接近すると、その分子に電荷の偏りが現れ誘起双極子が生じる。この誘起双極子と永久双極子の間で作用する引力を誘起力(induction force)という。誘起力は永久双極子の持つ双極子モーメントの大きさと相手分子の分極率に比例するが、温度には依存しない。原子核は変形しないので、変形するのは電子の分布状態(電子雲)である。分子が電場に置かれた時、全体がどの程度変形しやすいかを示す尺度が分極率である。極性分子が無極性分子に接近すると、無極性分子が双極子を誘起することで電荷の偏りが生じて発生する力であるが、非常に小さいため、無視できる。
「水素結合力」とは、F-H、O-H、N-H分子など、電気陰性度が大きな原子と水素原子が結合すると、その強い極性により、水素原子が他の分子の電子供与性の高い原子と結びつく力を言う。van der waals 力よりも強い力を持つ。表面自由ネエルギー水素結合成分γhで示す。
炭素に対して酸素・窒素・硫黄・ハロゲンは電気陰性度が高い。これらの原子が炭素と結合すると電化に偏りが生じ、電気的双極子を生じる。電気陰性度が異なる原子同士が結合していると、わずかであるが電荷に偏りが生じる。図のように、H-O結合では、酸素原子の方が電気陰性度が高いため、水素原子の電子を引き付ける。そのため、酸素原子が少しマイナスの電荷を帯び、水素原子が少しプラスの電荷を帯びることになる。
電気陰性度が強いということは、電子を引き付ける力が強いということである。電子はマイナスの電荷を帯びているので、電気陰性度が強い酸素原子の方がマイナスの電荷を帯びるということが理解できる。このように正電荷と負電荷の偏りが非常に近くで存在している状態を分極といい、分極している分子を双極子という。
炭素に対して酸素、窒素、硫黄、ハロゲンは電気陰性度が高い。これらの原子が炭素と結合すると電化に偏りが生じ、電気的双極子を生じる。分極している分子を、中性分子に近づけると、中性分子に双極子が誘起される。双極子が誘起された中性分子(誘起双極子)は永久双極子と相互作用することになる。その引き合う相互作用を双極子-誘起双極子相互作用という。
マイナスの電荷とプラスの電荷をもつものは、お互いに引き寄せられる。双極子もプラス・マイナスの電荷をもっているので、対応する電荷が来ると引き寄せられてしまう。この双極子が「イオンの電化」によって引き寄せられる場合、イオン-双極子相互作用が生まれる。また、「他の双極子」に引き寄せられる場合は双極子-双極子相互作用が生まれる。双極子の電化は、イオンほど強くないのでイオン-双極子間相互作用の方が結合が強い。
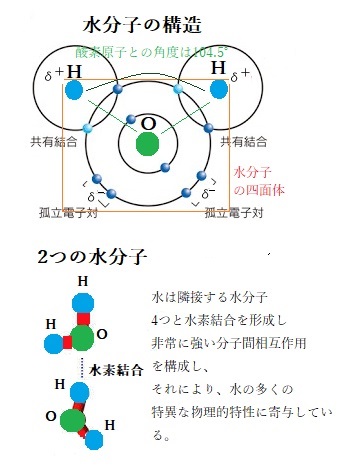
一般に、分子と分子との間には、分子間力と呼ばれる分子同士が互いに引き合う力が作用する。液体の表面張力、その力の大きさは、分子の種類・分子間の距離・分子の向き(配向)によって決まりる。水の固体である「氷」は、水分子の分子間力によって分子同士が互いに引き合い、多数の分子が結合した状態にある。それぞれの水分子の位置は、殆ど固定して、ある点を中心に振動しているだけの「固体」の状態にある。
しかし、温度が高くなると、水分子の熱運動が激しくなり、分子は位置を変えながら、ある程度自由に動けるようになるが、まだまだ分子間力による引き合いの効果のほうが大きいため、分子と分子がばらばらになることはない。これが「液体」の状態である。
さらに温度が高くなると、水分子の熱運動がさらに激しくなり、分子間力による引き合いを振り切って完全に自由に移動できるようになる。これが「気体」の状態であり、この水の状態が「水蒸気」である。また、気体の状態では、分子がばらばらになるため、表面というもの自体が生じないため、「気体の表面張力」という状態もありえない。
このような、固体・液体・気体という「物質の3態」には、分子間力intermolecular force(ɪ̀ntɚməlɛ́kjəlɚ fˈɔɚs)が大きく関わっているが、実は表面張力にも、分子間力が大きく関わっている。
液体の表面付近における分子の状態を考えてみると、まず、液体のバルクBulk(物体・流体のうち、界面と接しない物質本体)に存在する1個の分子(図ではB)に着目する。この分子Bと、その周辺に存在するあらゆる分子との間には分子間力が働いている。液体中の分子は、他の分子からの分子間力の作用を受けながら、ほぼ無秩序に熱運動している。したがって、それぞれの分子に働く分子間力は、大きさも向きも様々である。
分子Bにしても、様々な方向から様々な大きさの分子間力が働いている。それでも、例えば1秒間ほどで区切って、その時間内の平均的な分子間力は、あらゆる方向から同じような大きさの分子間力が常に働いているので、結果的にはバランスが保たれている。バランスが保たれているということは、安定している状態を言う。
一方、気体/液体の界面の表面に存在する分子(図ではS)に着目すると、この分子にも、両隣やバルク側の分子から分子間力が働くが、気体側の分子からは、殆ど分子間力が働かない。気体側の分子数密度は、バルク側のそれと比較して、分子それぞれが熱運動をしてバラバラで、その分子間力は無視できる。
その結果、分子Sは、分子Bのようにバランスが保たれておらず、常にバルク側に引き込まれることになる。このような現象が、表面のいたるところで起こっている。分子Sのような表面の分子は、分子Bのようなバルクの分子と比べて不安定であると言え、表面に留まり続けられるほど安定していない。表面に存在している分子には、常にバルクに潜り込むように他の分子間力がインバランスimbalanceに働いている状態にある。その結果、液体の表面積をできるだけ小さくしようとする現象が起こる。即ち、表面には、その表面積を小さくしようとする「張力」となり、これが「表面張力」として作用する。
液滴の体積が一定であれば、表面積が最小となる形状は球である。宇宙空間のような無重力状態では、液滴に対して重力は作用しない。表面張力以外の外力が働かなければ、液滴表面に働く表面張力が、できるだけ表面積を小さくしようとするから、液滴の形状は完全な球状になる。
地球上のように重力が作用する場合は、液滴の体積がごく小さければ、その質量も小さいので、重力よりも表面張力による作用の方がまさる。そのため、液滴形状は球に近くなる。
次第に、液滴の体積が大きくなると、その質量も大きくなり、重力の作用が働き、液滴形状は潰れていく。その接触角は小さくなる。
分子間力の大きな液体では、それ自身の凝集力が大きいということを意味する。表面の分子をできるだけバルク側に移動させて表面積を小さくしようとする作用が大きければ、結果的に、その液体の表面張力も大きくなる。
それが、固体に対する濡れという現象にあらわれる。液体の表面張力が大きいほど、それ自身が広がるよりも凝集して丸まろうとする傾向が強く、固体表面に対して濡れにくいという現象として現れる。
目次へ
 降雨が土壌表面に達すると、その大部分は土壌に浸透して、一 時的に土壌中に含まれる。含水された水の一部は、植物に吸い上げられる。他の部分は下方に移動するか、蒸発によって直接、大気圏に還る。
降雨が土壌表面に達すると、その大部分は土壌に浸透して、一 時的に土壌中に含まれる。含水された水の一部は、植物に吸い上げられる。他の部分は下方に移動するか、蒸発によって直接、大気圏に還る。土壌が雨や潅水によって十分に水に満たされている場合、水は重力により土壌粒子間の間隙を通って、間隙中の空気と部分的に置き換わる。あるいは空気をとらえて下に浸み込んでいく。粘土のように粒径が小さければ、粒子間の通路はより狭くなるため保水性が高くなる。砂や礫では、粒径が大きいので、粒子間に大きな通路ができ、保水性が低くなる。
水は毛管現象capillarity (k`æpəlˈærəṭi)によって土壌粒子間に引き込まれるので、小さな粒子間の通路から先に満たされていく。そのため粘土質の土壌では、多くの水が重力に抗して保持される。土砂降りの雨の数日後でも、粘土質の土壌では体積の40%が水である。砂や礫では、粒子間の空間が広すぎて、水は隙間から流出して、僅かに粒子の表面か、粒子間の通路がごく狭い空間に留まっている傾向がある。そのため水浸しになったとしても、その体積の15%の水しか含まれていない。
土壌粒子と土壌の水との相互作用は、水分子と土壌表層を構成する分子、およびイオンそれぞれに固有な、種々の特質と質量massによって起きる。この相互作用は、土壌が水を吸い込み、それを浸透させ保持し、植物に供給するなどの具体的な能力として現われる。それが、土壌に種々の性質、吸湿性・透水性・保水能・毛管上昇能・容水量などを決定づけることになる。例えば、砂礫など粗粒物質の地層は、粒子間の空隙が大きいため透水層となる。
溶液を等しい体積の2つの区画(AとB)に分ける平面を仮定する。すべての分子はランダムに動いているので、いずれの溶質分子も一定時間にある確率で仮想平面を横切る。ある一定の時間にAからBへ移動する分子数は、その時間内の最初のAの分子数に比例する。またBからAへ移動する分子数は、Bの分子数に比例する。
もしAの濃度がBより高いなら、BからAへの溶質分子の移動よりも、AからBへ移動の方が多い。この場合、AからBへ溶質の正味の移動が起きていることになる。従って、各々の分子がランダムに動いても、拡散は濃度の高い方から低い方へ正味の分子移動が起こる。各々の分子は独立して動くから、それぞれの区画内の分子数は等しくなり、正味の移動が少なくなる。この平衡状態では、2種類の分子がランダムに均一に分布する。当然、すべての地点と時刻で溶質中の2分子それぞれの合計は一定である。
日本列島ように湿潤先端が消滅し、表層から下層に向かう平衡化が進む条件下では、降雨後、長時間にわたって下降運動が行われ、下層に水が貯留されるようになる。
土壌の水を飽和状態にして、蒸発を行なわせると、ある時期まで乾燥速度が一定で、その後、乾燥速度が低下して行く。前者を恒率乾燥期間、後者を減率乾燥期間と呼ぶ。
この期間、水は液相liquid phaseの形態で表面まで移動し、蒸発は土壌表面で行われる。乾燥速度は、外部の乾燥条件によって決定され、土壌内部の水移動に依存しない。この間の土壌内部の水の分布は、水の流動抵抗が小さいため平衡状態に近い状態にある。
 土壌の水ポテンシャル(Ψ = Ψp + Ψs + Ψ g)も、植物細胞の水ポテンシャルと同様に、浸透ポテンシャル・圧ポテンシャル・重力ポテンシャルから構成される。土壌の浸透ポテンシャルosmotic
potentialは、塩を多く含む土壌(―0.2Mpa以下)でなければ、一般的に溶質濃度を無視できる程度で、通常、―0.02Mpaである。また、圧ポテンシャルpressure
potential も湿った土壌であれば「0」に近い。土壌が乾燥していくとΨpは低下し、負になるが、この土壌の陰圧はどこから生じるのだろう。
土壌の水ポテンシャル(Ψ = Ψp + Ψs + Ψ g)も、植物細胞の水ポテンシャルと同様に、浸透ポテンシャル・圧ポテンシャル・重力ポテンシャルから構成される。土壌の浸透ポテンシャルosmotic
potentialは、塩を多く含む土壌(―0.2Mpa以下)でなければ、一般的に溶質濃度を無視できる程度で、通常、―0.02Mpaである。また、圧ポテンシャルpressure
potential も湿った土壌であれば「0」に近い。土壌が乾燥していくとΨpは低下し、負になるが、この土壌の陰圧はどこから生じるのだろう。水は空気―水界面の面積を最小にしようとする強い表面張力を持つ。一方、水は付着力により土壌粒子の表面に密着しようとする。土壌の含水量が減少していくと、水は土壌粒子間の隙間に後退して、空気ー水界面の面積を小さくする傾向と土壌粒子への水の誘引のバランスを表す曲線を持つ空気―水面界を形成する。湾曲した表面を持った水は、以下のような式で概算できる。
Ψp = -2 T/ r
Tは、水の表面張力(7.28 × 10-8MPam)、rは空気―水界面の曲率半径(曲線を局所的に円弧とみなしたときの円の半径をその点における曲率半径と言う。)である。土壌が完全に湿潤状態であると仮定された場合の毛管現象の式と同じである。
土壌が乾燥すると、まず水が土壌粒子間の大きなスペースからなくなり、引き続いて土壌粒子間や内部の小さなスペースからなくなっていく。この過程で、土壌水のΨp値は、連続的に径が小さくなっている孔の空気―水界面の曲率半径が減少するため、大きく低下することになる。
例えば、最も大きい粘土粒子ほどの大きさの
r = 1μm であれば、Ψp値は、-0.15MPaに相当する。その値も、空気―水界面が粘土粒子間より小さな空間の中に後退していくと、簡単に-2から-1 MPaに達するようになる。
3つ目の構成要素は、重力ポテンシャルgravitational potentialΨ gである。重力は排水において重要な役割を果たす。水の下方へ移動は、Ψgが高さに比例することにより、高いほど高く、低いほど低い。
体積流または質量流は、ひとまとめの分子の協調した移動であり、体積輸送bulk transportは、水や溶質を体積流により、通常は、圧力勾配に応答して輸送する。木部や篩部で見られる。庭のホースによる放水や川の流れが、体積流としてよく知られている例である。
土壌水の圧力は、屈曲した空気―水界面の存在による。水は、水が満たされた空間が大きくΨpの高い領域から、含水量の少ない、つまり水が満たされる空間が小さく、空気―水界面がより屈曲したΨpの低い領域に流れる。水蒸気の拡散も、ある程度の水の移動を担い、乾燥中の土壌では重要である。
植物は、土壌から水を吸収するので、根の表面近くの水を減少させる。これにより根の表面近くのΨpが低下し、隣接する土壌のΨpの方が高くなり圧力勾配が形成される。土壌の水で満たされた空間は、互いに繋がっている。水は圧力勾配に従って通路を通って、体積流として根の表面まで移動する。
土壌中の水の移動速度は、2つの要素に依存する。土壌中の圧力勾配の大きさと土壌の水透過性である。水透過性は、土壌中の水の通りやすさの尺度となる。土壌の種類と含水量によって変化する。砂質の土壌では、粒子間の間隙が広いため、水透過性が大きい。粘土質の土壌では、粒子間の間隙が狭いため、水透過性は小さい。
土壌の水分量が減少し水ポテンシャルが低下すれば、土壌水の透過能力も劇的に低下する。それは、土壌間隙の水が空気に置き換わったことで、水の進路の多くが空気に満たされていくと、水の流れは、空気に満たされた狭い通路に妨げられため、土壌水の透過能力も大いに低下する。
目次へ
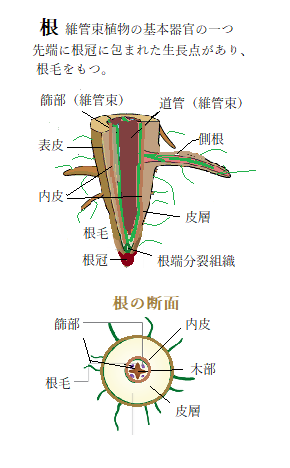 根は植物の体を基盤に固定し、支持し、またそこから成分を吸収する構造である。より厳密には、維管束植物の地下部分であり、茎から出て地中に伸びる棒状で放射相称の構造である。真の根は維管束植物vascular
plantだけの独自の構造であり、それ以外の群ではみられない。維管束植物とは、木部と篩部から構成される維管束をもつ植物、即ちシダ植物・裸子植物・被子植物を言う。
根は植物の体を基盤に固定し、支持し、またそこから成分を吸収する構造である。より厳密には、維管束植物の地下部分であり、茎から出て地中に伸びる棒状で放射相称の構造である。真の根は維管束植物vascular
plantだけの独自の構造であり、それ以外の群ではみられない。維管束植物とは、木部と篩部から構成される維管束をもつ植物、即ちシダ植物・裸子植物・被子植物を言う。水は、植物が成長し機能するために必要な資源の中で、最も豊富であるが、農業生産が阻害される最大要因にもなる。農業生産のために灌漑をするのは、水が農作物にとって重要な資源となるからだ。植物は非常に大量の水を必要とするため、水利用は、生産性においても制限要因となり、降水量によって植生様式に際立った違いが生じる。
根で吸収された水の殆ど(~97%)は、植物体を通して運ばれ、葉の表面から蒸散transpirationする。根で吸収された水のごく一部が、成長(~2%)、または、光合成や他の代謝過程の生化学反応に消費される。
大気への水の消失は、陸上で光合成を行うには避けることはできない。二酸化炭素の取り込みと共通した拡散径路を通して水分を消失する。二酸化炭素が拡散により葉の中に取り込まれると、水蒸気は、拡散により葉から出ていく。
葉が水分消失を駆動する勾配は、二酸化炭素取り込みの勾配よりもずっと大きい。1分子の二酸化炭素を獲得するのに400分子の水を失う。このような大きな水の消失は、植物の形態と機能の進化に大きな影響を与え、水が植物の生理機能に重要な役割を果たしていることを表している。
根の基本的な構造は、少なくとも種子植物では、分類群が異なってもそれ程の差はない。これは主として地中器官であり、その必要とされる機能が殆どの群で違いが見られないために、進化の速度が遅いのだと思われる。根は茎の下方向の延長としてある場合と、茎および根の側枝・側根として出る場合がある。
水中で進化した植物が、陸上で生存するには、いくつかの切実な課題を克服しなければならなかった。特に水の獲得と保持である。その陸上環境に適応するため、植物は根と葉を進化させた。根は植物を固定して支え、水と養分(ミネラル)を吸収する。葉は光を受けてガス交換をする。植物が大型化すると、根と葉の距離はますます遠くなる。このため根と地上部の間では、根で吸収した水や栄養を葉に送り、葉で合成された光合成産物を根に送る効率的な長距離輸送システムを進化させてきた。
 木部は、植物の地上部分に根系からの水やミネラルを輸送する組織であるが、篩部は根を含む成長している組織や貯蔵組織に、成熟葉から、光合成産物特に糖を輸送する組織である。篩部はまた、食物ホルモンやタンパク質などの制御分子の輸送を介してソースとシンクとの間でシグナル分子を伝送し、植物体全体への水及び種々の化合物の再配分を行う。これらの分子のすべてが、篩部を通して輸送される糖やタンパク質と共働する。
木部は、植物の地上部分に根系からの水やミネラルを輸送する組織であるが、篩部は根を含む成長している組織や貯蔵組織に、成熟葉から、光合成産物特に糖を輸送する組織である。篩部はまた、食物ホルモンやタンパク質などの制御分子の輸送を介してソースとシンクとの間でシグナル分子を伝送し、植物体全体への水及び種々の化合物の再配分を行う。これらの分子のすべてが、篩部を通して輸送される糖やタンパク質と共働する。篩管を介して再配分される化合物には、木部を通してはじめて成熟葉に運ばれるもの、修飾されずに成熟葉からそのまま再配分されるもの、再配分の前に代謝返還を受けるものなどがある。
篩管液は、「上向き」または「下向き」にだけ輸送されるのではなく、また、その輸送方向は重力に従うものでもない。篩管液は、糖のソースと呼ばれる供給源から、シンクと呼ばれる代謝や貯蔵の場へ輸送される。
ソース(source)は、糖を組織外へ輸送するすべての器官をさし、多くの場合、自身が必要とする以上の光合成産物の合成能をもつ成熟葉を指す。光合成産物という用語は、光合成により同化され生成物を言う。もう一つのソース組織の例に、貯蔵物質を他の器官へ輸送する貯蔵組織がある。たとえば、二年生のテンサイ(砂糖大根)の野生種の貯蔵根は、一年目にはシンク(sink)としてソース葉から輸送されてくる糖を貯蔵する。2年目の成長期には、同じ貯蔵根が、今度はソースとして機能する。糖は地上部へ輸送されて新しい茎の成長に使われ、その茎は生殖成長を行うようになる。
シンクは、植物のすべての非光合成組織と自らの成長や貯蔵に必要な光合成産物を生み出さない光合成組織である。根・塊茎・成長中の果実、および未成熟葉は、正常な発達に炭水化物の供給が必要であるから、すべてがシンク組織である。
篩部輸送は、単純にソースからシンクへの輸送であると言えるが、特定の輸送経路はそれほど単純ではなく、シンク組織とソース組織の距離、発展段階や維管束の繋がり方、輸送経路の変化などにより左右される。すべてのソース組織が、すべてのシンク組織へ糖を供給するわけでなく、特定のソースから、特定のシンクへ供給される傾向がある。
根の表面からは根毛(こんもう)と呼ばれる、ごく細かい毛状の突出物が出る。これは、根の表層の細胞から生じる突起で、水や栄養の吸収面積を大きくする役割がある。
植物における根は、肥料成分や水を吸収するための構造であると共に、植物体を支える役割を果たす。樹木では深く広く土壌に侵入するが、草の根はそれほど深くない。その形は植物の種類によってもある程度決まっているが、地形や土質によって変化する。根の細胞のための栄養分は地上部から篩管を通じて送られる。
ガス交換は一般には根の表面で行われる。したがって、湿地や水中では根は深く侵入できないことが多い。湿地に森林が成立しにくいのはそのためである。一部の植物では呼吸根(こきゅうこん)を伸ばして、根の内部に空気の通る管を形成してこれに対応している。呼吸根は、気根の一種で、地下部の空気が乏しい環境で根腐れを防ぐために地下から地表へと突き出した根である。水草や湿地に育つ植物(ヌマスギ、イチョウ、あるいはオヒルギなどのマングローブ植物)などに見られる。
光合成を活発に行なっている植物は、同時に多量の水を蒸散によって大気中に放出している。蒸散によって生じる葉の水ポテンシャルの低下が木部液に張力を発生させ、これによって根から葉に水が引き上げられ、土壌からの植物細胞へ水の供給が行われる。それが、葉における蒸散によって根系から葉へ植物体の木部内をマスフローmassflowとして流れる水の移動を指す。マスフローとは、植物の蒸散作用によって、水が根の近傍へ運ばれる際に物質も同時に運ばれることを言う。
葉における蒸散によって、葉の水ポテンシャルが低下し、葉の木部道管内では、圧ポテンシャルが低下する。このとき、根付近の木部内の水との間に生じた圧ポテンシャル勾配を原動力として、茎や幹の木部道管内で液体の水がマスフローとして移動し蒸散流を形成する。
水は水素結合によって水分子同士、あるいは道管壁と強く結びつく力となる凝集力が非常に強い。それらの水を凝集する力によって、水柱内部がたとえ負の圧力となっても、容易に気泡が発生しない。蒸散によって導管内の水柱が引っ張られると、水の凝集力によって負の圧力(張力)が生まれ、水が蒸散流として長距離を移動する。道管内の負の圧力は-1 MPaにもなることが確認されている。このように植物体内の水移動を、水の凝集力と張力によって説明する考え方を「凝集力と張力の理論cohesion-tension theory」と呼ぶ。 半径40μm(10−6 m)の木部道管中を水が樹高100mの木の頂上まで輸送されるために必要な木部の張力は、重力を考慮すると約3 MPaと見積もられる。1㎠に30kgの過重に相当する。このような大きな張力が植物の木部に存在することは、水分子間の凝集力によって、道管内の水柱が分断されないで保たれるとする凝集力説に基づいて説明される。
木部に大きな張力が存在しうるか否かについては、これまでいろいろな議論があったが、これを証明する実験、あるいは木部内張力の直接測定から、実は、その存在自体が、現在では再確認されている。
水は植物の生育にとって不可欠で、欠乏すると植物は種々の影響を受ける。体内水分の不足は、 土壌水分が減少した時だけでなく、蒸散の多い条件下では、土壌水分が十分あっても起こる。しかも、体内水分欠乏の状態は、土壌水分の絶対量や蒸散条件だけで決まるわけではない。植物個体の生育過程などによって、現在おかれている土壌水分や大気環境が等しくても、個々の植物の生理的影響physiological effectsは著しく異なる。そのため、降雨が少なく土壌水分が著しく減少する地域だけでなく、降雨の多い湿潤な地域でも、植物は水分欠乏の状態になりうる。
生育期間中に水分条件が急激に変化すると、植物は脱水枯死する。湿潤土壌に生育していても、急激な高温乾燥条件下となれば、植物の器官、あるいは個体全体が短時間に水を失って枯死することがある。例えば、水稲においては出穂開花期にフェーンが起こると穂が水分を急激に失って穂全体が枯死しるが発生する。フェーンが到来すると短時間のうちに高温、低湿度、強風の高い蒸散を引きおこす条件となる。この時の白穂(しらほ;実らずに枯れて白くなった稲穂)は穂首(ほくび)の道管の水分通道機能が著しく低下することによって起こる。この水分通道機能の低下は気泡による道管の閉塞によって起こる。自穂はフェーンが昼よりも夜に起こった方が多く発生することが知られている。夜間のフェーンによる白穂の発生は、穂の蒸散が多くなるだけでなく、穂の蒸散が短時間に急激に増加することに関係しているかもしれない。
高温乾燥強風条件下における水稲の白穂は、穂首における水分通道抵抗の著しい増加によって起こる。この水分通道抵抗の増加は、高温乾燥強風条件下で、穂の蒸散速度が増加し、穂の水ポテンシャルが低下し、それが茎の水ポテンシャルの低下の引金となって起こる。白穂となった水稲の止葉から穂首までの茎の道管には、チロシスtylosesの形成や損傷が認められなかったことから、茎の水分通道抵抗の増加には、茎の水分通道抵抗に関係するもう1つの要因、すなわち、道管内に空気が入り込むキャビテーションcavitationが関係しているらしい。
Tylosesとは、「肥厚化」・「異常発達」の意味で、道管や道管に隣接する木部柔組織に形成される空胞状の派生物である。 根及び茎葉の外観に、全く健全な株が急に萎凋することがあり、原因は不明とされてきたが、顕微鏡観察の結果、本症発生株の茎基部や根部の90~100%の道管がチロシスによっ て閉塞されている特異的な現象が認められた。チロシスの形成機構は、複雑であるが、道管内の水分通道と関係があり、水分収支の生理的不均衡に起因すると推察される。高温乾燥な どの原因で機能の低下した根から水分供給量が減少し、一方、生育旺盛な茎葉部の吸引力が強いため、管内の圧力が低下し、周囲の活力に富んだ柔細胞が、道管の膜孔や薄膜部を貫いて管内に膨出してチロシスを形成し、それが加速度的に増殖し、全道管の閉塞が起り、水分通道が妨げられて萎凋に至ると推測されいる。
キュウリで問題となっている急激な萎凋と枯死、その収穫期間中のキュウリの急性萎凋は、収穫期の梅雨が数日続いた直後の暖かい梅雨晴に多く発生し、茎の道管がチロシスによって閉塞されることによって起こる。急性萎凋は、吸水と蒸散の不均衡による葉の水ポテンシャルの低下が引き金となり、茎の水の通道抵抗が著しく増加することによって発生し、それは茎の道管に形成さるチロシスによると考えられている。萎凋の認められない健全なキュウリの根の一部を切除すると、葉の水ポテンシャルの低下に伴って茎基部の水分通道抵抗が増加し、チロシスの形成も多く認められた。チロシスは蒸発散の盛んな環境で吸水が蒸散に追いつかず、葉や茎の水ポテンシャルが低下すると発生する。そして一度チロシスが形成されると水の通導抵抗の増加、茎葉の水ポテンシャルの低下、それによるチロシスの増加が正のフィードバック(ある変化が起こったとき、その作用をさらに強めるように働くこと)となって作用する。
目次へ
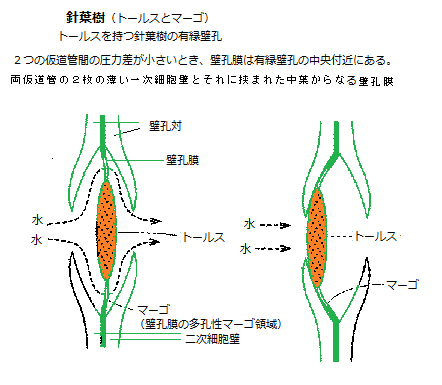 土壌水分が大きく低下し、植物が著しい水ストレス状態に置かれると、木部内に気泡が発生し、 (仮)道管内の水の繋がりが切れることがある。これをキャビテーション
cavitationと呼ぶ。キャビテーションが発生した (仮) 道管は、通水機能を失う。それにより、キャビテーションが多量に発生すれば、木部の水輸送機能が大きく低下し、水ストレスを助長する。気泡は、木部液に溶けていた気体がガス化して発生すると言うよりも、木部液のなくなった隣接する(仮)道管内の空気が、張力によって、壁孔から機能している
(仮) 道管内に引き込まれたり、(仮)道管壁の疎水性の割れ目から侵入したりするものと考えられている。
土壌水分が大きく低下し、植物が著しい水ストレス状態に置かれると、木部内に気泡が発生し、 (仮)道管内の水の繋がりが切れることがある。これをキャビテーション
cavitationと呼ぶ。キャビテーションが発生した (仮) 道管は、通水機能を失う。それにより、キャビテーションが多量に発生すれば、木部の水輸送機能が大きく低下し、水ストレスを助長する。気泡は、木部液に溶けていた気体がガス化して発生すると言うよりも、木部液のなくなった隣接する(仮)道管内の空気が、張力によって、壁孔から機能している
(仮) 道管内に引き込まれたり、(仮)道管壁の疎水性の割れ目から侵入したりするものと考えられている。被子植物の木部(道管)において、長い細胞が縦に並び、隔膜が消失して互いに繋がり、長い管になり、この管が根から茎や葉への水分の通路となる。道管細胞の一次壁の内側に二次壁が肥厚し、リグニンを沈着させ、そのリグニン沈着によって、細胞壁に疎水性と強度が与えられ、植物体の支持と長距離の水分通道が可能にる。
キャビテーションの発生した(仮)道管は、植物が水ストレスから解除され、根圧によって木部内に正の水圧が生じるまでは、再度、木部液で満たされることがないため、キャビテーションの補修はないと見られていた。しかし、根圧の値は小さ過ぎるため、キャビテーションの補修を説明しきれなかった。事実、キャビテーションの補修には、他のエネルギーや細胞の水輸送の特性の変化などが関与していた。晴天日の日中に発生したキャビテーションが、午後のまだ蒸散している条件でも補修されることが、凍結走査電子顕微鏡(凍結した生物試料を化学固定剤で固定し、試料の微細構造の変形・物質の移動・抽出を少なくする)の観察から明らかになった。
日本列島では、一年を通してみれば降雨量が、蒸発量を上回る湿潤状態が常である。それでも、夏の畑作物は、数年に一回の割合で旱魃となる。このため、作物の生育期間中に水環境が、湿潤から乾燥へと急激に変化することになる。盛夏の直前の梅雨期が、通常、夏作物の旺盛な栄養生長期でありながら、湿潤・ 低日照条件で茎葉は大きく繁茂するが、根系の発達が阻害され浅くなる。この直後に乾燥しやすい夏となれば、茎葉からの蒸散は多くなるが、浅い根であるため、水分は土壌の表層からしか吸収できない。その結果、土壌の深いところに水が多く残っていても、作物は大きな水ストレスを受けることになる。
キャビテーションは、土壌水分が大きく減少した条件で起こるだけでなく、比較的湿潤な土壌に生育していても起こることがいくつかの草本植物で認められている。しかも、深刻な影響を及ぼす著しいキャビテー ションが、湿潤土壌に生育している植物でも発生していた。台風の通過などに伴って発生す るフェー ンfoehn(山地を吹き越え、反対側の斜面を吹きおろす高温乾燥の強風)によって、湛水状態の水田で出穂期の水稲の穂が一夜にして白くなる現象が時々見られる。これは穂の水分が著しく減少することによるもので、白穂(しらほ;イネ科農作物の穂が水分の異常な欠乏により白色に枯れ上がること)と呼ばれている。
白穂の多く発生した水稲は、著しい減収となる。この白穂は穂首の道管に発生す多量のキャビテーションによって、水分通道抵抗が大きくなり穂への水供給が著 しく阻害されている。 白穂の発生には品種間差があり、発生しやすい品種は穂首の水分通道抵抗が大きくなりやすい。白穂は昼よりも夜のフェー ンによって発生することが、経験的に知られている。昼よりも夜に著しいキャビテーションが発生する理由が解明できれば、白穂発生の品種間差の生じる要因や耐性の解明、及び品種の育成につながると考えられる。
昼のフェーンに比較して、夜間にフェーンが発生すると、穂からの蒸散速度が急激に大きく増加し、木部の張力が急激に大きくなる。著しいキャビテーションの発生には、このような木部の張力の急激で大きな増加が関係しているようだ。
湿潤土壌に生育していても、晴天日の日中には水ストレスによる気孔の閉鎖が植物に起こる。キャビテーションが発生すれば木部における水輸送機能の低下となり、水ストレス はより高まる。このような状況であれば、植物キャビテーションが発生している可能性が高い。ただ、土壌水分が徐々に減少していく時は、植物はこれにかなりよく対応できる。水分が多く残っている土壌で生長を開始した植物は、土壌水分の減少に伴って根系を土壌深くまで、そして密に伸ばすとともに、茎葉の繁茂を抑制し、さらに茎葉からの蒸散が、根からの吸水を大きく上回る時には、気孔を閉じるなどして水分の消費を抑制するよう馴化する。 蒸散量は、土壌の有効水分量と根の密度や広がりと深さなどが関係する。土壌水分は土壌の表層から減少していくので、土壌水分の減少よりも根の生長が早く、根は早くから土壌の深くまで伸ばすため、土壌表層の水分の減少の影響を受けにくくなる。
生育初期の晴天日には、葉は萎凋しやすいが、生育の後期には、根は、既に水分が多く残る土壌深く達するため、萎凋は起こりにくい。生育期間中の降雨が著しく少なく、蒸散量の多い半乾燥地でも、潅水なしでも通常多くの植物は、地上部の生長は著しく抑制されるが、途中枯死することなく、子実をつけることができる。
しかし、水稲のある萎凋性突然変異系統では、土壌水分が十分にあっても蒸散が盛んであれば、キャビテーションが多発し、それが葉の著しい萎凋をひき起こす原因の一つとなっている。キャビテーションの発生の難易は、耐乾性と関連した植物種の地理的分布とも深く関わる。
地球環境の温暖化に伴う気象条件の変動の激甚化が、植物の蒸散速度の増加に影響し、植物のキャビテーション現象が、いろいろな諸条件とからみ、顕在化してくる危険が高まっている。
わが国の水稲栽培では、胚乳の澱粉蓄積の一時的不良によって起こる米の品質低下が、近年大きな問題となっている。この米の品質低下の原因 としては、登熟期の高気温があげ られているが、これに加えて、登熟期のフェーンも、品質低下をひき起こす要因の一つであることが、気象条件の解析から指摘されている。
米は、晴天率が非常に高く、日照時間が長いという環境により、光を十分に吸収した葉緑素が、光合成を行うことにより、デンプンを作る。晴天が続き、夜涼しい気候で、稲穂の穎果caryopsis(えいかkæ`riɑ'psis)が育つと、日中に作られた養分は安定的にうまみ成分となって実に蓄積され、美味しい玄米になる。昼と夜の「寒暖差」が、特に重要である。昼間暑い間に養分を作り、夜寒くなると、その養分を実に回す。
寒過ぎると植物は枯れてしまう。その危機を察知し、子孫を絶やさないために、植物は種子を充実させようとする。それで、昼暖かく、夜寒い地方の米が美味しくなる。最も重要なのは稲の出穂後、約30日間の登熟期間の気温が、「お米」の味を決定づける。出穂後30日間を目安に落水する。この期間、平均気温25~26℃が理想的な気温となる。近年、登熟期に、異常高温や夜のフェーンがしばしば起きており、これが原因で、品質や収量が落ちている。
水ストレスが急激に起こると、ゆるやかに起こる時よりも葉の水利用効率(蒸散速度に対する光合成速度の比)が低下することがある。葉内水分が徐々に低下する時は、葉全体にわたって均一に閉じる気孔が、葉が急激に水分を失う時には均一に閉じなくなるためである。ヒマワリでは、気孔閉鎖の不均一のレベルは、空気湿度の高い条件に生育すると大きくなる。また、空気湿度の高い条件に生育したヒマワリの葉は、水ポテンシャルの低下に対する閉鎖反応が鈍くなり、土壌水分が減少すると、枯死しやすくなる。
このように乾燥条件に対する植物のいろいろな反応を観察すると、植物は徐々に乾燥する条件では、極めて粘り強い馴化を示すが、これに比べて湿潤から短期間に急激に乾燥する条件下では、それほど厳しい乾燥条件でなくても著しく大きな影響を受ける。これには、すでに述べ たように湿潤条件に生育することによって、乾燥耐性に関与する最も重要な性質の一つである根系の適応の発達程度が劣ることがあげられる。さらに、根だけでなく茎の導管の閉塞や気孔反応の鈍さや不均一性から推察されるように、湿潤から乾燥へと急激に変化する水環境に十分に対応できる機構が、茎や葉に備っていないことも関係しているようである。
葉や茎が形成される過程で、水環境は道管の閉塞や気孔反応に関与する性質にも大きな影響を及ぼしているかも知れない。いずれにしても、まだ明らかでない点が多い。種間差とともに品種間差が認められる場合も多いので、 耐性を示す種や品種の性質を検討することによっ て、発生機構や耐性に関わる性質を明らかにできると考えられている。乾燥地域だけでなく湿潤地域で起こる水ストレスの実態を明らかにし、水ストレスの耐性機構を解明していくことが、植物の水環境に対する反応に関する理解を深め、水ストレス耐性植物の育成に繋げることも重要であると言える。
キャビテーションの発生が、陸上植物の進化や種分化の過程で重要な影響を与えてきたことを示唆している。
目次へ