| Top 車山ブログ DNA 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 早春のスミレ 車山高原の笹 諏訪の植生 諏訪に生息する哺乳類 車山の山菜 車山高原の野鳥 諏訪の植生 諏訪に生息する哺乳類 ニホンジカの生態 |
 |
DNA DNAが遺伝物質 生物進化と光合成 葉緑素とATP 植物の葉の機能 植物の色素 葉緑体と光合成 花粉の形成と受精
ブドウ糖とデンプン 植物の運動力 光合成と光阻害 チラコイド反応 植物のエネルギー生産 ストロマ反応
植物の窒素化合物 屈性と傾性(偏差成長) タンパク質 遺伝子が作るタンパク質 遺伝子の発現(1) 遺伝子の発現(2)
遺伝子発現の仕組み リボソーム コルチゾール 生物個体の発生 染色体と遺伝 対立遺伝子と点変異 疾患とSNP 癌変異の集積
癌細胞の転移 大腸癌 細胞の生命化学 酸と塩基 細胞内の炭素化合物 細胞の中の単量体 糖(sugar) 糖の機能 脂肪酸
生物エネルギー 細胞内の巨大分子 化学結合エネルギー 植物の生活環 細胞のシグナル伝達 キク科植物 陸上植物の誕生
植物進化史 植物の水収支 拡散と浸透 細胞壁と膜の特性 種子植物 馴化と適応 水の吸収能力 稲・生命体 胞子体の発生
花粉の形成と構造 雌ずい群 花粉管の先端成長 自殖と他殖 フキノトウ
| 目次 |
| 1)花の形成 |
| 2)受粉と受精 |
| 3)花の起源 |
| 4)シアノバクテリア |
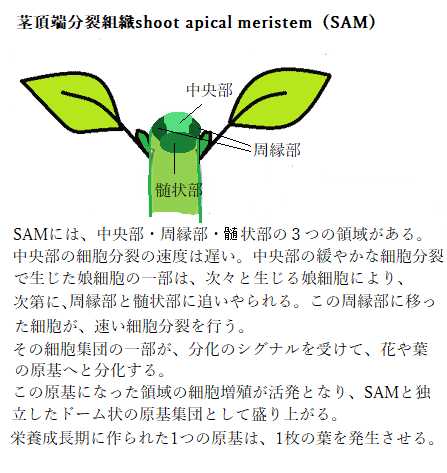 植物は発芽した後、通常は葉を作り続ける。春から秋、特定の季節が到来すると葉の発生を停止して花芽の形成(花成)を開始する。植物は花を決まった時期に開花させるために、花芽を作る花成期を見極めている。光と温度を頼りに季節の変化を感知しているようだ。それは植物の種子が発芽反応する条件と同様だ。光は「日の長さ(日長;にっちょう)」であり、温度は「一定期間の低温」とその後の「適度な成長温度」である。
植物は発芽した後、通常は葉を作り続ける。春から秋、特定の季節が到来すると葉の発生を停止して花芽の形成(花成)を開始する。植物は花を決まった時期に開花させるために、花芽を作る花成期を見極めている。光と温度を頼りに季節の変化を感知しているようだ。それは植物の種子が発芽反応する条件と同様だ。光は「日の長さ(日長;にっちょう)」であり、温度は「一定期間の低温」とその後の「適度な成長温度」である。こうした環境変化を、植物は葉で感じ取り、日長と温度が一定の条件を超えれば、茎の先端の「茎頂分裂組織」に、葉や茎に代わる花芽を形成させる。
植物が、花を咲かせる条件を葉で感じ取っていることは、既に20世紀の初めに突き止められていた。すべての葉を失うと植物は花を咲かせなくなる。たった一葉でも葉が残れば、それに適応する日長や温度を感知し花を咲かせる。そうした環境変化を感知する葉が、花を咲かせる上で重要な役割を果たしていた。
環境の変化を感じ取る葉が花を咲かせるのは、茎の先端にあり、そのため葉から茎の先端へ何らかの物質を信号として送っているようだ。
昭和12(1937)年、ソ連のチャイラヒヤン(M.H.Chailakhyan)は、葉から茎の先端へ送られる物質が花成ホルモンであると考えフロリゲン(florigen)と名付けた。それは「花を咲かせるもの」という意味だ。植物が光周期(1日の明暗サイクル)に反応して花をつける仕組は、葉がその光周刺激を受け取ると、葉にある種の物質が合成され、これが頂芽へ運ばれ頂芽の未分化な細胞を花の原基(個体の発生段階で、その形態や機能が器官としてまだ分化していない状態の細胞群)として変換させることによって花芽が形成されると考えられている。
葉と芽の間の茎を熱で破壊し、しかも環状剝皮すると光周誘導を行っても花芽は形成されないことから、花成ホルモン(フロリゲン)は師部(しぶ;養分の通道を主要な役目とする組織で、蔗糖を含む有機性栄養素を、植物全体の需要のある部分に輸送する生体組織)を通って移動するものと考えられている。
「冬生一年草」のシロイヌナズナは、秋に発芽し春に花を咲かせる。冬が過ぎ早春になり昼が長くなると葉が花成に必要な日長を感じる。すると葉の細胞に「CD」と名付けられたタンパク質が蓄えられる。「CDタンパク質」は「FT」と呼ばれるタンパク質を作る遺伝子(FT遺伝子)のスイッチをオンにして「FTタンパク質」を作る。つまり「CDタンパク質」は「FTタンパク質」の遺伝子の転写を制御する因子なのだ。
葉で作られた「FTタンパク質」は、維管束の篩管(細胞の内側)を通って茎頂分裂組織にまで送られる。そこで「FTタンパク質」は「FD」と名付けられたタンパク質と結合し「AP1」という花を作る遺伝子のスイッチをオンにして、葉になるはずだった芽を花へと変化させる。ここでは「FTタンパク質」は「AP1遺伝子」の転写因子になっている。
「フロリゲン」の正体は、かつては植物ホルモンの一つみられていたが、葉から茎頂分裂組織へ送られる「FTタンパク質」だった。タンパク質という物質自体が、生体内の反応に大きく影響を与える。イネでも同様のタンパク質が突き止められ、他の植物でも葉で作られたタンパク質により花成が行なわれているようだ。それで「花成ホルモン」とは言わなくなった。
花は茎頂分裂組織に「フロリゲン」が送られると、それまで作られてきた茎や葉の代わりに作られる。花は大きく「萼片」「花弁」「雄しべ」「雌しべ」の4つの器官で構成される。この4つの領域には、大きく3つのA.B.C.という転写調節因子が働き適切な時期に適切な器官を作り出している。これを「ABCモデル」という。
1. Aの転写調節因子が働いて「萼片」を作る。
2. AとBの転写調節因子が働いて「花弁」が作られる。
3. B.とCの転写調節因子が働いて「雄しべ」を作る。
4. Cの転写調節因子が働いて「雌しべ」を作る。
A転写調節因子の正体は、花成のプロセスで登場した「AP1遺伝子」が作る「AP1タンパク質」である。葉を作っていた茎頂分裂組織に「FT」と「FD」の2つのタンパク質が作用すると「AP1遺伝子」が発現し、「AP1タンパク質」が作られる。それが花器官を作る最初のスイッチとなり、領域1で「萼片」が作られていく。それに合わせてBやCの転写調節因子が働き始めて、残りの「花弁」と「雄しべ」「雌しべ」がそれぞれの領域で形成される。
「ABCモデル」として作用する3つの転写調節因子を作る遺伝子に変異が生じると、「萼片」「花弁」「雄しべ」「雌しべ」の4つの花器官は正常に作られず、他の器官に変化する。B転写調節因子を作る遺伝子に変異が生じれば、領域2ではA転写調節因子だけが機能して「萼片」が作られ、領域3ではC転写調節因子がけが働いて「雌しべ」が作られる。さらにABCの転写調節因子を作る遺伝子すべてに変異が生じれば、「萼片」「花弁」「雄しべ」「雌しべ」などの花器官は作られず、すべて葉のようなものに置き換わる。このような変異が生じた個体を「三重突然変異体」という。
花成とは、花芽が形成され始めることをいい、植物に花成を促すのが花成ホルモンと言われてきた。花成ホルモンの実態が明らかになってきたのは、ここ数年のことである。モデル植物、イネおよびシロイヌナズナを用いた分子遺伝学的な研究の成果により、現在、花成ホルモンは、花成に関与する特定の遺伝子から作られる「球状たんぱく質」であることが明らかになり、分子レベルでの理解も深まってきている。
この花成タンパク質は葉で作りだされ、花芽が形成される茎頂まで運ばれ、そこで花芽形成に関与する。その花成タンパク質の構造や作用の解明が進めば、その応用研究により、作物や樹木などの開花を人為的に調節できるようになる。
この植物の花成を人為的に制御できれば、農業において非常に有用である。特に樹木は苗から開花するまでに数年から数十年を要し、交配による品種改良は容易でなかった。フロリゲンにより幼樹段階で開花させられれば、育種期間は大幅に短くなり、品種改良の速度は格段に促進される。
目次へ
シアノバクテリアを取り込むことに始まる植物の20数億年の進化の過程から見れば、花を咲かせるようになったのは、それほど古くはない。
2015年4月14日、新華報業網によると、「中国遼寧省で見つかった花の化石が世界最古の花と認定された。この花は1億6200万年前のジュラ紀に生息していた花で、これまでは1億2500万年前の化石が世界で最も古い花とされていた。今回新たに認定された花は典型的な花の形をしており、花弁や雄しべ、雌しべなどがある。今回の発見が被子植物の起源の研究に新たな1ページを切り開くと期待されている」とある。
それは花の化石レベルの話で、考古学上では「それ以前に植物が花を咲かせる証拠は見つかっていない」と言うだけで、生物学上、それだけで、その年代が花の起源とまでは言えない。それでも生物40億年の歴史から見れば日が浅い。
花とは、種子植物(顕花植物)の持つ生殖器官を指す言葉で、有性生殖をする種子植物が、花と呼ばれる生殖器官の特殊化が進んで、胚珠(種子植物の種子になる部分)が雌しべの基部にあたる子房の中に収まったものをいう。被子植物の生殖器官でも、本来必ずしも美しい形や色をしているものではなかった。
車山高原で一段と高く聳えるトウヒのような裸子植物の生殖器官は目立たない。6月を新緑の時季とする車山の寒冷地であればこそ、山桜・コブシ・フジが車山山麓の山肌を彩る。 花はどのようにして今のような姿になったのか。花の進化は、詩人としても形態学者としても実績のあるドイツの文豪ゲーテをはじめ、多くの研究者によって論じられてきた。 何が花の起源なのだろう。
 ゲーテは1,970年に『植物変態論』という本を出版した。文豪ゲーテは、劇作家でもあり、植物学はもとより、地質学・気象学などの自然科学の分野でも多くの業績を残していた。
ゲーテは1,970年に『植物変態論』という本を出版した。文豪ゲーテは、劇作家でもあり、植物学はもとより、地質学・気象学などの自然科学の分野でも多くの業績を残していた。『植物変態論』で、「花は葉のメタルフォーゼ(変形)である」と述べている。ゲーテは、従来の植物研究に拘らず、その詳細でしかも正確な観察により、現代の自然科学の分野に多くの業績を残した。
そのゲーテの仮説は、それから20年後の1,991年、遺伝子レベルで実証された。花を作る遺伝子に異常がある個体は、花が咲くべき位置に葉が生える。
花を咲かせ、実を結び、種子によって繁殖する顕花植物には、裸子植物と被子植物を含む。その花を構成する萼・花弁・雄しべ・雌しべの諸器官の起源を探ると、すべて葉の変形したものと言える。 花の誕生は植物のみならず、その後の生物の進化には画期的な影響を与えた。人間が食べるものの75%以上は、「顕花植物」に由来する。
特に被子植物の赤や黄色、紫やピンクなど、多くの花の色彩は、虫や鳥など花粉を運んでくれる動物たちを引き付けるためだ。鳥媒花の色は、通常、赤か白である。虫媒花より濃い密を出すのが特徴だ。昆虫が未だ活動しない早春に赤い花を咲かせるヤブツバキ・サザンカは、メジロやヒヨドリにより送粉が行なわれる。白い花のコブシは蜜を出さないが、花弁に栄養があり、それを食べるヒヨドリが花粉を運ぶ。6月-7月に赤い花を咲かせるザクロには、ヒヨドリが訪花する。
針葉樹のマツ・ヒノキ・スギ・イチイなどの裸子植物は、花を咲かせても、形や色など、その花の美しさが問われることはない。花粉は風により運ばれる風媒花、風まかせであるため多量の花粉を放出する。スギは雌雄同株であるが、単性花といわれ、雄花と雌花を別々に咲かせる。これを雌雄異花(しゆういか)と言う。キュウリ・カボチャ・スイカ・カラスウリなどもそうだ。
スギ花粉は、2月から4月まで飛散するため、スギ花粉症の患者はこの時期に急増するため、春になると花粉症の元凶として嫌われる。雄花の長さは5mmほどの楕円体で、枝先に下向きにぎっしりと群がっている。枝と共に風にあおられて揺れる。1個の雄花の中には、24~40万個の花粉が入っている。その雄花の上の方で下向きに咲いているのが雌花だ。そのため、花粉を受けにくい。200万個当り1個の確率で受粉できるという。
そのため、よく晴れた風の強い日に、雄花から小さくて軽い花粉を大量に放出し、あたり一面に散布する。 その前に、雄しべの先端にある葯の中で細胞が分裂を繰り返し、多数の「花粉母細胞」が作られる。その「花粉母細胞」は、2度行なう細胞分裂によって「花粉四分子」と呼ばれる4つの未熟な花粉になる。
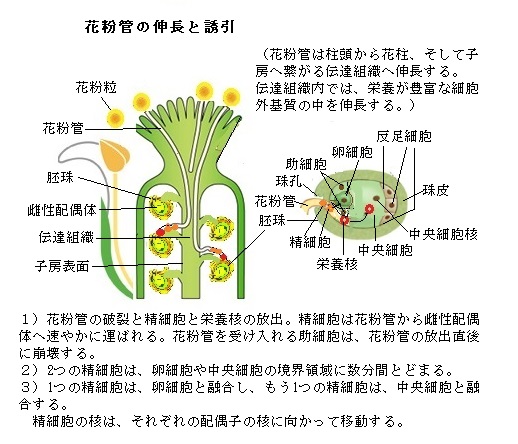 「花粉四分子」は更に細胞分裂を重ね、栄養細胞(あるいは花粉管細胞)と雄原細胞(ゆうげんさいぼう)に分かれる。雄原細胞は花粉管細胞(栄養細胞)に取り込まれるので、二重膜の状態で存在する。この状態のまま、雄原細胞は再度分裂し、2つの精細胞になる。つまり、2つの精細胞も、花粉管細胞の細胞膜に由来する膜で包まれている。一つの花粉の中は、一つの栄養細胞と、それに取り込められた二つの精細胞で構成されている。その花粉が雄しべの葯から飛び出して、雄しべの柱頭にたどりつく。そこで雌しべのチェックを受ける。別の個体と認められると受粉が成立する。
「花粉四分子」は更に細胞分裂を重ね、栄養細胞(あるいは花粉管細胞)と雄原細胞(ゆうげんさいぼう)に分かれる。雄原細胞は花粉管細胞(栄養細胞)に取り込まれるので、二重膜の状態で存在する。この状態のまま、雄原細胞は再度分裂し、2つの精細胞になる。つまり、2つの精細胞も、花粉管細胞の細胞膜に由来する膜で包まれている。一つの花粉の中は、一つの栄養細胞と、それに取り込められた二つの精細胞で構成されている。その花粉が雄しべの葯から飛び出して、雄しべの柱頭にたどりつく。そこで雌しべのチェックを受ける。別の個体と認められると受粉が成立する。 雌しべは、大まかに言えば、「柱頭(ちゅうとう)」「花柱(かちゅう)」「子房」から構成され、子房の中の基部となる胚珠で卵細胞(らんさいぼう)が作られる。「子房」は後に果実となり、その中でやがて種子となる胚珠で卵細胞が形成される。 受粉に成功した花粉は、雌しべの胚珠に向って「栄養細胞(花粉管細胞)」の一部を管状に伸ばしていく。この「花粉管」とよばれる管の中を2つの精細胞が前方に進んでいく。自分自身で移動するのではなく、この大きな花粉管細胞の膜中に包みこまれたまま前方に輸送されて行く。植物の精細胞は動物の精子と異なり、運動能力を持たない。そのため花粉管を胚珠に向けて伸長させ、この花粉管細胞の中で精細胞を運ぶ仕組みを作った。
近年、名古屋大学の東山哲也教授らは、卵細胞の部分が母体組織から突き出る「トレニア」というユニークな植物を使って、卵細胞の隣にある「助細胞」が誘引物質を分泌することを世界に先駆けて証明した。 この物質の正体は、低分子のアミニ酸・ペプチドであることが分かった。「LURE(ルアー;誘引する意)」と名付けられた。「LURE」は植物の種類ごとに違ったアミノ酸で構成されていた。これにより花粉が異なる植物種の雌しべに受粉しても「受精」しない理由の一つとして知られた。
「トレニア」は熱帯原産の植物で、実験材料として研究室などでもよく用いられる。通常、被子植物の胚嚢は子房の中に入り込んでいるため、生きた状態で観察はできない。「トレニア」は例外的に胚嚢の一部が剥き出しになっている。そのため、特に受精のメカニズムを解明する研究などでは、最も扱いやすい植物とされている。 動物では一つの精子と一つの卵細胞が受精するのが通例である。被子植物では、二つの精細胞によって受精のような現象が2度起こる。そのため被子植物の受精を「重複受精」と呼ぶ。この重複受精という現象は被子植物だけが特有するものではない。裸子植物で、主として東南アジアに分布するグネツム科グネツム属の常緑低高木グネモンの果実は、中に1個の種子があり、その種皮の中に胚乳も作られ、インドネシアなどでは食用とされている。
被子植物では、二つの精細胞が花粉管が胚嚢まで達すると、それぞれが「卵細胞」と「中央細胞」という生殖細胞に届くと受精は完了する。受精が完了すると、花粉がどれだけ雌しべについていようと、それらが花粉管を伸ばしている途中であろうと、助細胞による誘引は終了する。
精細胞と「卵細胞」が受精して「受精卵」となり、それがやがて「胚」となって植物の幼体となる。 精細胞と「中央細胞」が受精した方は、種子の中で「胚」の栄養となる「胚乳」になる。「胚」と「胚乳」は、それらを包む種皮とあわせて種子を形成する。胚珠が種子になると胚嚢を包んでいた「珠皮」が種皮となり、子房が発達して果実になると、それを包む「子房壁」が果皮に変わる。
植物はヒトと違って、一つの個体にいくつもの花を咲かせ、同時に受精し多くの種子を誕生させる。有性生殖を行なう生物は、進化の過程で、自分の遺伝情報を受け継いだ子孫が、有利に生存を全うできるようと生殖の仕組みを獲得していく。被子植物の重複受精も、花粉が他の花粉たちによる競合状態を制するため、それぞれが受精を成し遂げた花粉による種子が有利になるように、重複受精によって大きな胚乳を作り上げた。その遺伝情報を継ぐ種子の生存力を一段と高めるため、その栄養源を発達させていった。
一方、植物全体では、その体内で作られる多くの種子が、できるだけ数多く散布されるよう、一つひとつ種皮の中の胚乳に偏りが生じないよう調整をする。またその後の自らの生存も、もとより重要だ。しかし、植物は全般的にリスキーだ。無責任に多量に散布するが、日が当たる土地でなければ、光合成は不可能で生存は無理だ。条件が整った土地に種を落すことの確立の低さを想定し、多量の種子を犠牲にしながらも、稀に環境に適合する大地で芽吹くことを期待している。
植物によっていろいろな型があるが、被子植物の胚珠の中にできる雌性配偶体、いわゆる胚嚢内で花粉管先端が破裂し、精細胞が放出された後は、この精細胞の外側の膜は観察されない。精細胞が花粉管から放出される際に、物理的に失われるのではないかと見られている。外側の膜はなくなるので、2つの精細胞はそれぞれ別個に、卵細胞あるいは中央細胞の膜と融合でき、重複受精がなされる。この際、精細胞と、卵細胞や中央細胞の受精が起こる領域には、細胞壁はない。 雄原細胞が花粉管細胞に取り込まれた直後には、細胞壁成分が観察される。受精前までに分解されるということになるが、その詳しい制御の仕組みは分からないままだ。
スギの雄花の中心の軸のまわりには、花粉の入っている袋、葯がいくつも付着している。雄花の表面は、魚の鱗のような鱗片で覆われている。雄花が成長し鱗片が開くと、中にある葯が乾燥して破れ、花粉が放出される。こうして無数に出た花粉は、風に乗って、極めて幸運な奇跡的とも言える好条件の下、ごく稀に雌花にたどり着く、すると雌花の中にある管のようなものの先に、透明な液体がにじみ出る。雌花はここで、飛んできた花粉を捕らえる。管の先についた花粉は、ゆっくりと雌花の内部へ入り込む。この時、花粉は水を吸収して破裂する。破裂して出てきた丸いものが、精細胞(せいさいぼう)である。受粉しても種子が必ず作られるとは言えない。受粉の後に、動物と同様、「受精」しなければ意味がない。葯から放出される花粉には、動物の精子に相当する「精細胞」という生殖細胞が含まれている。その花粉が雌しべの柱頭に送粉され付着する。すると花粉から花粉管が伸び、その中を「精細胞」が移動して子房の中核となる胚珠の基部に納まる「卵細胞」にたどり着き合体する。 この精細胞が雌花の卵細胞と出会い、ここに「受精」が成立し種子が作られる。やがて実を結ぶ。
雌雄異株(しゆういしゅ)では雄の植物と雌の植物に分かれている。動物が雄と雌に分かれているのと同じだ。草本類でもアサなどで確認されているが、圧倒的に樹木に多い属性である。イチイ・イチョウ・アオキ・モクセイ・クワ・ヤナギ・ツルウメモドキ・キウイなどがある。自家受粉を避け、他の株の花粉を受精した方が、より多様な遺伝子を潜在化させられ、環境変化が当たり前とされる自然選択に耐えられる戦略をシステム化させた。
神社や街路樹に植えられ、東京都のシンボルマークとなっているイチョウも雌雄異株の植物である。最近では街路樹のイチョウの実ギンナンが路上に落ちて臭くて汚いと言い、雄の株だけを選んで植えているようだ。
目次へ
モデル植物であるシロイヌナズナの遺伝子解析が行なわれた結果、花の形成を制御している遺伝子の多くには、決まった配列があることがわかった。MADS boxとK boxと呼ばれる配列である。
シダとしては珍しい水草のリチャードミズワラビにあるMADS遺伝子(MADS boxを持つ遺伝子をこう呼ぶ)はどんな働きをしているのだろう。この植物のMADS遺伝子は9個あり、塩基配列の類似性によって二つのグループに分けられた。各グループから一つずつ遺伝子を選び、作用している時期や場所を調べてみると、ともに胞子嚢が形成される初期から発現していた、という。 シロイヌナズナの花も、萼・花弁・雄しべ・雌しべという器官からできている。雌しべを作る指令を出すAGAMOUS(ǽgəməs)遺伝子は、雌しべができると新たな器官が出来ないように作用する。シロイヌナズナでは、このAGAMOUS遺伝子が突然変異を起こして機能しなくなると、雌しべの代わりに花弁や萼が次々と作られ、いわゆる八重咲きのような花になる。多くの花の八重咲きも同じような仕組みで起きていると考えられている。
シロイヌナズナの場合、この八重咲きの花を短日条件(昼の長さより夜の長さの方が長い)などで育てると、2重の花になることがある、という。 バラのAGAMOUS遺伝子の作用が弱まると、本来雌しべになるべき場所から、新たな花が作られることがある。これを「貫性のバラ」と呼ぶ。園芸用のバラでは、養分過多などいくつかの条件がそろうと、このような花が見られるそうだ。
バラの原種は5枚の花弁と多数の雄しべを持っている。園芸品種のバラの花弁の数が多いのは、雄しべが花弁に変化するAGAMOUS遺伝子の突然変異体を人為的に選抜し、育種してきたためと考えられている。
リチャードミズワラビMADS遺伝子は二つの群に分かれ、一方は種子植物のAG群(AGAMOUS遺伝子と似た配列を持つ遺伝子の群)と同じ系統になり、他方はどの群とも近縁ではなかった。つまり、リチャードミズワラビでは、種子植物のMADS遺伝子が「入れ子構造」になっていた。それを系統樹でとらえれば、植物MADS遺伝子の中の各遺伝子群は、シダ類と種子植物が分岐するよりも前に既に双方に存在していたことになる。即ち、シダ類と種子植物の間には、共通の祖先はなく樹枝状の線を異にしていた。
ところで、被子植物でたくさん見つかっているAG群の遺伝子のうちの一つは、本来作用しないはずの萼などで働かせると、胚珠が形成されるという研究がある。リチャードミズワラビのAG群に属する遺伝子も、その役割は、胞子嚢を葉の表皮から誘導することではないだろうか。
ゲーテが最初に気付いた「花は葉のメタルフォーゼ(変形)である」という萼・花びら・雄しべ・雌しべといった花の各器官は、葉から生殖細胞を包む袋、すなわち胞子嚢を誘導することに起源があるためとみられる。
MADS遺伝子は様々な作用をする遺伝子として機能分化しているが、そのほとんどは生殖器官の形成に関与している。MADS遺伝子の研究が深まれば、花の起源にまで迫る。そのため、花の起源を知ろうとして花の形成に関与する遺伝子の起源を探った結果、それは、胞子嚢の形成に関与する遺伝子であるというところまで分かってきた。これからは、MADS遺伝子の一群が、どこでどのように作用するか解明されれば、地味な胞子嚢作りから、やがて華やかな花の形成までにいたった過程が解明されていくだろう。
花作りに関係している遺伝子は、MADS遺伝子以外にも知られているので、今後も、それらの遺伝子群の解析が急がれると同時に、その関連性が明らかになれば、生物学自体の進化が一段と加速される。
今世紀初めに、植物同士を形で比べることにより、植物全体の大まかな系統樹ができ上がった。近年、DNAの塩基配列を比較し、系統関係が新たに調べ直され、様々な生物の解析に活用されるようになった。光合成など植物が生きるために必須な生命活動を司る遺伝子は、どの植物にも存在するので、これを利用すれば、形では比べようもない遠縁のもの同士でも比較できるようになった。そして、DNA解析を行なうと、これまで形から得られていた結論とは異なるデータ解析の研究成果が続出している。
たとえば、現存の裸子植物は、イチョウ・ソテツ・針葉樹・グネツム類に分類され、互いに形、特に生殖器官の形が大きく異なるので、違う系統だと考えられてきた。そして、グネツム類は雌の生殖器官が被子植物に似ているため、被子植物にもっとも近縁だとみられていた。ところがこれらを、裸子植物のDNAと比べてみると 、どの裸子植物も共通する祖先から派生していた。被子植物と裸子植物は祖先を共有せず、系統樹における樹枝状の線を異にしている。 つまり、そのいずれも被子植物の祖先とはなりえず、現存する裸子植物と被子植物が分かれたあとに、裸子植物の生殖器官が進化の過程で多様化しようだ。
目次へ
藍藻(らんそう、blue-green algae)は、藍色細菌(らんしょくさいきん、cyanobacteria)の旧名である。同じく葉緑素(クロロフィル)を持つが、藻類ではなく、シアノバクテリア(藍色細菌)と呼ばれる細菌(バクテリア)の1群である。
光合成によって酸素を生み出す酸素発生型光合成細菌である(バクテリアは細菌のラテン語での呼び名)。細菌とは、原核細胞つまり核膜は無いが、固い細胞壁がある単細胞の生物である。大腸菌などの細菌類や藍藻類などと同様、染色体がほぼ裸のまま細胞内にあり、核膜(真核生物の核を細胞質から隔てている生体膜)がない原核生物(バクテリア)だ。
ヒトなど脊椎動物・植物・原生動物などほとんどの生き物が、細胞内に核膜をもつ真核生物である。
シアノバクテリアが、バクテリアの仲間といっても、他のバクテリアと違って葉緑素を持ち光合成をすることができる。顕微鏡下で観察すれば、単細胞で浮遊するもの、少数細胞の集団を作るもの、細胞列が糸状に並んだものなどがある。
シアノバクテリアは、その名の通り、青っぽい緑色藍色細菌とも呼ばれる細菌の1群であり、光合成によって酸素を生み出す。それはまさに葉緑素をもち、太陽光エネルギーを化学エネルギーに変換する、地球上で最初に光合成を行なった光合成細菌と言える。
細胞は水以外の殆どの部分がタンパク質でできている。タンパク質は、生体の細胞内にあるリボソームで合成される。原核生物も細胞内のタンパク質は、その細胞のリボソームで合成される。
葉緑体のリボソーム(Ribosome)RNAの塩基配列は、シアノバクテリアの中にも含まれる。リボソームは、mRNA(messenger RNA)の遺伝情報を読み取ってタンパク質を合成するという場である。葉緑体を生み出す細胞内共生が1回だけ起きたという進化が、その後の地球の運命を変えた。次に、植物や二次共生藻類の起源が、シアノバクテリアを細胞のなかに取り込んで葉緑体として共生する進化の過程を経たことに始まる。
目次へ