| Top 車山高原 車山高原お知らせ 車山ブログ
|
 |
|||||||||
| 車山のフキノトウ 3月25日撮影 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生
花粉の形成と構造 雌ずい群
| 目次 |
| 1)被子植物の受精過程 |
| 2)乳頭細胞のフット構造 |
| 3)極性が生み出す様々な発生過程 |
| 4)栄養核と精細胞 |
| 5)エクスパンシンexpansin類と酸成長 |
| 6)花粉管と胚珠 |
| 7)受容体キナーゼは、先端成長の主要な制御因子である |
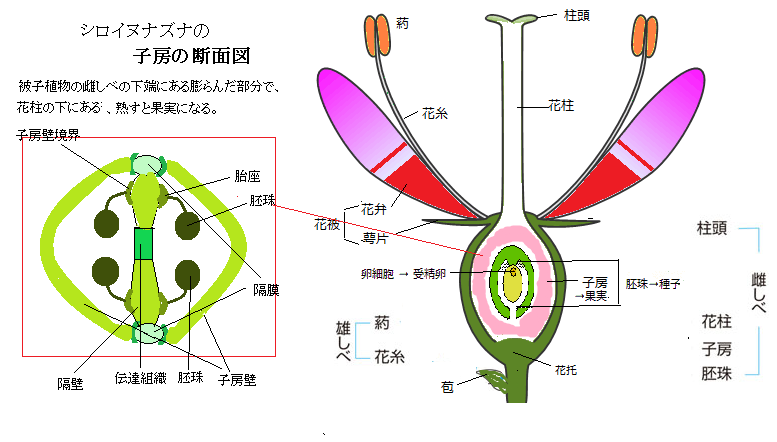 胚珠ovule(ɔ'vjuːl)は種子植物が備える特有の構造で、被子植物の場合は、雌ずい群(心皮carpel; kɑ'ːrpəl)を構成する雌性生殖器官であり、子房ovary(óʊvəri)の中核組織である。胚珠は、大胞子、即ち雌性配偶体(卵細胞)を形成する。
胚珠ovule(ɔ'vjuːl)は種子植物が備える特有の構造で、被子植物の場合は、雌ずい群(心皮carpel; kɑ'ːrpəl)を構成する雌性生殖器官であり、子房ovary(óʊvəri)の中核組織である。胚珠は、大胞子、即ち雌性配偶体(卵細胞)を形成する。被子植物angiosperms(angio;果皮、sperm;精液・精子)の受粉は、花の雄性器官である雄ずい(雄しべ)の葯の内部で作られる花粉が、花の雌性器官である雌ずい(雌しべ)の柱頭に運ばれることである。
シロイヌナズナやイネなどの植物種では、花粉が同じ個体の雄ずいの柱頭に受粉する自家受粉(自殖)を行うが、花粉が他の個体でなければ受粉しない他家受粉(他殖)の種も多い。多くの植物種では、自家受粉と他家受粉の両方が可能であるが、自家受粉を回避し他家受粉を優先する植物種も多い。更には、自家受粉を不可能にまでしている例も稀ではない。
他家受粉であれば、花粉が柱頭に達するまでに、非常に長い距離を風任せて移動する。その使命を達するために花粉は、大量に生産され、風ばかりか鳥・虫・哺乳類を介して受粉される。雄性配偶子自体は移動しないが、この散布により、基部陸上植物であるシダ植物やコケ植物の精子は、泳げる範囲よりも遠くまで運ばれる。
シダ植物やコケ植物では、胞子体世代で胞子を作り、その胞子が散布され配偶体へと成熟する。その配偶体では、雄では造精器が、雌では造卵器が分化する。
シダ植物の配偶体は、その胞子が発芽して形成される雌雄同株の前葉体prothallium(prouθǽliəm)で、それは有性世代すなわち配偶体であるので、造精器および造卵器が生じる。多くのシダ類の前葉体は、薄い1~2層の細胞から成る心臓形で、横径数 mm~10mmの葉状である。その中の造精器で運動性の高い精子が、片方の造卵器から卵細胞が、それぞれ作られる。
コケ植物では、葉のような形をした配偶体世代が、生活環の大半を占めるが、種によって造精器と造卵器は同じ個体(雌雄同株)にあったり、別の個体(雌雄異株)それぞれに生じたりする。
運動性の精子は、その後造卵器の内部に進入し、卵と受精することにより2Nの核相を持つ受精卵となる。受精卵は、そのまま配偶体組織に囲まれて胚へと生育する。その後、種子を形成することなく成長して2Nの胞子体になる。
被子植物の受粉の成否は、周辺環境に左右され、温度・時期、そして重要なのが花粉を受け入れる柱頭の受容性などの要因に影響される。トマトなどのように、高温により障害を受ける植物種もあるが、多くの花粉は、長期間腐食することもなく、着地した大地の高温と乾燥に耐性を備えている。
被子植物の雌性配偶体は、子房組織により厳重に保護されている。そのため精細胞が、卵細胞に到達するには、柱頭から胚珠へ向かう花粉管によって運ばれる必要がある。
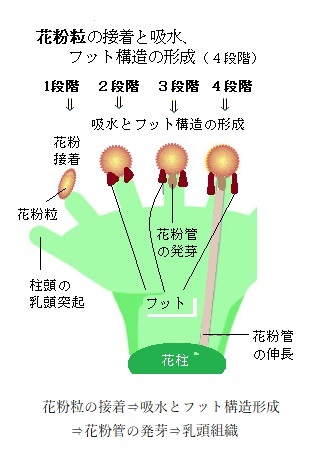 例えば、シロイヌナズナの雌ずいでは、他の被子植物と同じように、6段階の過程をたどる。
例えば、シロイヌナズナの雌ずいでは、他の被子植物と同じように、6段階の過程をたどる。花粉は、雌ずいの表皮(乳頭細胞)に接着し、吸水した後に発芽する。雌ずいには様々な花粉が運ばれて来るが、交配相手として適切な花粉のみが、この過程で選択される。被子植物の生殖は、非常に選択的である。アブラナ科植物の自家不和合性では、花粉は雌ずい先端の乳頭細胞で自他識別が行われる。乳頭細胞は、多種多様な花粉粒を識別して、類縁性のない植物種の花粉などは、ここで拒絶され、和合性のある花粉粒だけが、柱頭の乳頭細胞に物理的に接着できる。
雌ずい上で吸水・発芽した花粉管は、胚珠まで正確に誘導されていき、そこで受精が行われる。花粉と雌ずいの細胞との間で、実に様々なコミュニケーションがなされている。これは、花粉のタンパク質や脂質と、柱頭表面のタンパク質間の物理的、および化学的な相互作用が働いているからで、花粉粒は、異なる科の植物の柱頭では、その相互作用が生じないため接着ができない。
植物の花には、湿性型と乾性型、いずれかの柱頭がある。湿性型の柱頭の表面細胞は、タンパク質・脂質・多糖類からなる粘着性の混合物を分泌する。一方、アブラナ科のような乾性型の柱頭の表面細胞は、細胞壁・クチクラ・タンパク質の外被で覆われている。
花粉粒は、湿性型の柱頭上では、自然に吸水するが、乾性型の柱頭上での吸水は、極めて厳密に制御されている。柱頭に付着した後、花粉表面から脂質とタンパク質が柱頭に流出して、乳頭細胞由来の物質と混ざり合って、フットと呼ばれる構造体を形成する。この構造体によって、花粉粒は乳頭細胞の先端に確実に接着できるようになる。
このフット構造を形成する過程では、脂質が再構成され、柱頭から花粉粒へ水分やイオンを運ぶ毛細管現象が起きるようだ。この仕組みにより、花粉粒が乾性型の柱頭上で吸水するという矛盾する過程が生じる。
目次へ
 花粉が吸水される際の脂質の役割を示す例として、長鎖脂質の代謝に異常のあるシロイヌナズナの変異体では、花粉外被のない花粉が形成される。そのため、これらの花粉粒は、柱頭では吸水できない。この異常は、花を高湿度に置くことや、外部から柱頭へ脂質を与えることにより回復し、変異体の花粉粒でも吸水すれば、花粉管を伸長することが可能になる。
花粉が吸水される際の脂質の役割を示す例として、長鎖脂質の代謝に異常のあるシロイヌナズナの変異体では、花粉外被のない花粉が形成される。そのため、これらの花粉粒は、柱頭では吸水できない。この異常は、花を高湿度に置くことや、外部から柱頭へ脂質を与えることにより回復し、変異体の花粉粒でも吸水すれば、花粉管を伸長することが可能になる。乳頭組織からフット構造への水分の移動のメカニズムは、いまだ明らかにされていないが、原理的には、細胞膜にある水チャンネル(アクアポリンaquaporin; aqua=水、porin=孔の意)を通して、水分が与えられるか、小胞が細胞膜に融合して、エキソサイトーシスexocytosis(開口分泌とも言う。小胞内に隔離した固体や溶液を、その小胞と細胞膜を融合させることによって、細胞内物質を放出する作用)により内容を分泌するからだと考えられる。
アクアポリンは、細胞膜を横切る水選択性チャンネルを構成する膜内在性タンパク質で、細胞膜を横切る水の移動を促進する。アクアポリンは、膜を横切る水輸送の速度を変えられるが、水輸送の方向や駆動力は変えられない。しかし、細胞内のpHやCa2+のレベルなどを、生理的パラメーターparameter(媒介変数)にして可逆的に開閉する。それにより、植物は細胞膜の水透過性を制御する。
エキソサイトーシス分泌による水分移動のメカニズムを証明するのが、ゴジル小胞のエキソサイトーシスに必要な遺伝子の変異体では、花粉粒が雌ずい上で吸水できないことがあげられる。
植物は、発生の初期にはしなやかな細胞壁を作り、その柔軟さにより、細胞壁を伸ばして、新たな構造素材を組み入れるようにする。一般的に細胞壁は、長いセルロース微繊維が、水和した非セルロース性の多糖類と、少量の非酵素タンパク質に埋め込まれてできる薄い層が、何層か重なった構造からできている。つまり、一次細胞壁は、セルロース微繊維が、ペクチンとヘミセルロースhemicelluloseのマトリック(膜内に含まれるコロイド状の水相)に埋め込まれた構造がベースとなる(ペクチンとヘミセルロースは、マトリック多糖類とも呼ばれる)。セルロース微繊維は、細胞の表面で合成され、部分的にヘミセルロースに覆われ、それにより微繊維が互いに離れた配置となる場合がある。ペクチン類は、一体化したマトリックとなり、微繊維間の間隔や間隙率を制御している。この構造により、成長中の細胞壁が、伸びる柔軟性と強度を、同時に併せ持つことを可能にした。
一次細胞壁に共通するのが、成長中に作られること、セルロース微繊維間には高度に水和したマトリックが満たしていること、少なくとも細胞成長中には伸展ができることなどである。一次細胞壁には、かなりの水の量が含まれるが、その約75%は、マトリックに局在している。それが一次細胞壁の物理的特性を主に決定づけている。例えば、細胞壁は、水含量が乏しくなると伸展しなくなる。これにより、水不足時に、植物の成長が抑制される。やがて細胞壁の脱水は、リグニン化lignificationの過程をたどるようになる。この点で二次細胞壁とは大きく異なる。二次細胞壁は高密度に固められ、細胞が拡大しないように構造的に補強する働きを担う。
目次へ
マトリック多糖類は、ゴルジ装置内の膜結合型転移酵素により合成され、分泌小胞のエキソサイトーシスを介して、細胞壁まで移送する。つまり、分泌小胞が細胞膜と融合して分泌し、細胞表面に多糖類を蓄積するのである。
シロイヌナズナの花の柱頭上で起こる花粉の接着と吸水は、
花粉粒の接着⇒吸水とフット構造形成⇒花粉管の発芽⇒乳頭組織を使って花柱に向かう伸長
という4段階である。
花粉粒は、吸水中に生理的に活性化される。栄養細胞へのカルシウムイオンの流入は、細胞骨格の再構成を惹き起こし、細胞内に生理的及び構造的な極性polarity(poulǽrəti)を生じさせる。これにより、生物体の細胞・組織が、ある軸に沿って、形態的・生理的な差異を示すことになる。
種子植物の胚発生は、たった1個の接合子(有精卵)から、複雑な個体を成熟した種子の中に作り出す。胚発生により、多くの極性が生じ、様々な発生過程を経て、植物の基本構造が形成される。例えば、真正双子葉植物のシロイヌナズナの胚発生と、単子葉植物のイネや他の多くの植物種の胚発生と比較すると、胚の形とサイズ、細胞の数、分裂パターンに明らかな違いがある。これらの違いがあるものの、すべての種子植物には、一般化できる大きな共通点が見られる。それらの共通点の最も基本的な部分に、深く「極性」が関与している。
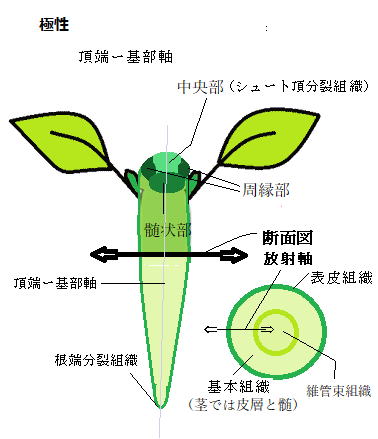 単一細胞の接合子(有精卵)として発生を開始した胚は、2つの軸、頂端―基部軸apical-basalaxis(頂点のāpikəl;基部béisəl;ǽksis軸)、と放射軸radial axisの両軸に沿った発生を通して、極性の度合いを急速に高めていく。頂端―基部軸は、胚のシュート頂と幼根の頂端を結ぶ軸であり、放射軸は頂端―基部軸と垂直に交差し、胚の中心から維管束組織・基本組織(茎では皮層と髄、根では皮層・内鞘=側根分裂組織)・表皮組織を横切って表層に向かう。頂端―基部の極性は、胚発生の初期に確立する。
単一細胞の接合子(有精卵)として発生を開始した胚は、2つの軸、頂端―基部軸apical-basalaxis(頂点のāpikəl;基部béisəl;ǽksis軸)、と放射軸radial axisの両軸に沿った発生を通して、極性の度合いを急速に高めていく。頂端―基部軸は、胚のシュート頂と幼根の頂端を結ぶ軸であり、放射軸は頂端―基部軸と垂直に交差し、胚の中心から維管束組織・基本組織(茎では皮層と髄、根では皮層・内鞘=側根分裂組織)・表皮組織を横切って表層に向かう。頂端―基部の極性は、胚発生の初期に確立する。カルシウムイオンの供給源は不明だが、花粉粒の細胞質または乳頭細胞の細胞壁と見られている。シロイヌナズナの花粉粒の生きている細胞を可視化する「ライブイメージング法live cell imaging methods」により、細胞質内の遊離カルシウムイオンの濃度は、吸水後、直ぐに花粉管の発芽予定部位で上昇し、花粉管が発芽するまで、その上昇状態が保たれていることが観察されている。
Ca2+は、植物のみならず他の真核生物においても、最も普遍的なセカンドメッセンジャーである。セカンドメッセンジャーとは、シグナル受容後、比較的高いレベルで、迅速に生産 あるいは移動する低分子化合物やイオンで、標的となるシグナル伝達タンパク質の活性を変化させるものであり、シグナルを強めたり増幅させたりするもう1つの方法でもある。
こうして、Ca2+は、セカンドメッセンジャーとして、シグナルを増幅し、標的シグナル伝達タンパク質の活性を調節して、生理反応を惹き起こす。
すべての真核生物で、最も普遍的なセカンドメッセンジャーは、カルシウムイオン、即ち二価陽イオンのCa2+である。植物では、共生相互作用、防御反応、様々なホルモンや非生物ストレスに対する応答を含む非常に多くのシグナル伝達経路に関与している。サイトゾルのCa2+濃度は、Ca2+透過性のイオンチャンネルが開き、貯蔵Ca2+からサイトゾルなへの受動的なCa2+の流入が可能になると急激に増加する。
イオンチャンネルは、ゲート開閉する。つまり、チャンネル孔が、膜電位の変化・膜張力・翻訳語修飾・リガンドとの結合により開閉を行う。植物のCa2+透過性チャンネルのいくつかのファミリーが、既に同定されている。
細胞質カルシウム濃度の上昇は、カルシウムセンサータンパク質であるカルモジュリン(CaM)・Ca2+依存的タンパク質キナーゼ(CDPK)・カルシニューリン-B様タンパク質/CBL相互作用性タンパク質キナーゼ(CBL/CIPK)を活性化させる。これらのタンパク質は、細胞内の様々な場所に局在している。
水分とカルシウムイオンのほかにも、柱頭からは、花粉管の発芽を促進する様々な物質が分泌されているようだ。
特定のタンパク質が出合う数千もの分子のうち、結合できるのは1個ないし数個に過ぎない。タンパク質が結合する物質は、イオン・有機小分子・巨大分子を問わず、そのタンパク質のリガンド(ligand)と呼ぶ。Ligandは、ラテン語で結合を意味するligareを語源とする。
タンパク質が、選択的にしかも親和性(ある物質が他の物質と容易に結合する性質や傾向)でリガンドと結合するのは、水素結合・静電引力・ファンデルワールス引力van der Waals forceなど一連の弱い非共有結合に加えて、疎水力が働くからである。疎水性の尾部を構成する分子や疎水基をもつ分子などは、細胞内の水溶液中にあると、水との接触を避けるために水と接する面積を減らそうと、互いに会合する疎水力が働く。C-H結合を数多く持つ炭化水素は、非極性結合であるため疎水性である。そのため水分子を引き付けず、溶液中に出ていくことはない。
目次へ
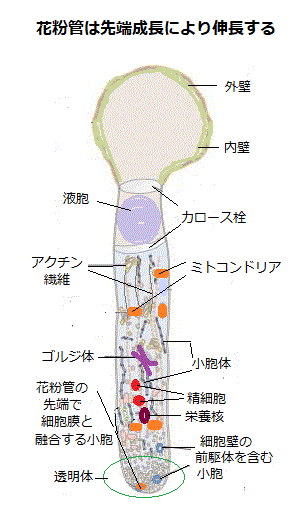 アクチン繊維と分泌小胞は、共に発芽口直下に蓄積し、栄養核が精細胞より先に発芽中の花粉管に入ることを可能な位置に移動させる。アクチン繊維(F-アクチン)は、球状タンパク質であるG-アクチンを単量体とする重合体である。その高分子集合体は、直径7nm(10−9m)の凝縮された1本の鎖、らせん状構造をもつ。アクチン繊維は、酵母のみならず高等動植物の全ての真核細胞内に遊離して存在し、真核細胞にとって非常に重要な働きをしている。実際に、細胞内で特徴的なパターンをもって、細胞内に張り巡らされたネットワークを形成し、動的な平衡状態を保っている。
アクチン繊維と分泌小胞は、共に発芽口直下に蓄積し、栄養核が精細胞より先に発芽中の花粉管に入ることを可能な位置に移動させる。アクチン繊維(F-アクチン)は、球状タンパク質であるG-アクチンを単量体とする重合体である。その高分子集合体は、直径7nm(10−9m)の凝縮された1本の鎖、らせん状構造をもつ。アクチン繊維は、酵母のみならず高等動植物の全ての真核細胞内に遊離して存在し、真核細胞にとって非常に重要な働きをしている。実際に、細胞内で特徴的なパターンをもって、細胞内に張り巡らされたネットワークを形成し、動的な平衡状態を保っている。アクチンの単量体には、1分子のヌクレオチドが結合できるため、ATPまたはADPが結合して、アクチン繊維は極性を持つようになり、両端の性質が異なることになる。極性は両端の伸長速度の違いによって見分けられる。伸長速度の高い方がプラス端plus end、低い方がマイナス端minus endである。アクチン繊維では、単量体のアクチン分子自体がアクチン繊維の極性を生じさせている。分子中のATP/ADP結合間隔が露出している方がマイナス端側を、その反対側がプラス端側を向いている。
アクチン繊維の形成過程は、最初はATPを結合したG-アクチンが集合してF-アクチンからなる小さな重合核を形成するのが始まりである。その重合核が安定して伸長するためには、ある程度の大きさが必要であるが、時間がかかる。
球状のG-アクチンが、何千もの構成単位となって集まってタンパク質の糸を作り、細胞全体を横断していることもある。これが.アクチン繊維だ。
重合核が十分な大きさになると、プラス端の重合がより速まり、マイナス端の重合はゆっくり進む。
やがて、プラス端側での重合とマイナス端側での脱重合が始まり、その速度が等しくなる定常状態に達する。G-アクチンに結合したATPは、プラス端に付加された後、次第にADPへと加水分解される。マイナス端側からG-アクチンが解離される際には、その脱重合されたG-アクチンは、ADP結合型となっている。
細胞質は絶え間なく動いている。ミトコンドリアや膜で囲まれた小さな細胞小器官や小胞は少しずつ動き、時には止まって、また動き出す。この跳躍運動は、熱運動による連続的な細かい運動よりも、持続的で方向性がある。
その植物細胞での跳躍運動は、アクチン繊維に沿って起こるが、その動きの原動力はモータータンパクmotor proteinである。アクチン繊維を含む、これらのタンパク質は、ATPの加水分解を何度も繰り返して得たエネルギーを使って、アクチン繊維上を決まった方向に着実に動いている。 モータータンパクには、他の細胞成分もくっつくので、それを積み荷にしてアクチン繊維に沿って輸送しているようだ。
アクチン繊維の重合核形成因子であるArp2/3複合体actin-related protein 2-3 complexと呼ばれるタンパク質が、重合核形成に関わる。前側に向かってアクチン繊維の重合を惹き起こし、その結果、細胞膜が前側に押し出される。Arp2/3複合体は、既存のアクチン繊維の分岐から、新しいアクチン繊維を特定の角度で伸長させることができる。その一方後部では、アクチン繊維とミオシン繊維が相互作用し、収縮力を発揮させる。これにより細胞後部が特定方向に引き込まれて行く。
アクチンとモータータンパク質であるミオシンが、細胞質内で協調して働いて運動をすることから、アクトミオシンactomyosin細胞骨格系と呼ぶことが多い。植物の細胞小器官は、運動しているのが常態である。個々の細胞小器官の運動は、粘性のある細胞質中でありながら、周囲の細胞質や細胞小器官を巻き込んで連動することにより、大きな流れとなる原形質流動の原動力となっている。しかも、大きな細胞ほど細胞小器官の流動速度が速い。その一方では、細胞小器官が互いに逆方向へ移動する「制御された方向性を持った細胞小器官運動directed organelle movement」があり、更には、方向性が定まった細胞小器官運動でありながら、他の細胞小器官や細胞骨格・細胞膜に繋ぎとめられる「細胞小器官の繋留organelle tethering」とも協調している。
モータータンパク質(分子モーターmolecular motor)は、方向性を持つ細胞小器官運動と繋留tetheringの両方に関与する。植物には、ミオシンとキネシンの2種類の分子モーターがある。ミオシンは、アクチン繊維と可逆的に結合する。植物にあるミオシンは、細胞小器官の繋留に主に関与するものと、栄養成長期の非分裂細胞で細胞小器官運動を担うものと、2つのファミリーがある。花粉管の発芽口直下で、アクチン繊維が働く意味合いが、これから推測される。
発芽後、先端成長tip growthを開始する花粉管は、乳頭細胞のクチクラ層の隙間を通り抜けて、乳頭細胞の細胞壁に進入する。ある種の植物では、花粉管の伸長速度は極めて高く、根毛の先端成長が毎秒10~40nm(10−9m)であるのに対して、生体内での花粉管伸長は、毎秒5µm(10-6 m)を超えるものもある。例えば、トウモロコシにひげは、心皮の花柱であるから、その花粉管の長さは、40cmにも達する。その伸長速度は、当然、速くなる。
伸長中の花粉管は、大きな液胞とカロースcallose栓(グルコースを重合した多糖、1,3-β-グルカン)によって仕切り後部を遮断することで、細胞質と2つの精核、1つの栄養核を先端部の伸長領域に限局させる。花粉管の先端部は、透明体clear zoneと呼ばれが、実際には透明ではなく、多数の小さな分泌小胞・ゴルジ体・アクチンを含む。ただ、核・小胞体・ミトコンドリアのような大きな細胞器官は含まれない。透明体の後方では、中心軸の両側で、原形質流動の循環が見られるが、透明体内部では見られないため、透明体形成の分子基盤は、原形質流動を担うアクチンケーブルactin cable(アクチン繊維の束)の分解と再構成に関わるもののようだ。
シロイヌナズナ雌性配偶子の1つである中央細胞に受精するのを解析すると、アクチンケーブルが細胞膜から核に向かって活発に動いており、精核もそれに沿って移動する様子が観察できる。アクチン繊維の機能を欠損させると、中央細胞のみならず卵細胞にも精核が移動しなくなる。
透明体の細胞質には、小さな分泌小胞に満ちており、これらの小胞は、花粉管の細胞壁の材料や新しい細胞膜を伸長端に供給しているようだ。
花粉管などが先端成長する際の極性制御は、伸長端のイオン勾配が関係しているようだ。伸長中の花粉管先端は、カルシウムイオンとpHが局所的に勾配を形成することで極性化する。細胞質内のカルシウムイオン濃度は、花粉管の最先端部で高く(モル濃度3~10µM;マイクロモーラー10−6 M)、先端から20µm以内では、基底レベル(モル濃度0.2~0.3µM)にまで下がる。
細胞質のpHは、透明帯の最先端部では、pH6.8とわずかに酸性であり、透明帯の基部ではpH7.5とアルカリ性である。
透明帯のカルシウムイオンの濃度と細胞質のpHは、花粉管の伸長速度の振動と相関して周期的に振動することから、相互関連が推測される。カルシウムイオン濃度とpHによる電気的・化学的変化は、花粉管の極性維持に必要な細胞内の情報伝達・細胞骨格の動態・膜輸送・エキソサイトーシスに関わることが分かっている。
サイトゾルや細胞壁のpHの変化は、ホルモンやストレス応答のセカンドメッセンジャーとして働いている。休止細胞では、サイトゾルのpHは一定に保たれて、通常pH7.5程度であるが、細胞壁は酸化されpH5.5か、それ以下である。
細胞外のpHは、内的あるいは環境シグナルに迅速に反応して変化する。一方、細胞内pHは、細胞の緩衝能力によりゆっくりと変化する。例えば、成長中の胚軸(発芽した苗の茎)では、植物ホルモンのオ―キシンが、細胞膜のH+-ATPase(プロトンATPアーゼ)をC末端におけるリン酸化により活性化する。これにより細胞壁がより酸性化し、タンパク質群であるエクスパンシンexpansin類などの細胞壁をゆるめる酵素を活性化することで、酸誘導性成長と細胞壁の応力緩和stress relaxationにより細胞伸長が促進されるようだ。
植物細胞が形を変えるには、細胞壁の伸展の方向と速度を制御する必要がある。この過程は、セルロース微繊維を特定の向きに揃えて配置して、細胞伸長の方向性を決め、微繊維間の接着を選択的にゆるめることで制御する。この生化学的なゆるみが、セルロース繊維とそれをとりまくマトリック多糖類(植物細胞壁のコロイド状の水相を構成する、セルロース微繊維以外の多糖類で、一次細胞壁では、ペクチンとヘミセルロースおよび糖タンパク質からなる)の移動や滑りを可能にして、それにより、細胞壁の表面積が増す。それと同時に細胞壁のゆるみは、細胞壁にかかる物理的応力を低下させる。
細胞壁の応力緩和が成長中の植物細胞の膨圧と水ポテンシャルを低下させ、これにより細胞が吸水して膨らむことを可能にする。応力緩和が生じなければ、細胞壁が合成されても伸展することはできない。その吸水された水は、液胞へ入る。細胞拡大が進むにつれ、液胞が占める細胞内の体積比率は増加し続ける。
成長中の細胞壁に共通する特徴の1つに、中性pHより酸性pH下の方が、より速い速度で伸展することがある。この現象を酸成長acid growthと言う。
酸に誘導された成長は、根毛分化の開始の際にも見られ、根の表皮細胞の特定部位で隆起が生じると、局部的に細胞壁のpHの値が4.5に低下する。このpH依存性の細胞壁伸展機構は、進化的にすべての陸上植物が保持している過程で、様々な成長過程にも関与している。
このような酸誘導性クリープcreep(時間依存性の不可逆的な伸展過程)は、成熟した細胞壁では見られない、伸長中の細胞壁の特徴である。
目次へ
タンパク質が酸成長には必要であるという考えは、細胞壁の再構成実験により確認された。この実験では、熱処理で失活した細胞壁に、成長中の細胞壁から抽出したタンパク質を加えると、酸性成長が殆ど回復することが実証されている。この活性成分は、エクスパンシンと呼ばれるタンパク質群である。
酸成長仮説acid growth hypothesis(hɑɪpάθəsɪs)によれば、オーキシンは水素イオンhydrogen ionを介して細胞壁を緩めている。その水素イオンの源は、細胞膜H+-ATPase(プロトンATPアーゼ)であり、その活性はオーキシンに応答して上昇する。オーキシンが細胞壁へプロトンprotonを放出を促進するための時間は、10~15分間かかり、これは成長速度上昇のタイミングと一致している。エクスパンシンは、pHが酸性の時に活性化し、細胞壁の構成要素である多糖類間の水素結合を弱め、細胞壁を緩める。
オーキシンが細胞壁へのプロトン放出を促進する時間は、そのエクスパンシン類による、pH依存型の細胞壁の伸長とは逆に、オーキシンが、根の細胞伸長を阻害する場合、オーキシンは細胞壁の迅速なアルカリ化を促す。前記の過程は、Ca2+(カルシウムイオン)にも依存している。同様なCa2+依存的なpHの変化は、植物の多くの環境ストレス応答で観察されている。
エクスパンシン類は、大きな遺伝子スーパーファミリーに属する。それはα-エクスパンシン(EXPA)とβ-エクスパンシン(EXPB)の2大ファミリーと、さらに2つの小さいファミリーに分けられる。
EXPAは、細胞壁のセルロース微繊維同士の接着点を緩めるため、真正双子葉類の細胞壁では、より活性が高い。EXPBは、細胞壁多糖類間の非共有結合による接着を緩めるため、イネ科植物の細胞壁に対して、より活性が高い。
 ペクチンとヘミセルロースは、マトリックス多糖類と呼ばれ、植物細胞壁は、足場となるセルロース微繊維と、それを埋める高分子性のマトリックスからなる。マトリックスは、植物種や細胞型、細胞壁内の部位により異なる。
ペクチンとヘミセルロースは、マトリックス多糖類と呼ばれ、植物細胞壁は、足場となるセルロース微繊維と、それを埋める高分子性のマトリックスからなる。マトリックスは、植物種や細胞型、細胞壁内の部位により異なる。真正双子葉植物の典型的な一次細胞壁は、ペクチンを多く含み、セルロースやヘミセルロースは少ない。二次細胞壁では、セルロースやヘミセルロースを多く含み、ヘミセルロースの分子種は、一次細胞壁のそれとは異なる。また二次細胞壁には、芳香族化合物の重合体であるリグニンを含み、その量は細胞壁の種類ごとで異なる。
原子核の陽子の数に比べて最外殻電子の数が少ないために、正に荷電している陽イオンをカチオンKationと呼ぶ。植物細胞に吸収されるカチオンは、有機化合物と非共有結合で、複合体を形成し、金属結合をする。植物は多量要素であるカルシウムイオン・カリウム・マグネシウムや、微量要素である鉄・銅・マンガン・コバルト・ナトリウム・亜鉛のイオンを、この有機化合物を介して非共有結合で同化する。
カチオンと炭素化合物の間で形成される非共有結合には、配位結合coordinate bondと静電結合electrostatic bondの2種類がある。この2種類の結合により、植物が必要な様々な栄養素を同化できるようになる。特に、根による鉄吸収と、その同化にとって重要である。
配位結合は、一方の原子から2個の最外殻電子を供出する場合をいう。配位複合体の形成では、炭素化合物中の数個の酸素原子または窒素原子が非共有電子を供与して、カチオンとの結合を生じさせる。これにより、カチオン上の正電荷は中和される。カルシウムイオンの大部分は、このやり方で細胞壁に結合していると考えられている。
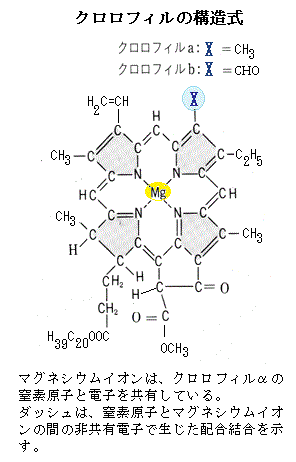
配位結合は、主として多価カチオンと炭素化合物の間で生じる。例えば、2個の最外殻電子を持つマグネシウムイオンとクロロフィルαの複合体では、マグネシウムイオンは、クロロフィルαの窒素原子の電子と共有結合している。他方では、2つの電子と非共有電子として配合結合している。
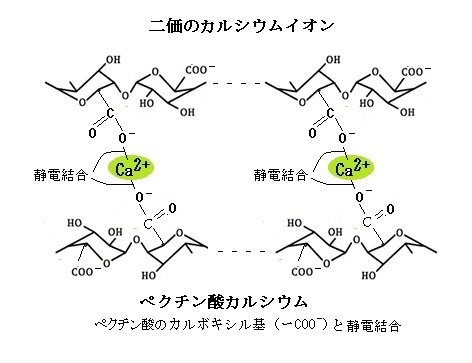 配位結合複合体として同化される栄養素には、銅・亜鉛・鉄・マグネシウムイオンなどがある。カルシウムイオンも、細胞壁のポリガラクツロン酸残基の水酸基(−OH)の酸素を共有する配位複合体を形成している。その結果、カチオン上の正電荷は中和される。カルシウムイオンの大部分は、これにより細胞壁に結合している。
配位結合複合体として同化される栄養素には、銅・亜鉛・鉄・マグネシウムイオンなどがある。カルシウムイオンも、細胞壁のポリガラクツロン酸残基の水酸基(−OH)の酸素を共有する配位複合体を形成している。その結果、カチオン上の正電荷は中和される。カルシウムイオンの大部分は、これにより細胞壁に結合している。原子番号 20、原子量 40.08 の金属元素(単体で光沢や延性があって、電気や熱をよく伝導する元素)であるCaは、最外殻電子を2つもつので、他の原子と反応しやすい2価のカルシウム陽イオン(Ca2+)になりやすい。生体内では骨などの主要成分として骨格形成に必要なほか、筋肉収縮や細胞内の信号伝達、補酵素など重要な働きをする。2価のカルシウム陽イオンは、ベクチン酸のカルボキシル基(-COO-)の酸素電子の負電荷を誘引して静電結合を形成する。静電結合は、配位結合とは異なり、静電結合のカチオンは、正電荷を保持したままである。このカルシウムのクロスリングcross ringは、細胞壁の構造を形成する役割を果たす。
カリウムイオンK+のような一価のカチオンは、多くの有機化合物のカルボキシル基と静電結合を形成している。特に、浸透圧調節や酵素の活性化に関わる殆どのカリウムイオンは、サイトゾルや液胞中に遊離している。
カルシウムイオンのような二価のカチオンは、ペクチン酸やポリガラクツロン酸のカルボキシル基と静電結合を形成する。カルシウムイオンの大部分は、これにより細胞壁と結合するようになる。
通常、マグネシウムイオンやカルシウムイオンは、アミノ酸やリン脂質、その他の負に荷電した分子と、配位結合や静電結合をして同化される。
陸上植物は独立栄養を営み、土壌や大気から獲得した無機態の栄養素から、すべての有機態の分子を合成することができる。多くの無機栄養素を有機化する過程では、根による土壌からの吸収と、成長と分化に必要な有機化合物への取り込みが含まれている。
色素・酵素コファクター・脂質・核酸・アミノ酸など有機化合物形成のための無機栄養分の取り込みを、栄養素の同化nutrient assimilationと呼んでいる。
目次へ
花粉管は、雌ずいに進入すると、まず雌ずい先端の乳頭細胞の細胞壁の内部に、更に伝達経路transmitting tract(柱頭から子房の珠孔まで花粉管が成長する径路)を通って細胞間隔を伸長する。
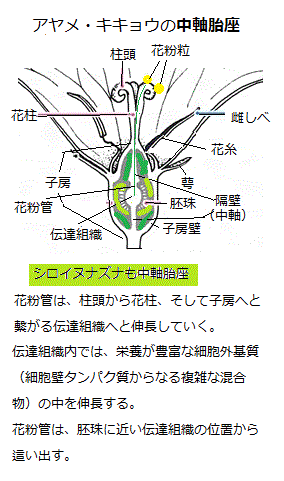 子房内にあって胚珠がつく位置を胎座placentaと呼ぶ、その子房内の様々な特殊組織から、胚珠原基は生じる。
子房内にあって胚珠がつく位置を胎座placentaと呼ぶ、その子房内の様々な特殊組織から、胚珠原基は生じる。胎座組織の位置は、植物種によって異なり、
面生胎座laminar placentation・辺縁胎座marginal placentation・側膜胎座parietal placentation・中軸胎座axial placentation・基底胎座basal placentation・独立中央胎座free central placentation
などがある。
その子房内での胎座の位置により、果実内の種子の配置が定まる。
心皮とのかかわりでみれば、オダマキ属・トリカブト属・サラシナショウマ属などキンポウゲ科に多く見られる1室の心皮からなり、心皮の向軸面よりも縁辺にそって胚珠が2列につく縁生胎座と、スイレン科・アケビ科・ハナイ科・トチカガミ科のように子房は1室で、その向軸面全体に胚珠がつく面生胎座とがある。
また、子房とのかかわりでみれば、イグサ科・ラン科・ヤナギ科・スミレ科・ミツガシワ科・リンドウ科などに見られる一室子房の子房壁の縁辺近くに胚珠がつく側膜胎座、キンポウゲ科やバラ科の一部に見られる単一子房のもの 、ミクリ科・カヤツリグサ科・コショウ科・タデ科・イラクサ科・ムラサキ科・シソ科・キク科・カヤツリグサ科など複合子房のものがあり、いずれも少数個の胚珠が子房の基底部につく基底胎座、ナデシコ科やサクラソウ科にみられる、子房室は1室で基部が柱状に伸び出た部分に胚珠つく独立中央胎座、ユリ科・アヤメ科・ウマノスズクサ科・オトギリソウ科・ツツジ科・アカバナ科・カタバミ科・フウロソウ科・オオバコ科・キキョウ科などに見られる、隔壁で仕切られた2つの子房室をもち、その隔壁が中軸になって、それに沿うように胚珠がつく中軸胎座などがある。
助細胞からは、花粉管の誘引物質が絶えず作られて分泌される。それにより、花粉管が助細胞へと確実に伸長できる。被子植物の花粉管の伸長速度は、1時間あたり約10µm~20,000µm(2cm)以上とかなり幅がある。これは、裸子植物の花粉管の伸長速度の約100倍である。
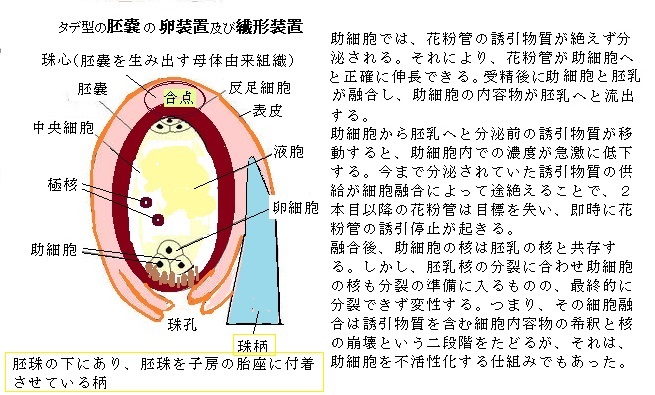 受精後に助細胞と胚乳が融合し、助細胞の内容物が胚乳へと流出する。助細胞から胚乳へと分泌前の誘引物質が移動すると、助細胞内での濃度が急激に低下する。
受精後に助細胞と胚乳が融合し、助細胞の内容物が胚乳へと流出する。助細胞から胚乳へと分泌前の誘引物質が移動すると、助細胞内での濃度が急激に低下する。このため安定して分泌されていた誘引物質の供給が細胞融合によって途絶えることで、2本目以降の花粉管は目標を失い、花粉管の伸長ができなくなる。
融合後、助細胞の核は胚乳の核と共存する。しかし、胚乳核の分裂に合わせ助細胞の核も分裂の準備に入るものの、最終的に分裂できず変性する。つまり、その細胞融合は誘引物質を含む細胞内容物の希釈と核の崩壊という二段階をたどるが、それが助細胞を不活性化する仕組みである。
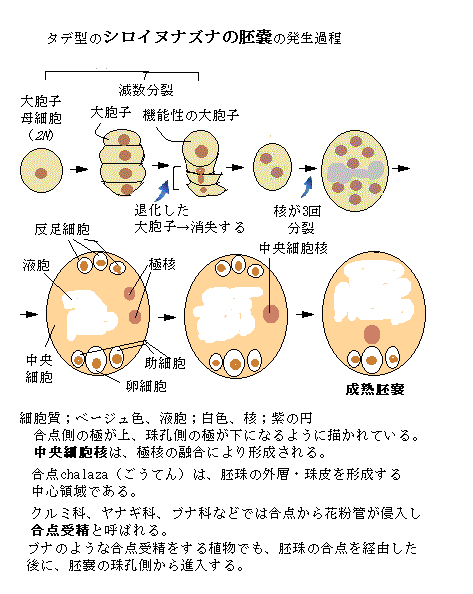
花粉管は、胚珠の入り口である珠孔を通って胚嚢に進入すると、伸長を停止し、先端が破裂することで、助細胞の1つに対して2つの精細胞が放出される。花粉管から精細胞が放出されると重複授精が起こる。2つの精細胞の内の1つが卵細胞と結合して受精卵を形成し、もう1つは、中央細胞と融合して三倍体の一次胚乳細胞を形成する。
吸水した雌性配偶体と精細胞が受精すると、胚発生が始まり、胚珠は種子に成長する。同時に、子房は肥大化して果実を育てる。
目次へ
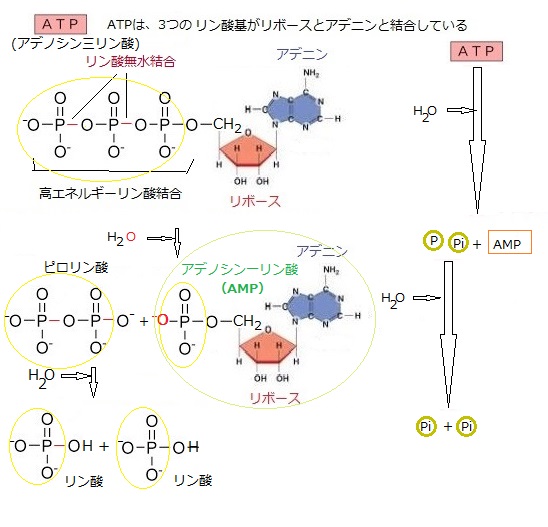
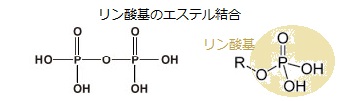 キナーゼkinaseは、リン酸化を触媒する酵素のことで、タンパク質などの基質にATPに由来するリン酸を付加することにより、その性質を変化させる。
キナーゼkinaseは、リン酸化を触媒する酵素のことで、タンパク質などの基質にATPに由来するリン酸を付加することにより、その性質を変化させる。受容体の機能を持つ、あるタンパク質が、シグナルを受容すると、その「シグナルを受容したこと」を、他の分子をリン酸化することによって、他の分子に伝達する。その場合のタンパク質を受容体キナーゼreceptor kinaseと呼ぶ。 受容体キナーゼのタイプにより標的タンパク質は、タンパク質分子上で、そのタンパク質を構成している様々なアミノ酸残基(セリン・スレオニン・チロシン・ヒスチジン)がリン酸化されることになり、生物活動が変化する。そのため、受容体キナーゼは、多様な動物のシグナル伝達機構において重要な役割を果たしている。
 植物でも、限定的であるが、重要な機能を担っている。特に、ホルモンであるブラシノステロイドbrassinosteroidの受容体であるBRR1受容体は、植物の発生に中心的役割を果たす。
植物でも、限定的であるが、重要な機能を担っている。特に、ホルモンであるブラシノステロイドbrassinosteroidの受容体であるBRR1受容体は、植物の発生に中心的役割を果たす。ブラシノステロイドは、オーキシンやジベレリンのように普遍的なホルモンで、陸上植物の出現前に、既に存在していたようだ。ブラシノステロイドは、細胞分裂・細胞伸長・細胞分化・光形態形成・生殖成長・発芽・葉の老化・ストレス応答など、植物の様々な器官に存在し、多様な発生現象に中心的な役割を果たす。例えば、花・葉・根には低い濃度で含まれ、花粉・未熟種子・果実では、比較的高い濃度で存在する。
シロイヌナズナのブラシノステロイド欠損変異体は、暗所で脱黄化de-etiolatedの表現型も示し、トウモロコシでは、雄花が雌性化する。
暗所で育った芽生えのシュートは、ひょろ長く、光合成活性を有しないため、シュート頂はフック(鉤状)のまま子葉は開かず、薄黄色の未展開の葉になっている、黄化etiolated(ˈiːtiəleɪtɪd軟白化)状態にある。逆に明所での芽生えは、短くて太い軸と光合成活性を示す葉緑体を持った子葉を展開する。暗所での芽生えを暗所形態形成skotomorphogenisis、明所での芽生えは光形態形成photomorphogenesisと呼ぶ。
暗所での、単子葉植物のトウモロコシの黄化の特徴は、当然、緑化は無理で、子葉の展開もできず、幼葉鞘(単子葉植物の発芽時に、芽生えを保護する円筒状の鞘)と中胚軸の伸長が著しい。
暗所と明所の2つ形成形態の切り替えは、複数の光受容体が光を浴びることにより、ゲノム全体での転写と翻訳の変化が関わる。 その過程は、一般的に、胚軸伸長速度の減少、シュート頂フックの立ち上がりの開始、光合成色素の合成開始と、複雑さを極めているが、その暗所形態形成から光形態形成への移行はすこぶる速い。この時、光は芽生えの形態変化を誘導するシグナルとなって、土の中では 、成長を促進し、明所では光エネルギーを効率的に収集し、必須となる糖・タンパク質や成長に必要な資質を生産できるように働いている。
暗所で育ったマメ科植物の芽生えは、1回のパルス光(約ピコ秒、即ち10-12秒単位の時間幅を持つ放射光)を照射した数分後には、複数の発生上の変化が起きると言う。
植物界には他の生物界と比較して、多くの受容体様セリン/スレオニンキナーゼreceptor-like serine/threonine⁽θríːənìːn ⁾ kinase(RLK)が存在する。RLKは、植物と病原菌との相互作用において重要な役割を果たすが、植物の受容体キナーゼの中でも、最大となるグループである。多くにRLKは、細胞外リガンド結合領域と細胞質キナーゼ領域の両方を持つ、膜貫通タンパク質として細胞膜に局在している。
細胞質キナーゼ領域は、標的タンパク質のセリンまたはスレオニン残基のリン酸化を介して、情報を細胞内に伝達する。チロシン残基をリン酸化するRLKもある。いくつかのRLKでは、リガンドが同定されている。そこには、生物間相互作用によって生産される、化学シグナルやブラシノステロイド・オーキシン・ペプチドホルモンなど、植物体が内生する植物ホルモンが含まれる。
RLKが介するブラシノステロイドシグナル伝達経路は、シグナルの増幅と抑制因子の不活性化による組み合わせにより、細胞外のブラシノステロイドホルモンの情報を転写反応に変換する過程である。それは、細胞膜上のブラシノステロイド受容体キナーゼのBRI1にブラシノステロイドが結合すると、リン酸化カスケードが活性化され、抑制タンパク質であるBIN2を不活性にする。これにより転写因子(BES1やBZR1)やそれに続く遺伝子発現が活性化される。
シグナルの受容と増幅における性質や構造は、極めて多様で複合的でありながら、実に精緻な仕組みが機能しており、しかも、被子植物における柱頭から子房内の珠孔まで花粉管が成長するまでの伝達経路transmitting tractには、一定の特徴が共有されている。
環境からの刺激や発生シグナルは、それぞれ特異な細胞表層の受容体に作用し、次に細胞内に拡散している標的酵素や細胞内受容体などのセカンドメッセンジャーsecond messengerに濃度変化を惹き起こす。それに関わる細胞内の機能変化を誘発し、その一連の動きを活性化させる。至適な反応が完了すると、フィードバック機構がシグナルを減衰させ、センサー機能がリセットされる。
細胞内受容体は、細胞全域に分布し生物界で保存されている。その受容体は、細胞膜・サイトゾル・内膜系や核などに含まれている。 植物の細胞は、様々な細胞の極性を制御するために、GTPase活性化タンパク質(GTPase-activating protein【GAP】)は、活性型の低分子量GTPaseと結合しているグアノシン5'-三リン酸GTPの加水分解を促進することにより、GTPase(guanosinetriphosphataseグアノシントリフォスファターゼ)を不活性化し、グアノシン5'-二リン酸GDPにするタンパク質である。GTPaseは、シグナル伝達経路において分子調節因子として働く。これらの調節性GTPaseは、活性型であるGTP結合型と不活性型であるGDP結合型を切り替える分子型スイッチである。
調節性GTPaseが活性型の場合、下流の情報伝達経路が誘導される。活性型から不活性型の切り替えは、GTPase自身によって行われ、結合しているGTPがGDPに加水分解される。GTPaseの型の転換過程は、別のタンパク質で調節される。これらのタンパク質は、GTPの加水分解、あるいはGDPの解離のいずれかの速度を変化させる。そのタンパク質の1つであるグアニンヌクレオチド交換因子guanine nucleotide exchange factor (GEF)は、GDPをGTPに交換することで、不活性型のGTPaseを活性型に変える。
一方、また別のタンパク質であるGTP加水分解酵素活性化タンパク質GTPase-activating protein(GAP)は、GTPの加水分解を促進することで、GTPaseを不活性化する。
植物の先端成長と極性を持つ細胞伸長(極性伸長)は、ROP(Rho-like of plants;GTPaseのグループの1つ)と呼ばれる特殊な低分子量GTPaseのファミリーによって調節される。ROPは、細胞骨格や小胞輸送の制御に関与する。シロイヌナズナには、11個のROP遺伝子があり、その内の7遺伝子は、成熟花粉粒と花粉管での発現量が多いか、あるいは選択的な発現をしている。
GTPaseであるROP1は、伸長中の花粉管先端の細胞膜のキャップ領域に局在し、先端成長を制御する。他の調節性GTPaseと同様に、ROP1の活性は、GEFによってオンに、GAPによってオフに切り替わる。そのGEFとGAPは、低分子量GTPase(ROP)の活性を制御し、花粉管の切り替え分子として機能する。
GEF自身も、シロイヌナズナのゲノム内で、大きな遺伝子ファミリーに属する受容体様キナーゼreceptor-like kinase(RLK)による情報伝達機構によって活性化される。花粉で発現するRLKが、GEFと直接的に相互作用をして先端成長を制御するというメカニズムが提唱されている。
未知のリガンドによって活性化されたRLKがGEFを活性化し、活性化されたGEFが、今度はROP1を活性化する。局所的に活性化されたROP1は、次にNADPHオキシターゼの活性を上昇させ、活性酵素種(ROS)が産出される。活性酵素種は、細胞外空間からのカルシウムイオンの流入を促進することで先端成長が促進される。
ROP1は、ROP-interactive CRIB motif-containing protein(RIC)と呼ばれるタンパク質群とも特異的に相互作用をする。RIC3とRIC4をシロイヌナズナで過剰発現させると、花粉管の極性とエキソサイトーシスが変化することから、RIC3とRIC4は、ROP1の下流で働くことが示唆された。さらなる解析により、RIC4径路は、アクチン繊維の重合を促進し、エキソサイトーシス小胞を先端部に蓄積させることが明らかになったものの、RIC3とRIC4が、花粉管の極性を変化させる詳細な仕組みは、未だ解明されていない。
目次へ